RESEARCH ARTICLE
Transcriptional Profiling of Developing Mouse
Epidermis Reveals Novel Patterns of
Coordinated Gene Expression
Hisham Bazzi,1Katherine A. Fantauzzo,1Gavin D. Richardson,3Colin A.B. Jahoda,3and Angela M. Christiano1,2*
The mammalian epidermis is the first line of defense against external environmental challenges including dehydration. The epidermis undergoes a highly intricate developmental program in utero, transforming from a simple to a complex stratified epithelium. During this process of stratification and differentiation, epidermal keratinocytes express a defined set of structural proteins, mainly keratins, whose expression is controlled by largely unknown mechanisms. In order to identify novel factors contributing to epidermal morphogenesis, we performed a global transcriptional analysis of the developing mouse epidermis after separating it from the underlying dermis (E12.5–E15.5). Unexpectedly, the recently identified genes encoding secreted peptides dermokine (Dmkn), keratinocyte differentiation-associated protein (krtdap), and suprabasin (Sbsn) as well as a largely uncharacterized embryonic keratin (Krt77), were among the most highly differentially expressed genes. The three genes encoding the secreted proteins form a cluster in an ⬃40-Kb locus on human chromosome 19 and the syntenic region on mouse chromosome 7 known as the stratified epithelium secreted peptides complex (SSC). Using whole mount in situ hybridization, we show that these genes show a coordinated spatio-temporal expression pattern during epidermal morphogenesis. The expression of these genes initiates in the nasal epithelium and correlates with the initiation of other epidermal differentiation markers such as K1 and loricrin (Byrne et al. [1994]Development120:2369 –2383), as well as the initiation of barrier formation. Our observations reveal a coordinated mode of expression of the SSC genes as well as the correlation of their initiation in the nasal epithelium with the initiation of barrier formation at this site.Developmental Dynamics 236:961–970, 2007.©2007 Wiley-Liss, Inc.
Key words:epidermis; morphogenesis; differentiation; coordinate expression; dermokine; keratinocyte differentiation-associated protein; suprabasin;Krt77
Accepted 23 January 2007
INTRODUCTION
Epidermal stratification and differen-tiation during mammalian develop-ment are critical for the survival of the
animal in the postnatal terrestrial en-vironment (Sengel, 1976; Byrne et al., 2003; Segre, 2003). In the mouse em-bryo, the commitment to the transfor-mation of the simple epithelium into a
stratified complex epidermis starts as early as embryonic day 7.5 (E7.5) and is completed by E17–E18.5, when the full epidermal barrier is established (Byrne et al., 2003; Koster and Roop,
The Supplementary Material referred to in this article can be viewed at www.interscience.wiley.com/jpages/1058-8388/suppmat. Grant sponsor: Kirsch Foundation.
1Departments of Genetics and Development, Columbia University, New York, New York 2Dermatology, Columbia University, New York, New York
3Department of Biological Sciences, University of Durham, Durham, United Kingdom
*Correspondence to: Angela M. Christiano, Ph.D., Department of Dermatology, Columbia University, College of Physicians & Surgeons, 630 West 168th Street, VC-1526, New York, NY 10032. E-mail: [email protected]
DOI 10.1002/dvdy.21099
Published online 1 March 2007 in Wiley InterScience (www.interscience.wiley.com).
ABBREVIATIONSDmkn/DmknDermokine gene/proteinKrtdap/Krtdapkeratinocyte differentiation-associated protein gene/protein
Sbsn/Sbsn Suprabasin gene/protein Krt77 keratin complex 2 basic, gene 39 Krt keratin gene K Keratin protein EDC epidermal differentiation complexSSCstratified epithelium secreted peptides complex
2004; Laurikkala et al., 2006). During this transformation, the forming epi-dermis expresses the cytokeratin pair 5 and 14 (K5/14) and down-regulates cytokeratins 8 and 18 (K8/18), mark-ers of single-layered simple epithelia (Koster and Roop, 2004; and this work). The expression of the tran-scription factor p63 (TAp63 in partic-ular) precedes that of K14 and regu-lates its expression. Indeed, p63 is both a master regulator and among the earliest signs of the commitment to epidermal stratification and differ-entiation (Koster et al., 2004; Koster and Roop, 2004; Laurikkala et al., 2006).
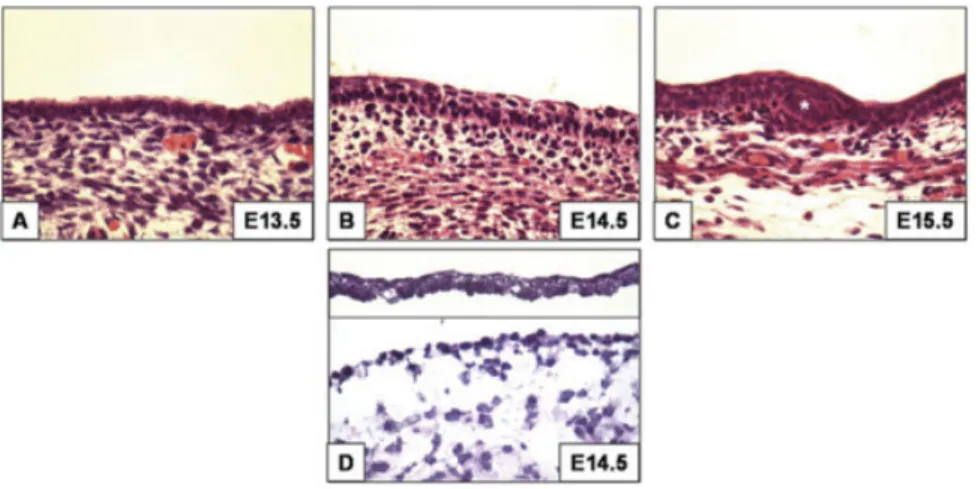
The first asymmetric vertical cell di-visions in the epidermis give rise to the periderm (⬃E9.5). This relatively simple structure is believed to tran-siently protect the embryo from the amniotic fluid until the functional bar-rier forms (Byrne et al., 2003). How-ever, it is not until around E14.5– E15.5 that the first morphological signs of a three-tiered stratification and differentiation program are evi-dent (Fig. 1) (Byrne et al., 1994). In this respect, the differentiated kera-tinocytes of the spinous layer of the epidermis appear above the prolifera-tive basal keratinocytes and show cy-tokeratin 1 (K1) immunoreactivity, an established marker for the spinous layer in adult life (Histology in Fig. 1B,C; E14.5–E15.5) (Fuchs and Green, 1980; Byrne et al., 1994; K1 immunofluorescence not shown). At E15.5 and later, the granular layer keratinocytes arise from the differen-tiating spinous layer and express the cornified envelope precursor protein loricrin (Histology in Fig. 1B,C; E14.5–E15.5) (Byrne et al, 1994; Lori-crin immunofluorescence not shown). The final stages of barrier formation begin with the terminal differentia-tion of the cornified keratinocytes to form the hydrophobic cornified layer (Hardman et al., 1998; Byrne et al., 2003).
