MECHANISMS OF ANTIBIOTIC RESISTANCE IN NEISSERIA GONORRHOEAE AND THE DEVELOPMENT OF LPXC INHIBITORS AS NOVEL THERAPEUTICS
Shauna Marie Swanson
A dissertation submitted to the faculty of the University of North Carolina at Chapel Hill in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the
Department of Microbiology and Immunology
Chapel Hill 2014
Approved by:
Robert A. Nicholas
Miriam Braunstein
Marcia M. Hobbs
Anthony R. Richardson
Matthew C. Wolfgang
© 2014
Shauna Marie Swanson ALL RIGHTS RESERVED
ABSTRACT
Shauna Marie Swanson: Mechanisms of antibiotic resistance in N. gonorrhoeae and the development of LpxC inhibitors as novel therapeutics
(Under the direction of Robert A. Nicholas)
Neisseria gonorrhoeae is a Gram-‐negative diplococcus that causes the sexually
transmitted infection gonorrhea. Antibiotics are required to treat gonorrhea.
Unfortunately, the gonococcus has developed resistance to all classes of antibiotics that have been used to treat gonorrhea. Antibiotics primarily diffuse through the outer
membrane porin channels into the periplasm, but we have shown that they can also access the periplasm via the outer membrane secretin PilQ; mutation or deletion of PilQ increases resistance to penicillin by approximately threefold. In its mature form, PilQ exists as an SDS-‐resistant multimer. However, immature PilQ monomers are also present in the outer membrane. We initiated studies to determine if antibiotic permeation occurred through the mature secretin or via immature PilQ complexes. Our data indicate that immature, SDS-‐ labile PilQ complexes form channels that allow antibiotics to traverse the outer membrane. To help meet the growing need for new antibiotics for the treatment of gonorrhea, we have validated the enzyme LpxC as a drug target in N. gonorrhoeae. LpxC catalyzes the first committed step in the biosynthesis of lipid A, a major component of the outer
tested in vivo using the murine model of gonorrhea, and one of these inhibitors was able to clear the infection in eight out of nine infected mice.
We also evaluated the capacity of N. gonorrhoeae to develop resistance to LpxC inhibitors. Spontaneously arising mutants displayed a 4-‐ to 16-‐fold decrease in
susceptibility to LpxC inhibitors, and in several of these mutants, mutation or deletion of the pqiA and pqiB genes was responsible for the resistance phenotype. The functions of PqiA and PqiB are unknown, but the loss of pqiAB does not alter the MICs of other
antibiotics and does not affect LpxC expression or growth rate. The results of the studies described within this dissertation further our understanding of antibiotic resistance mechanisms in N. gonorrhoeae and provide the basis for the development of LpxC inhibitors as potential new therapeutics for the treatment of gonorrhea.
To my family, especially my parents, for supporting me throughout college and graduate
school. You did not always understand what I was working on, but you always believed in me.
ACKNOWLEDGEMENTS
I would like to thank all the people at UNC who have helped me become a better scientist and have supported me throughout graduate school. Rob welcomed me into his lab and provided guidance and patience. However, this work would not have been possible without the help of generous coworkers. Sobhan Nandi told me “one thing” (actually, many, many things) about gonorrhea and helped me find my feet in the lab. Josh Tomberg taught me about proteins (and that you can microwave stir bars without causing an explosion) and helped me troubleshoot whenever cloning was “broken”. Meg Duncan, Kate White, Tasha Blatt, and Swati Rayasam provided extra sets of hands on crazy days, lots of giggles about inside jokes, and sympathetic ears when projects were not going as planned. I’d also like to thank my friends and family for always being interested in hearing about whatever I was doing in graduate school. When you’re surrounded by people who have their PhD or are working on it, it is easy to forget what an accomplishment it truly is. Thank you for always encouraging and believing in me. Without you reminding me that there is life outside of graduate school, I might not have made it all the way through!
