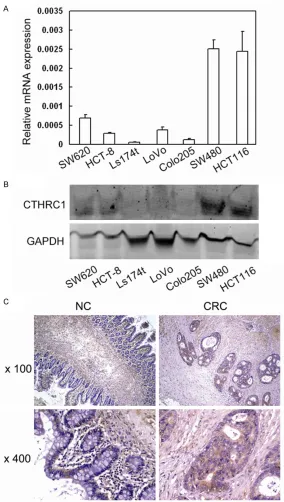
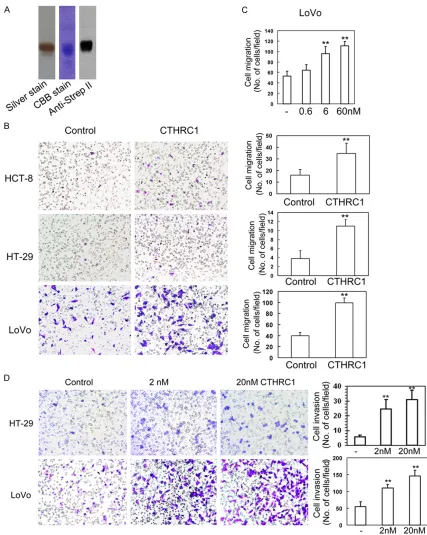
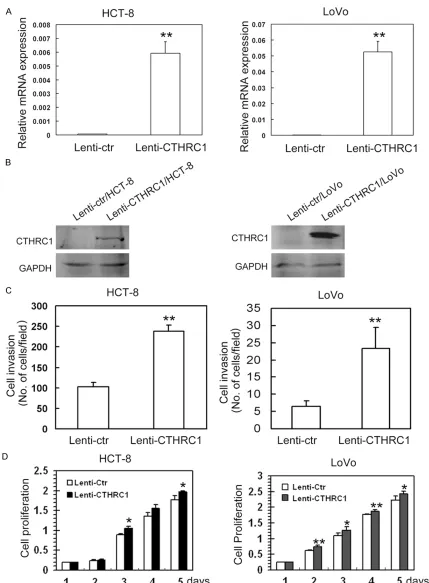
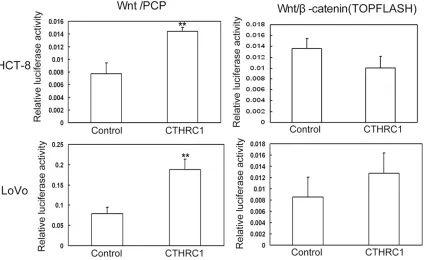
Original Article CTHRC1 promotes human colorectal cancer cell proliferation and invasiveness by activating Wnt/PCP signaling
Full text
Figure
Related documents
To understand the mechanisms of pharmacological actions, the in-vitro antioxidant activity of alcoholic and aqueous extracts of leaf, root and seeds of
In this study we demonstrated that BA had an in vitro and in vivo inhibitory effect on TNBC, via G2/M cell cycle arrest, induction of apoptosis, inhibition of cell migration,
2 illustrates process steps of reading the medical images, the segmentation processes in the medical images, the fractal geometry processes as the method required to be applied,
In Elliptic Curve the Secret Messages are hidden in the Curve points .The Elliptic Curve Randomly selects the Audio samples say 8 bits from the Secret File.. The Hidden message will
inhibition of BRAF with a specific BRAF-V600 inhibitor appears to cause paradoxical CRAF-mediated activation of the ERK signaling pathway in wild-type BRAF cells and
caused by mutations in human NADPH cytochrome P450 reductase.. Marohnic CC, Panda SP, McCammon K, Rueff J, Masters BSS, Kranendonk M 2010
The analysis unveils the existence of two propagating modes for the longitudinal and the transverse waves, leading near the position of the main peak to two peaks in the
Impact of Explicit Instruction Using Culturally Relevant Texts in Reading on Student Confidence Wendy Harriford Platt.. Follow this and additional works