In adult life, the same program of differentiation is reiterated with basal keratinocytes detaching from the basement membrane and ascending through the differentiation program until they are shed, a process that takes place every two weeks in mice (Sengel, 1976). The differentiating epidermal keratinocytes express a
large number of epidermal-specific differentiation genes, many of which are clustered on mouse chromosome 3 and the syntenic region on human chromosome 1q21 (Volz et al., 1993; Rothnagel et al., 1994; Mischke et al., 1996). This gene cluster is termed the epidermal differentiation complex (EDC) and harbors genes encoding three families of proteins (Volz et al., 1993; Mischke et al., 1996). The first family, characterized by short tandem peptide repeats in the central region, includes proteins such as loricrin, in-volucrin, and small proline-rich pro-teins (Sprr). The second family (the fused group) includes proteins such as profillagrin and trichohyalin with an EF-hand domain (Ca2⫹-binding do-main) in their N-terminal region fol-lowed by tandem repeats. The third family is represented by S100 family members also with EF-hand domains (Volz et al., 1993; Rothnagel et al., 1994; Mischke et al., 1996).
Recently, a new stratified epitheli-um-related gene cluster termed the stratified epithelium secreted pep-tides complex (SSC) was identified (Matsui et al., 2004; Moffatt et al., 2004). This gene cluster of⬃40 Kb on human chromosome 19 and mouse chromosome 7 was discovered using subtractive hybridization screens, sig-nal-trap assays, and high-throughput in situ hybridization (ISH), and har-bors at least three genes encoding se-creted peptides, dermokine (Dmkn), keratinocyte differentiation-associated protein (Krtdap), and suprabasin (Sbsn) (Oomizu et al., 2000; Park et al., 2002; Matsui et al., 2004; Moffatt et al., 2004; Tsuchida et al., 2004). Not only are these genes tightly clustered in the genome, but also they are tran-scribed in the same orientation and their mRNAs are expressed in the su-prabasal layers of stratified epithelia concomitant with the appearance of the differentiated layers (Fig. 2D,E) (Oomizu et al., 2000; Park et al., 2002; Matsui et al., 2004; Moffatt et al., 2004; Tsuchida et al., 2004).
In this study, we utilized microsur-gical techniques to separate develop-ing mouse skin at 12-hr intervals be-tween E12.5–E15.5 into its epidermal and dermal compartments (e.g., in Fig. 1, E14.5 histology of separated epidermis and dermis). We performed microarray analyses using MOE430A
chips from Affymetrix on the tran-scripts extracted from both compart-ments and compared their relative expression during morphogenesis. Surprisingly, we found that Dmkn andKrtdap had the two highest dif-ferentially expressed transcripts in the developing epidermis (more than 1,900-fold by E15.5), whereasSbsnis not represented with a probe on this array. Moreover,Krt77, a largely un-characterized mouse embryonic kera-tin gene (Hesse et al., 2001, 2004; Rog-ers et al., 2005; Senshu et al., 2005), was also among the highly induced mRNAs during epidermal differentia-tion (827-fold by E15.5). Using whole mount ISH, we observed a coordi-nated spatio-temporal expression of the SSC cluster genes that is corre-lated with the initiation of barrier for-mation in the nasal epithelium later in epidermal development.
RESULTS
Transcriptional Profiling of
Developing Mouse Epidermis
In order to identify novel factors in-volved in both epidermal and hair folli-cle morphogenesis, we used microdis-section techniques coupled with microarray analyses on the developing epidermis (Fig. 1) (E12.5–E15.5, see Ex-perimental Procedures section). We performed the microarray experiment with three or four biological replicates of epidermis at each time point. We used two commercially available soft-ware packages, GeneTraffic and Gene-Spring, to normalize and analyze the differentially expressed genes between the time points. We also utilized these programs to perform unsupervised hierarchical clustering analyses of the biological replicates to assess correla-tion within and between the replicates. Both softwares gave comparable results for clustering and differential expres-sion analyses. The biological replicates correlated closely with each other (un-der the same branch), and the lists of differentially expressed genes were highly comparable (data not shown). In addition, we fixed E12.5 as a develop-mental baseline, to which all the subse-quent time points were compared. The lists of differentially expressed genes generated by our analyses became larger as the timepoints became moreFig. 1. Histological samples of epidermal morphogenesis.A:From E12.5–E14, the epidermis is mainly composed of a single epithelial layer and an overlying periderm (e.g., E13.5 in A).B,C: E14.5–E15.5 morphological stratification and differentiation follows the earlier molecular changes (transcriptional program in this study). The hair follicle placodes and germs are also evident (C, E15.5 asterisk). D: A histological example of enzymatically and mechanically separated epidermis (top) and dermis (bottom) at E14.5 showing the two discrete compartments.