TABLE OF CONTENTS
LIST OF FIGURES ... xii
LIST OF TABLES ... xiv
LIST OF ABBREVIATIONS ... xv
Chapter 1. Introduction ... 1
1.1 Introduction ... 1
1.2 Public heath relevance ... 2
1.2.1 Epidemiology ... 2
1.2.2 Clinical manifestations ... 4
1.2.3 Gonorrhea and HIV co-‐infections ... 5
1.3 Structural components of the gonococcal cell wall ... 6
1.3.1 Lipooligosaccharide ... 7
1.3.2 Type IV pili ... 11
1.3.3 Opacity-‐associated proteins ... 16
1.3.4 Peptidoglycan ... 17
1.3.5 Porin ... 18
1.4 Infection ... 20
1.4.1 Adhesion and invasion ... 20
1.4.2 Bacterial defenses against the host immune response ... 22
1.4.3 Murine model of gonococcal infection ... 24
1.5 Genetic transformation ... 26
1.5.1 Natural competence ... 26
1.5.3 DNA binding and uptake ... 28
1.5.4 DNA integration ... 29
1.6 Antibiotic resistance ... 30
1.6.1 History of antibiotic resistance ... 30
1.6.2 Plasmid-‐mediated antibiotic resistance ... 34
1.6.3 Chromosomally mediated antibiotic resistance ... 35
1.7 LpxC inhibitors as a novel class of antibiotics ... 38
1.7.1 Lipid A biosynthesis ... 38
1.7.2 History of LpxC inhibitors ... 43
1.8 Introduction to the dissertation ... 48
1.8.1 Contribution of PilQ to antibiotic resistance ... 48
1.8.2 Validation of gonococcal LpxC as a target for antibiotics ... 48
1.8.3 Characterization of a mutant with reduced susceptibility to LpxC inhibitors ... 49
Chapter 2. Diffusion of antibiotics through the PilQ secretin in Neisseria gonorrhoeae occurs through the immature, SDS-‐labile form ... 50
2.1 Overview ... 50
2.2 Introduction ... 52
2.3 Materials and Methods ... 56
2.3.1 Bacterial strains. ... 56
2.3.2 Bacterial media, growth conditions, and MIC determinations. ... 57
2.3.3 Transformations and transformation efficiency assays. ... 58
2.3.4 Isolation of spontaneously arising penicillin-‐resistant clones. ... 59
2.3.5 Scanning electron microscopy. ... 60
2.3.6 Construction of pilM, pilN, pilO and pilP frame-‐shift and in-‐frame deletion mutants. ... 60
2.3.8 Overexpression of PilQ. ... 62
2.3.9 Gel filtration. ... 63
2.3.10 SDS-‐PAGE and Western blotting. ... 64
2.4 Results ... 67
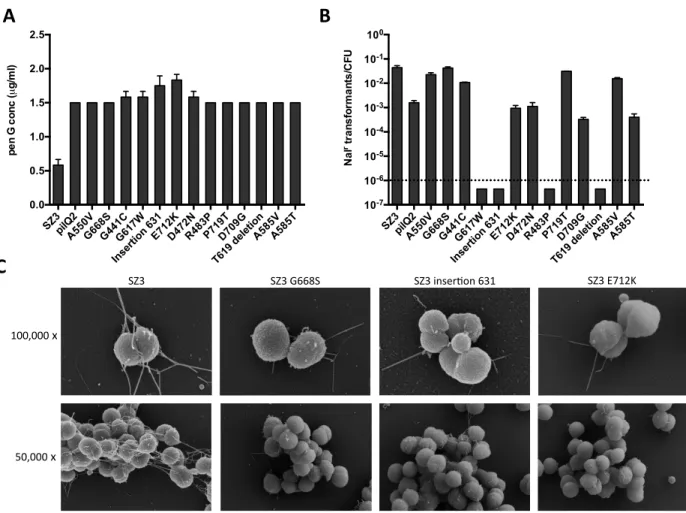
2.4.1 Phenotypic identification of spontaneously arising penicillin-‐resistant clones ... 67
2.4.2 Characterization of class 1 mutants ... 70
2.4.3 Analysis of class 2 mutants ... 74
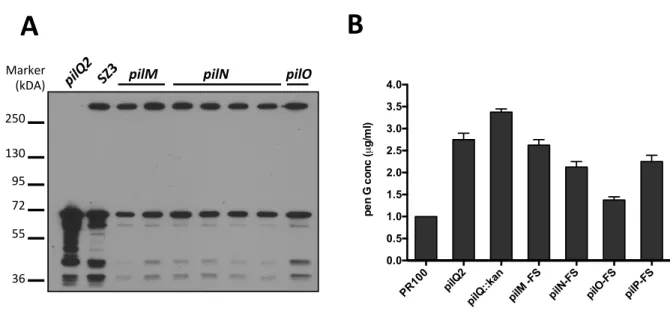
2.4.4 Phenotypes of the in-‐frame pilM, pilN, pilO and pilP deletions ... 77
2.4.5 Effects of titration of PilQ levels on penicillin resistance ... 81
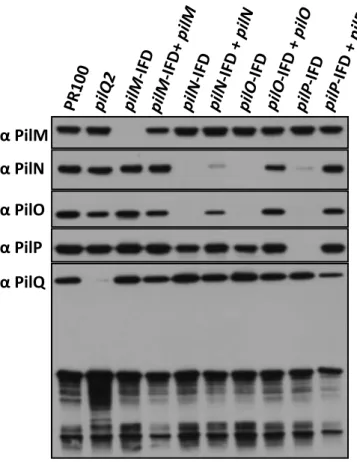
2.4.6 Analysis of PilQ levels in wild-‐type versus frame-‐shift mutants ... 83
2.4.7 Manipulation of PilW expression to regulate levels of the PilQ multimer ... 87
2.5 Discussion ... 91
Chapter 3. Diacetylene LpxC inhibitors as novel antibiotics with activity against multi-‐drug-‐resistant Neisseria gonorrhoeae ... 98
3.1 Overview ... 98
3.2 Introduction ... 100
3.3 Materials and Methods ... 103
3.3.1 Bacterial strains ... 103
3.3.2 Bacterial growth conditions ... 103
3.3.3 Antibiotic susceptibility testing ... 104
3.3.4 Bactericidal assay ... 105
3.3.5 Transformation of N. gonorrhoeae ... 105
3.3.6 Knockout and complementation of NgLpxC ... 106
3.3.7 Construction of strains expressing RlLpxC and EcLpxC ... 106
3.4.1 LpxC inhibitors are active against N. gonorrhoeae ... 109
3.4.2 LpxC is essential in N. gonorrhoeae ... 112
3.4.3 LpxC is the primary target of LpxC inhibitors in N. gonorrhoeae ... 114
3.4.4 Effect of existing resistance factors on susceptibility to LpxC inhibitors ... 117
3.4.5 LpxC inhibitors are active against ESC-‐resistant strains of N. gonorrhoeae ... 119
3.4.6 LpxC inhibitors display in vivo activity against N. gonorrhoeae ... 121
3.5 Discussion ... 124
Chapter 4. Deletion of pqiAB decreases the susceptibility of N. gonorrhoeae to LpxC inhibitors ... 126
4.1 Overview ... 126
4.2 Introduction ... 128
4.3 Materials and Methods ... 130
4.3.1 Bacterial strains and growth conditions ... 130
4.3.2 Isolation of spontaneous mutants ... 130
4.3.3 Susceptibility testing ... 131
4.3.4 Transformation of N. gonorrhoeae ... 131
4.3.5 Identification of the R7 mutation ... 131
4.3.6 Complementation of the R7 mutant ... 132
4.3.7 Gene disruption and knock-‐out ... 133
4.4 Results ... 134
4.4.1 Isolation and characterization of spontaneously arising mutants ... 134
4.4.2 Identification of the R7 mutation ... 138
4.4.3 Function of PqiA and PqiB ... 142
4.4.4 Requirement of both pqiA and pqiB for sensitivity to LpxC inhibitors ... 144
4.5 Discussion ... 148
Chapter 5. Discussion and future directions ... 151
5.1 Summary and significance ... 151
5.2 Pathogenic potential of pilQ mutants ... 153
5.3 LpxC overexpression ... 155
5.4 LpxC stability and regulation ... 158
5.5 Broad vs. narrow spectrum antibiotics ... 160
5.6 Future directions ... 161
REFERENCES ... 163
LIST OF FIGURES
Figure 1.1 Geographical distribution of gonorrhea infections across the United States ... 3
Figure 1.2. Structure of Kdo2-‐lipid A. ... 10
Figure 1.3 Type four pili of N. gonorrhoeae ... 14
Figure 1.4 Timeline of antibiotic resistance in N. gonorrhoeae ... 33
Figure 1.5 The lipid A biosynthesis pathway in E. coli. ... 42
Figure 1.6. LpxC inhibitors. ... 47
Figure 2.1 Western blots for PilQ from spontaneous arising penicillin-‐ resistant mutants of PR100. ... 69
Figure 2.2 Phenotypes of spontaneously arising pilQ mutants. ... 73
Figure 2.3 Effects of frame-‐shift mutations. ... 76
Figure 2.4 In-‐frame deletions of pilM, pilN, pilO and pilP affect transformation efficiency but not penicillin resistance. ... 79
Figure 2.5 Effect of pilM, pilN, pilO and pilP deletions on the stability of the proteins encoded by other genes in the operon. ... 80
Figure 2.6 Overexpression of PilQ increases susceptibility to penicillin. ... 82
Figure 2.7 The pilM frame-‐shift mutant decreases the levels of PilQ. ... 85
Figure 2.8 Gel filtration of FA19 membranes ... 86
Figure 2.9 Insertional inactivation of pilW. ... 89
Figure 2.10 Overexpression of PilW ... 90
Figure 3.1 LpxC inhibitors target the first committed step in the biosynthesis of lipid A. ... 102
Figure 3.2 Screening and optimization of LpxC inhibitors. ... 110
Figure 3.3 LpxC inhibitors are bactericidal for N. gonorrhoeae. ... 111
Figure 3.5 IPTG-‐inducible expression of LpxC. ... 115
Figure 3.6 LpxC is the primary target of LpxC inhibitors in N. gonorrhoeae. ... 116
Figure 3.7 Effect of known resistance factors on susceptibility to LpxC inhibitors. ... 118
Figure 3.8. Activity of LpxC inhibitors against MDR strains of N. gonorrhoeae. ... 120
Figure 3.9 Protocol for the in vivo efficacy testing of LpxC inhibitors. ... 122
Figure 3.10 In vivo efficacy of LpxC inhibitors. ... 123
Figure 4.1 Spontaneous mutants with reduced susceptibility to LpxC inhibitors. ... 136