Fig. 2. Dmkn, Krtdap, andSbsntranscripts are highly induced in the suprabasal epidermis during epidermal morphogenesis.A:DmknandKrtdap, which are, unlikeSbsn, represented on MOE430A chips, are very highly induced (compared to baseline levels at E12.5) at the mRNA level as early as E13.5 to reach more than 1,900-fold by E15.5. Please note the difference in scale on the Y-axis between this graph and subsequent ones. dpc, days post coitus.B:Semi-quantitative RT-PCR validations of the microarray results, in addition toSbsn, are highly representative of the microarray outcome (the numbers below).-actin is a loading control for this and subsequent RT-PCRs.C:qRT-PCR analyses of the three SSC genes also corroborate the microarray as well as the previous RT-PCR results.D–F:Sections of the whole-mount ISH at E15.5 (see Fig. 6) localize the mRNAs for all three genes exclusively to the suprabasal layers of the developing epidermis. Less intense signals were detected above the developing hair follicle placodes (asterisk in E). Dotted line designates the epidermal-dermal junction. Scale bars (D–F)⫽40m.
the Epidermis From E12.5–E15.5a
SSC
genes 12.5 13.5 14 14.5 15.5 Dmkn 0 36 220 1,105 2,484 Krtdap 0 56 237 636 1,914
a0, Baseline levels or no change;
developmentally distant from the base-line (e.g., E12.5 vs. E13.5 compared to E12.5 vs. E15.5). This result reflects the gradual accumulation of transcriptional changes due to the addition of newly
differentiated populations of keratino-cytes as epidermal morphogenesis progresses. We categorized these lists of differentially expressed genes based on gene ontology.
Dmkn
and
Krtdap
Are the
Highest Differentially
Expressed Genes on the
Microarray
The two most highly differentially ex-pressed transcripts at all time points studied were consistently those of DmknandKrtdap(Table 1, Fig. 2A). The transcript levels increased sharply as early as E13.5 (36- to 56-fold), continuing through E14 –E14.5 (200 to more than 1,000-fold), and re-main high at E15.5 (1,900- to more than 2,400-fold) (Table 1, Fig. 2A). Sbsn, which is located in the same SSC cluster as Dmkn andKrtdap, is not represented by a probe on MOE430A arrays used in our study. The microarray output forDmknand Krtdap, as well asSbsntranscript ex-pression, was assessed and/or vali-dated by semi-quantitative RT-PCR (Fig. 2B) as well as quantitative real time PCR (Fig. 2C). Our results show that Sbsn mRNA also gradually in-creases, although it lags slightly be-hind those ofDmknandKrtdap, dur-ing epidermal morphogenesis until E14.5 (Fig. 2B,C). Moreover, as re-ported elsewhere in several studies, Dmkn, Krtdap, andSbsnmRNAs are localized exclusively to the suprabasal layers of the developing epidermis, where Sbsn transcripts are more prominent in the developing granular layer (E15.5, Fig. 2D,E) (Oomizu et al., 2000; Park et al., 2002; Matsui et al., 2004; Moffatt et al., 2004; Tsuchida et al., 2004).
Genes Involved in Adhesion,
Communication,
Transcription, as Well as
Enzymes, Are Among the
Differentially Expressed
Groups of Genes
Our lists of differentially expressed genes during epidermal morphogen-esis included genes that are well known to play a role during epider-mal keratinocyte differentiation (Ta-ble 2, Fig. 3A,B). For example, genes encoding cell– cell communication junctions and adhesion molecules such as desmocollins (Dsc1,3), S100 proteins, and aquaporins (Aqp3), showed prominent changes during epidermal development (Table 2, TABLE 2. Genes Involved in Communication/Adhesion and
Transcription as Well as Enzymes and Others Are Differentially Expressed During Epidermal Morphogenesisa
12.5 13.5 14 14.5 15.5 Communication/adhesion Dsc1 0 0 0 11 130 Dsc3 0 0 0 4 10 Pkp1 0 0 2 4 9 Jup 0 0 0 2 5 Eppk 0 0 0 4 8 Evp 0 0 0 3 13 Ppl 0 0 0 2 6 Cdh4 0 0 ⫺2 ⫺2 0 Cdh13 0 0 ⫺2 ⫺5 ⫺10 Pcdh8 0 0 0 ⫺9 ⫺14 Pcdh10 0 4 10 0 0 S100A18 0 0 0 0 177 S100A6 0 0 0 4 33 S100A14 0 4 6 15 27 Aqp3 0 3 20 52 162 Transcription factors Klf4 0 5 7 13 37 Klf5 0 0 0 3 7 Klf6 0 0 0 3 4 Cebp␣ 0 0 4 8 10 Cebp 0 3 4 9 12 Fos 0 0 0 0 10 Jun 0 0 2 0 3 Junb 0 0 0 0 4 Jdp2 0 ⫺5 ⫺5 ⫺5 ⫺11 Dlx5 0 7 13 16 39 Ovol1 0 0 3 6 10 Irx6 0 0 0 ⫺7 ⫺43 Enzymes/Miscellaneous Tgm1, K 0 0 0 3 5 Tgm1, E 0 0 0 3 11 Serpinb5 0 0 2 8 83 Serpinb2 0 0 0 0 54 Serpinb3c 1 3 12 27 67 Serpinb11 0 0 0 0 22 Elov1 1 3 7 47 186 Lgals3 0 0 4 64 559 Lgals7 0 0 2 10 61 Lgals12 0 0 0 0 5 Lmna 0 3 6 12 18 Calm4 0 0 0 12 1,066 Casp14 0 0 0 0 55 p21 0 0 0 3 14 Calb1 0 ⫺4 ⫺10 ⫺9 ⫺12 Rdh12 0 0 4 25 88 Vav3 0 14 21 20 21
a0, Baseline levels or no change; ⫺, down-regulation; numbers, fold of baseline
(E12.5).
Fig. 3A). This was also true for genes encoding transcription factors that are known to regulate gene tran-scription in the epidermis (AP-1 fac-tors, Cebp␣/, Klf4) (Table 2, Fig. 3A). In addition, genes encoding en-zymes that cross-link the compo-nents of the cornified envelope (Tgm1) and other enzymes and their inhibitors (Serpins) were also differ-entially expressed (Table 2, Fig. 3B). Finally, our microarray lists uncov-ered some differentially regulated genes whose function in the develop-ing epidermis is yet to be determined (Rdh12, Calb1) (Table 2, Fig. 3B). We chose a representative subset of these genes and validated their ex-pression by semi-quantitative RT-PCR (Fig. 3C).
Krt77
mRNA Is Highly
Expressed Among Other
Keratins in the Epidermis
During Morphogenesis
Since keratins are the hallmarks of epidermal keratinocytes, they were also abundantly differentially ex-when the first signs of spinous differentiation are evident.B:Transcripts of enzymes for cross-linking the future cornified envelope (Tgm1, E, andKare two polypeptides), enzymes involved in retinoic acid biosynthesis (Rdh12), cytoskeletal regulators (Vav3), and nuclear intermediate filaments (Lmna) are induced during early epidermal differentiation.C:RT-PCR validation of selected genes (A,B) corroborate the levels of induction in the microarray (below).
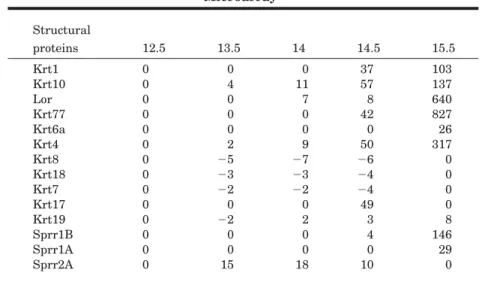
TABLE 3. The Structural Proteins That Are Hallmarks of Epidermal Keratinocytes Differentiation Are Differentially Expressed on the
Microarraya Structural proteins 12.5 13.5 14 14.5 15.5 Krt1 0 0 0 37 103 Krt10 0 4 11 57 137 Lor 0 0 7 8 640 Krt77 0 0 0 42 827 Krt6a 0 0 0 0 26 Krt4 0 2 9 50 317 Krt8 0 ⫺5 ⫺7 ⫺6 0 Krt18 0 ⫺3 ⫺3 ⫺4 0 Krt7 0 ⫺2 ⫺2 ⫺4 0 Krt17 0 0 0 49 0 Krt19 0 ⫺2 2 3 8 Sprr1B 0 0 0 4 146 Sprr1A 0 0 0 0 29 Sprr2A 0 15 18 10 0
a0, Baseline levels or no change; numbers, fold of baseline (E12.5); -, down-regulation.
Fig. 4. Well-established and novel structural proteins are highly expressed during epidermal morphogenesis.A:A graph showing the kinet-ics of mRNA expression of the well-studied ker-atin pair,Krt1and10, and the cornified enve-lope protein,loricrin, as well as the embryonic keratins,Krt6aandKrt77, during epidermal dif-ferentiation. Note the high expression levels of Krt77mRNA compared to the others by E15.5. A:RT-PCR data reflect data from the microar-ray (below).C:Indirect immunofluorescence of K8 and K18 shows their persistence in the per-iderm (E15.5) and the appearance of K18 in the basal layer and hair follicle (see text). DAPI nu-clear counterstain is in blue. The dotted line represents epidermal-dermal junction. Scale bars (C)⫽60m.
Fig. 5.
Fig. 6.
Fig. 7. The initiation sites of SSC gene ex-pression coincide with the initiation sites of bar-rier formation.A: The initiation sites for SSC gene mRNA expression, e.g.,Dmkn, are indi-cated in the specialized hair follicles around the eye and the nasal region in the inset (arrows).B: Using the well-established -galactosidase barrier assay, these same initiation sites of SSC expression are also initiation sites of barrier formation and remain devoid of a blue signal (arrows).
Fig. 5. Whole mount ISH of using a probe againstKrt77mRNA shows expression in the suprabasal layers of the differentiating epider-mis during morphogenesis.A:No signal is de-tected at E14.5 (even in the nasal epithelium whereKrt1andloricrinmRNAs are expressed). B:At E15,Krt77mRNA is detected in the whis-ker pad (also distinct fromKrt1mRNA expres-sion (see Results section) (Byrne et al., 1994), behind the ear, in some specialized hair follicles around the eye, in the palmar elevations (E), and some skin areas (armpit region). C,D,F: By E15.5–E16.5,Krt77transcripts are detected in wider areas of the skin until covering the entire body even the palmar skin (F). Plantar skin shows the same kinetics of expression but at later stages than palmar skin (not shown).G: Sections of whole mount ISH at E16.5 localize Krt77mRNA exclusively to the developing su-prabasal layers of the epidermis. Dotted line, epidermal-dermal junction. Scale bars (G)⫽40 m.
Fig. 6.Whole mount ISH forDmkn, Krtdap, and SbsnmRNA reveals that these genes are coor-dinately expressed during epidermal morpho-genesis.A–C:DmknmRNA is first detected in the nasal epithelium at E14 (shown E14.5, arrow in A and inset), then detected in larger areas of the skin as development progresses (B,C). The signals at the top of the head in B are artifacts due to the holes created in that region to pre-vent trapping of the probe in the brain during the ISH procedure.D–F:KrtdapmRNA is also first detected in the nasal epithelium at E14.5 (arrow in D, inset), and also at the proximal edge of the whisker pad (perhaps this embryo is⬃2 hr more advanced than that ofDmknin A); and at E15.5 (E) and E16.5 (F) it is indistinguishable from Dmkn pattern of expression. G–I:Sbsn mRNA expression pattern is identical to those ofDmknandKrtdapreflecting the coordination of their expression in a spatio-temporal fashion. J,K:Sections of theDmknISH at E15 through the nostril region localizes the mRNA to the stratifying nasal epithelium (arrows). K repre-sents the inset in J. Scale bars⫽(J) 160m, (K) 80m.
pressed during epidermal morphogen-esis (Table 3, Fig. 4A). Whereas the transcripts for the simple epithelial keratins such as Krt7,8and18 were down-regulated as early as E13.5, those of the spinous layer keratins such as the keratin pairKrt1and10 were concomitantly up-regulated (Ta-ble 3, Fig. 4A,B). Interestingly, Krt8 and Krt18 mRNA slightly increase again by E15.5 (Fig. 4B). In this re-spect, K8 expression is maintained in the periderm and is present in a dis-crete layer of the developing hair fol-licle by E16.5 (Fig. 4C, and data not shown), while K18 expression is ex-tended to the basal layer as well as the developing hair follicles by E15.5 and decreases in expression again by E16.5 (Fig. 4C, data not shown). These expression data are in accordance with the changes in their transcript levels (Fig. 4A,B). These changes in keratin expression were accompanied by the up-regulation of loricrin mRNA, a granular layer marker and a cornified envelope component, which was increased up to 640-fold by E15.5 (Table 3, Fig. 4A). Moreover, the mu-cosal type keratin, Krt4, whose ex-pression is undetectable in normal adult epidermis but is up-regulated under abnormal conditions (Virtanen et al., 2000), was also highly up-regu-lated during epidermal morphogene-sis (Table 3, Fig. 4A,B).
Notably, the most highly expressed keratin was a largely uncharacterized type II basic embryonic type keratin (Hesse et al., 2001, 2004; Rogers et al., 2005; Senshu et al., 2005). This kera-tin is known by several different names in the literature including ker-atin complex 2, basic, gene 39 (Kb39), Krt 77, K1-embryonic, and mouse K1b (mK1b) (Hesse et al., 2001, 2004; Rog-ers et al., 2005).Krt77mRNA, accord-ing to the new keratin nomenclature (Schweizer et al., 2006), increased steeply at E14.5 (42-fold) and reached 827-fold at E15.5, higher than any other keratin or structural protein transcript on our lists (Table 3, Fig. 4A).Krt6amRNA, another mouse em-bryonic type keratin gene, also in-creased sharply by E15.5 (26-fold) (Ta-ble 3, Fig. 4A). The differential expression of Krt77, Krt1, and Krt4 transcripts were validated by RT-PCR (Fig. 4B).
Krt77
mRNA Is Expressed in
the Suprabasal Layers of the
Developing Epidermis
The human orthologue of Krt77, KRT77, or K1b is expressed at both the mRNA and protein levels exclu-sively in the luminal cells of the sweat gland ducts of adult human skin (Langbein et al., 2005). Therefore, we wished to localize the expression of Krt77 mRNA during epidermal mor-phogenesis. Using whole mount ISH, we did not detect Krt77 mRNA at E14.5 (Fig. 5A). However, at E15 we detected Krt77 mRNA in the region surrounding the vibrissae follicles in the whisker pad, behind the pinnae of the ears, around a developing special-ized hair follicle in the snout region, in the skin under the forelimbs, and in the developing palmar skin (Fig. 5B,E). Some of these initiation sites of expression were distinct from those reported for Krt1, especially the ab-sence of signals from the nasal region and the vibrissae follicles hair canals, arguing against the cross-reactivity of theKrt77probe withKrt1mRNA (By-rne et al., 1994). By E15.5, Krt77 mRNA could be detected in a wider area of the lateral skin, limbs, and tail (Fig. 5C). By E16.5,Krt77mRNA ex-pression covered essentially the whole embryo including the entire palmar skin (Fig. 5D,F). Sections from these E16.5 embryos revealed that Krt77 mRNA was exclusively and promi-nently expressed in the developing su-prabasal layers of the epidermis (Fig. 5G).
Dmkn, Krtdap
, and
Sbsn
Are
Coordinately Expressed
During Epidermal
Morphogenesis
SinceDmknandKrtdap had the two most highly expressed transcripts on our microarray, we were interested in determining their expression kinet-ics during epidermal morphogenesis. Therefore, we used whole mount ISH to analyze the expression of Dmkn, Krtdap, and Sbsn during develop-ment. Intriguingly, we detected the mRNA of all three genes in the nasal region as early as E14 and E14.5 (data not shown, and Fig. 6A,D,G, arrows and insets). Sections from this region revealed that the mRNA is localized to
the suprabasal layers of the nasal ep-ithelium within the nostril (e.g., Dmkn, Fig. 6J,K). A control sense probe for each of these transcripts showed no signal at this time point or subsequent ones (data not shown). By E15.5, the mRNA signals for all three genes covered almost the entire body excluding the scalp region, which is covered by E16.5 (Fig. 6B,C,E,F,H,I). It is noteworthy that the whole mount expression patterns ofDmkn, Krtdap, andSbsnare virtually indistinguish-able at all time points studied.
The Initiation of Expression
of SSC Genes Correlates
With Barrier Induction in
the Nasal Epithelium
During our analysis of the expression patterns ofDmkn, Krtdap, and Sbsn, we noted that the initiating site of mRNA expression for these genes is the nasal epithelium (Figs. 6A,D,G,J,K, 7A). Moreover, by E15 the mRNA for these SSC genes is de-tected in the three developing hair fol-licles around the eye and near the mouth (Fig. 7A). Interestingly, these sites of expression correspond to the sites of initiation of barrier formation in the mouse embryo, which occurs⬃4 days later⬃E17.5 (Fig. 7B) (Hardman et al., 1998).
DISCUSSION
The transformation of the mamma-lian surface ectoderm from a simple epithelium to a complex epidermis is not completely understood. There is evidence correlating heterogeneity in the underlying mesenchyme and gene expression in the ectoderm, suggest-ing that the mesenchyme could pro-vide the earliest cues for the transfor-mation of the epidermis as early as E8.5 (Sengel, 1986). Early markers of commitment to this complex transfor-mation, along with periderm forma-tion, include the transcription factor TAp63 and its downstream target Krt14(Koster and Roop, 2004; Lechler and Fuchs, 2005; Laurikkala et al., 2006). The subsequent stratification and differentiation events that lead to the formation of the spinous layer ⬃E15.5 are other hallmarks during epidermal morphogenesis (Byrne et al., 1994; Koster and Roop, 2004;
Lechler and Fuchs, 2005; Laurikkala et al., 2006).
In this study, we were interested in identifying novel factors expressed around the time of epidermal morpho-genesis and differentiation, E12.5– E15.5, during the formation of the suprabasal layers of the mature epi-dermis. The clean separation of devel-oping epidermal and mesenchymal compartments as early as E12.5 re-sulted in a reliable list of differentially expressed transcripts during epider-mal morphogenesis (e.g., E14.5 in Fig. 1D). The abundance of well-studied proteins in the developing and mature epidermis, ranging from structural, enzymatic, communication, transcrip-tional, and others, provided strong in-ternal controls, lending validity to the technical procedures as well as com-putational analysis methods (Tables 1–3; Figs. 2– 4). The RT-PCR valida-tions of selected genes also corrobo-rate the differential gene expression obtained from the microarray analy-ses (Figs. 2B,C. 3C, 4B).
Intriguingly, the newly identified genes of the SSC cluster encoding se-creted proteins, Dmkn and Krtdap, were the most highly differentially ex-pressed genes as early as E13.5, and as high as 2,400-fold by E15.5 (Table 1, Fig. 2A–C). The third gene in the SSC cluster,Sbsn, which is not repre-sented on the MOE430A array used in our study, was also gradually and highly induced by E15.5 as deter-mined by RT-PCR and qRT-PCR (Fig. 2B,C). The expression of these genes in the epidermis as well as other strat-ified epithelia has been shown to be maintained in adult tissues (Oomizu et al., 2000; Park et al., 2002; Matsui et al., 2004; Moffatt et al., 2004; Tsuchida et al., 2004). Since these pro-teins are secreted and their mRNAs are expressed exclusively in the su-prabasal layers of the developing epi-dermis (Fig. 2D–F), and that their ex-pression levels surpass any other induced transcripts in the epidermis, are all indications that they play a significant role during early epider-mal differentiation (Matsui et al., 2004; Moffatt et al., 2004). Applying the same line of reasoning to the em-bryonic keratin Krt77 (827-fold by E15.5 and also expressed in the supra-basal epidermis), this Krt1 homolog represents a novel marker for early
epidermal differentiation. The differ-ence in the initiation sites of expres-sion ofKrt77(which is not expressed in the nasal epithelium) compared to Krt1suggests a complementary role of Krt77to that ofKrt1during epidermal morphogenesis (Fig. 5B).
The coordinated spatial and tempo-ral expression patterns of the SSC genes are striking and remarkable (Fig. 6A–I). As mentioned earlier, these genes are clustered in a rela-tively small region of the genome (⬃40 Kb on human chromosome 19 and mouse chromosome 7) and they are transcribed in the same orientation (Matsui et al., 2004; Moffatt et al., 2004). Collectively, these observations suggest an interesting mode of coordi-nated transcriptional regulation in the differentiating epidermis that is worthy of further investigation. We analyzed the upstream regions of each of the SSC genes as well as of Krt1, Krt10, andLor focusing on the tran-scription factors that showed differen-tial expression on our microarray (Ta-ble 2). We found a significant number of consensus binding sites for Cebp␣, AP-1 factors (mainly c-Fos and c-Jun), and AP-2 factors (mainly AP-2␣) within⬃1Kb of the upstream regions of these genes. Combinations of these factors have been shown to be essen-tial for regulating the expression of Krt1, Krt10, Lor, andinvolucrin (May-tin et al., 1999; Zhu et al., 1999; Jang and Steinert, 2002; Eckert et al., 2004). In this regard, in vitro experi-ments on keratinocytes (human or mouse) using differentiating agents such as calcium and phorbol esters (TPA) have shown that the induction of differentiation markers (including Krt1/10, Lor, Dmkn, Krtdap, Sbsn) op-erate through transcriptional regula-tors of the AP and Cebp families (Hen-nings et al., 1980; Maytin et al., 1999; Zhu et al., 1999; Jang and Steinert, 2002; Eckert et al., 2004). We repro-duced these findings for Dmkn and Sbsn using Calcium and TPA treat-ment of primary mouse epidermal keratinocytes (data not shown). Therefore, Cebp␣, AP-1, and AP-2 fac-tors are potential candidates for the regulation of expression of SSC genes, in addition to the aforementioned structural proteins.
Of particular interest is the initia-tion of mRNA expression of the SSC
genes in the nasal epithelium (E13.5– E14), which is reminiscent of mRNA expression initiation sites forKrt1and Loras early as E13.5, indicating that the nasal epithelium is the earliest site in the epidermis that under-goes stratified differentiation (Figs. 6A,D,G,J,K, 7A) (Byrne et al., 1994). Not only is the nasal epithelium per-haps the earliest epidermal differenti-ation site, but it is also likely to be the earliest to acquire a barrier⬃4 days later during epidermal development (Fig. 7B) (Hardman et al., 1998). The expression of the late constituents of the epidermal cornified envelope was previously correlated with barrier ini-tiation, albeit several days later in de-velopment than our study (Marshall et al., 2001). We believe that studying the early kinetics of differentiation of the nasal epithelium will shed more light on the links between stratifica-tion, differentiastratifica-tion, and barrier for-mation in the epidermis and perhaps the role of mesenchymal cues during these processes.
EXPERIMENTAL
PROCEDURES
Microarray Analysis
Dorsal skin was dissected from C57BL/6J embryos at 12-hr intervals starting at E12.5 (12.5 days post coi-tus, day of plug considered 0.5 days [d]; digit as well as whisker pad folli-cle formation were used as staging cri-teria) until E15.5 (refer to Fig. 1 for histology of selected stages), in DMEM with antibiotics and antimy-cotics (Invitrogen, Carlsbad, CA). The dissected skin was incubated in a 2:1 mixture of 2.5% trypsin (without EDTA) (Invitrogen) and filtered 8% pancreatin (Sigma-Aldrich, St Louis, MO) for 5 min at room temperature followed by 30 – 40 min at 4°C. The epidermis was carefully and cleanly separated from the dermis, and each tissue was resuspended and frozen in RLT buffer (Qiagen, Valencia, CA). Total RNA was isolated from the epi-dermal and epi-dermal tissues using the RNeasy威 Minikit according to the manufacturer’s instructions (Qiagen). Triplicate to quadruplicate RNA sam-ples (each biological sample often rep-resented a single embryo) were ampli-fied once and labeled for hybridization
on microarray chips (MOE430A) us-ing the Affymetrix reagents and pro-tocols (Affymetrix Inc., Santa Clara, CA). The data output was normalized and analyzed using both GeneSpring GX 7.0 (Agilent Technologies Inc., Palo Alto, CA) and GeneTraffic™ (Io-bion Informatics, La Jolla, CA) com-mercial software packages, which gave comparable results. The earliest time point, E12.5, was set as a base-line for comparison of subsequent stages. The Pvalue cutoff was set to 0.05 and the significant fold difference was considered two-fold higher or lower than baseline.
Semi-Quantitative RT-PCR
and Quantitative Real Time
PCR
Reverse transcription was carried out using Oligo (dT) primer and Super-Script™ III (Invitrogen) according to the manufacturer’s instructions. PCR was performed using PfuUltra™ Hot-start PCR Master Mix (Stratagene, La Jolla, CA) in a Peltier Thermal Cycler (MJ Research, Inc, Waltham, MA). The different cDNAs from each of the time points were equalized using -ac-tin primers as an internal control. Ali-quots of each PCR reaction were taken at 25, 30, and 35 cycles, run on a 1% agarose gel, and photographed using a Kodak camera. Quantitative real time PCR (qRT-PCR) was performed on an ABI 7300 (Applied Biosystems, Foster City, CA). Primers were designed ac-cording to ABI guidelines and all re-actions were performed using ABI Sybr Master Mix, 300-nM primers, 200 ng cDNA at the following consec-utive steps: (1) 50°C for 2 min, (2) 95°C for 10 min, (3) 95°C for 15 s, (4) 60°C for 1 min, repeat steps 3 and 4 for 40 cycles. The samples were run in quadruplicate and normalized to an internal control (Tubulin) using the accompanying software. The primers used and aplicon sizes are provided in Table S1 (see Supplemental Table S1, which can be viewed at www. interscience.wiley.com/jpages/1058-8388/suppmat).
Whole Mount In Situ
Hybridization
The templates for the in situ hybrid-ization probes were amplified by PCR
from the E15.5 epidermal cDNA stock (see RT-PCR section; the primers used and probe length are provided in Sup-plementary Table S1). The PCR prod-ucts were cloned into pCRII dual promoter (T7 and SP6) vector (Invitro-gen) and standard procedures were followed for the preparation of DIG-labeled cRNA (Roche Applied Science, Indianapolis, IN) antisense (AS) and control sense (S) probes. In situ hy-bridization was performed on differ-ent embryonic stages during epider-mal morphogenesis as per detailed published protocols (Wilkinson, 1998). The embryos were photographed us-ing an HRC Axiocam fitted onto an SV Stemi stereomicroscope (Carl Zeiss, Thornwood, NY). The stained embryos were post-fixed in 4% PFA, embedded in Tissue-Tek威 OCT compound (Fisher Scientific, Hampton, NH), sec-tioned, and mounted for histological photography.
Immunofluorescence Staining
Frozen sections of whole embryos were fixed in 4% PFA at ambient tem-perature followed by acetone at ⫺20°C. The sections were washed in 1⫻PBS, blocked in 10% of the appro-priate serum, and incubated with the primary antibody overnight at 4°C. After washing in 1⫻PBS, 594 Alex-afluor威conjugated secondary antibod-ies (Molecular Probes, Invitrogen) were applied and the signal was visu-alized using an HRC Axiocam fitted onto an Axioplan2 fluorescence micro-scope (Carl Zeiss). Guinea pig antibod-ies against K8 (1:2,000) and K18 (1: 1,000) were kindly provided by Dr. Lutz Langbein.Barrier Assay and In Silico
Promoter Analysis
The barrier assay monitors endoge-nous skin -galactosidase activity. The X-Gal substrate can penetrate the skin only where the barrier has not formed yet, and therefore, a blue color develops. This assay was performed as described previously (Hardman et al., 1998). For the analysis of the up-stream regions of SSC cluster genes as well as Krt1/10, we used the VISTA website for comparative genomics (http://genome.lbl.gov/vista/index. shtml) (Loots et al., 2002), in addition
to the AliBaba2.1 tool on the Gene-Regulation website (http://www. gene-regulation.com/pub/programs/ alibaba2/index.html?).
ACKNOWLEDGMENTS
We thank Dr. Yonghui Zhang from the Columbia Genome Center for provid-ing advice on the microarray analysis softwares. We thank Mr. Ming Zhang for excellent technical assistance. We also thank all the members of the Christiano and Jahoda laboratories for helpful assistance and discussion. This work was supported by a grant from the Kirsch Foundation to A.M.C. C.A.B.J. is grateful to the BBSRC for funding (grant Number G18988).
REFERENCES
Byrne C, Tainsky M, Fuchs E. 1994. Pro-gramming gene expression in developing epidermis. Development 120:2369 –2383. Byrne C, Hardman M, Nield K. 2003. Cov-ering the limb—formation of the integu-ment. J Anat 202:113–123.
Eckert RL, Crish JF, Efimova T, Dashti SR, Deucher A, Bone F, Adhikary G, Huang G, Gopalakrishnan R, Balasubra-manian S. 2004. Regulation of involucrin gene expression. J Invest Dermatol 123: 13–22.
Fuchs E, Green H. 1980. Changes in kera-tin gene expression during terminal dif-ferentiation of the keratinocyte. Cell 19: 1033–1042.
Hardman MJ, Sisi P, Banbury DN, Byrne C. 1998. Patterned acquisition of skin barrier function during development. Development 125:1541–1552.
Hennings H, Michael D, Cheng C, Steinert P, Holbrook K, Yuspa SH. 1980. Calcium regulation of growth and differentiation of mouse epidermal cells in culture. Cell 19:245–254.
Hesse M, Magin TM, Weber K. 2001. Genes for intermediate filament proteins and the draft sequence of the human ge-nome: novel keratin genes and a surpris-ingly high number of pseudogenes re-lated to keratin genes 8 and 18. J Cell Sci 114:2569 –2575.
Hesse M, Zimek A, Weber K, Magin TM. 2004. Comprehensive analysis of keratin gene clusters in humans and rodents. Eur J Cell Biol 83:19 –26.
Jang SI, Steinert PM. 2002. Loricrin ex-pression in cultured human keratino-cytes is controlled by a complex interplay between transcription factors of the Sp1, CREB, AP1, and AP2 families. J Biol Chem 277:42268 –42279.
Koster MI, Roop DR. 2004. p63 and epithe-lial appendage development. Differenti-ation 72:364 –370.
Koster MI, Kim S, Mills AA, DeMayo FJ, Roop DR. 2004. p63 is the molecular switch for initiation of an epithelial
stratification program. Genes Dev 18: 126 –131.
Langbein L, Rogers MA, Praetzel S, Cri-bier B, Peltre B, Gassler N, Schweizer J. 2005. Characterization of a novel human type II epithelial keratin K1b, specifi-cally expressed in eccrine sweat glands. J Invest Dermatol 125:428 –444. Laurikkala J, Mikkola ML, James M,
Tum-mers M, Mills AA, Thesleff I. 2006. p63 regulates multiple signalling pathways required for ectodermal organogenesis and differentiation. Development 133: 1553–1563.
Lechler T, Fuchs E. 2005. Asymmetric cell divisions promote stratification and dif-ferentiation of mammalian skin. Nature 437:275–280.
Loots GG, Ovcharenko I, Pachter L, Dub-chak I, Rubin EM. 2002. rVista for com-parative sequence-based discovery of functional transcription factor binding sites. Genome Res 12:832–839.
Marshall D, Hardman MJ, Nield KM, By-rne C. 2001. Differentially expressed late constituents of the epidermal cornified envelope. Proc Natl Acad Sci USA 98: 13031–13036.
Matsui T, Hayashi-Kisumi F, Kinoshita Y, Katahira S, Morita K, Miyachi Y, Ono Y, Imai T, Tanigawa Y, Komiya T, Tsukita S. 2004. Identification of novel keratino-cyte-secreted peptides dermokine-al-pha/-beta and a new stratified epitheli-um-secreted protein gene complex on human chromosome 19q13.1. Genomics 84:384 –397.
Maytin EV, Lin JC, Krishnamurthy R, Batchvarova N, Ron D, Mitchell PJ, Ha-bener JF. 1999. Keratin 10 gene expres-sion during differentiation of mouse
epi-dermis requires transcription factors C/EBP and AP-2. Dev Biol 216:164 –181. Mischke D, Korge BP, Marenholz I, Volz A, Ziegler A. 1996. Genes encoding struc-tural proteins of epidermal cornification and S100 calcium-binding proteins form a gene complex (“epidermal differentia-tion complex”) on human chromosome 1q21. J Invest Dermatol 106:989 –992. Moffatt P, Salois P, St-Amant N, Gaumond
MH, Lanctot C. 2004. Identification of a conserved cluster of skin-specific genes encoding secreted proteins. Gene 334: 123–131.
Oomizu S, Sahuc F, Asahina K, Inamatsu M, Matsuzaki T, Sasaki M, Obara M, Yoshizato K. 2000. Kdap, a novel gene associated with the stratification of the epithelium. Gene 256:19 –27.
Park GT, Lim SE, Jang SI, Morasso MI. 2002. Suprabasin, a novel epidermal dif-ferentiation marker and potential corni-fied envelope precursor. J Biol Chem 277: 45195–45202.
Rogers MA, Edler L, Winter H, Langbein L, Beckmann I, Schweizer J. 2005. Char-acterization of new members of the hu-man type II keratin gene family and a general evaluation of the keratin gene domain on chromosome 12q13.13. J In-vest Dermatol 124:536 –544.
Rothnagel JA, Longley MA, Bundman DS, Naylor SL, Lalley PA, Jenkins NA, Gilbert DJ, Copeland NG, Roop DR. 1994. Characterization of the mouse loricrin gene: linkage with profilaggrin and the flaky tail and soft coat mutant loci on chromosome 3. Genomics 23:450 – 456.
Schweizer J, Bowden PE, Coulombe PA, Langbein L, Lane EB, Magin TM, Mal-tais L, Omary MB, Parry DA, Rogers
MA, Wright MW. 2006. New consensus nomenclature for mammalian keratins. J Cell Biol 174:169 –174.
Segre J. 2003. Complex redundancy to build a simple epidermal permeability barrier. Curr Opin Cell Biol 15:776 –782. Sengel P. 1976. Morphogenesis of skin.
Cam-bridge: Cambridge University Press. Sengel P. 1986. Epidermal-Dermal
Inter-action. Berlin: Springer-Verlag. Senshu T, Ishida-Yamamoto A, Takahashi
H, Iizuka H. 2005. Prediction of a coding sequence for a novel type II keratin from N-terminal sequences of mouse epider-mal proteins site-specifically deiminated in embryonic development. J Dermatol Sci 37:41–48.
Tsuchida S, Bonkobara M, McMillan JR, Akiyama M, Yudate T, Aragane Y, Te-zuka T, Shimizu H, Cruz PD Jr, Ari-izumi K. 2004. Characterization of Kdap, a protein secreted by keratinocytes. J In-vest Dermatol 122:1225–1234.
Virtanen M, Torma H, Vahlquist A. 2000. Keratin 4 upregulation by retinoic acid in vivo: a sensitive marker for retinoid bioactivity in human epidermis. J Invest Dermatol 114:487–493.
Volz A, Korge BP, Compton JG, Ziegler A, Steinert PM, Mischke D. 1993. Physical mapping of a functional cluster of epider-mal differentiation genes on chromo-some 1q21. Genomics 18:92–99. Wilkinson D. 1998. In situ hybridization: a
practical approach. Oxford: Oxford Uni-versity Press.
Zhu S, Oh HS, Shim M, Sterneck E, John-son PF, Smart RC. 1999. C/EBPbeta modulates the early events of keratino-cyte differentiation involving growth ar-rest and keratin 1 and keratin 10 expres-sion. Mol Cell Biol 19:7181–7190. 970 BAZZI ET AL.