Figure 4.2 Phenotype of the R7 mutant. ... 137
Figure 4.3 Isolation of the DraI DNA fragment containing the mutation ... 139
Figure 4.4 Map of the region deleted in R7. ... 141
Figure 4.5 Both pqiA and pqiB are required for susceptibility to LpxC inhibitors. ... 145
Figure 4.6 Expression of pqiA and pqiB in other mutants. ... 147
LIST OF TABLES
Table 1.1 Proteins involved in type four pilus biogenesis ... 15
Table 2.1 Neisseria gonorrhoeae strains used in this study ... 66
Table 4.1 Genes that were truncated or deleted in R7. ... 140
Table 4.2 Amino acid sequence identity of PqiA and PqiB ... 143
LIST OF ABBREVIATIONS
CDC Centers for Disease Control and Prevention
CEACAM carcinoembryonic antigen cell adhesion molecule
CF Cystic fibrosis
CLSI Clinical and Laboratory Standards Institute
CMP-‐NANA 5’-‐cytidinemonophospho-‐N-‐acetyl neuramic acid
CR3 complement receptor 3
DGI disseminated gonococcal infection
DUS DNA uptake sequence
EDTA ethylenediaminetetraacetic acid
ESC expanded-‐spectrum cephalosporins
GC gonococcus
GGI gonococcal genetic island
GISP Gonococcal Isolate Surveillance Project
GlcNAc N-‐acetylglucosamine
HIV human immunodeficiency virus
HPSG heparin sulfate proteoglycan
IPTG isopropyl β-‐D-‐1-‐thiogalactopyranoside
Kdo 2-‐keto-‐3-‐deoxyoctulosonic acid
LPS lipopolysaccharide
LOS lipooligosaccharide
MIC minimum inhibitory concentration
MurNAc N-‐acetylmuramic acid
PBP penicillin-‐binding protein
PEA phosphoethanolamine
PID pelvic inflammatory disease
PG peptidoglycan
ROS reactive oxygen species
Opa opacity-‐associated proteins
SDS sodium dodecyl sulfate
SEM scanning electron microscopy
STI sexually transmitted infection
ssDNA single-‐stranded DNA
TFP type four pili
T4SS type IV secretion system
UDP-‐GlcNAc uridine diphosphate N-‐acetylglucosamine
WHO World Health Organization
Chapter 1. Introduction
1.1 Introduction
Neisseria gonorrhoeae is a Gram-‐negative diplococcus that causes the sexually transmitted infection (STI) gonorrhea. It is one of the two pathogenic members of the Neisseriaceae family; the other is N. meningitidis, the etiological agent of bacterial meningitis. Gonorrhea is the second-‐most prevalent reportable infectious disease in the United States (Division of STD Prevention 2012). Patients do not develop a protective immune response against gonorrhea, making antibiotic treatment necessary for curing the infection. However, antibiotic resistance in the gonococcus (GC) is a serious public health problem. In the US, gonorrhea is currently treated with the expanded-‐spectrum
cephalosporin ceftriaxone in combination with either azithromycin or doxycycline (Centers for Disease Control and Prevention. 2012). Unfortunately, strains resistant to ceftriaxone have been isolated in the last several years (Unemo, Golparian et al. 2010, Ohnishi, Saika et al. 2011, Unemo, Golparian et al. 2012). This dissertation describes the interconnected mechanisms of antibiotic action and antibiotic resistance in N. gonorrhoeae, as well as the validation of a new potential drug target for the treatment of antibiotic-‐resistant
gonorrhea.
1.2 Public heath relevance
1.2.1 Epidemiology
The Centers for Disease Control and Prevention (CDC) received reports of 321,849 cases of gonorrhea in the United States in 2011, the most recent year for which data are available. This number represents a rate of 104.2 cases per 100,000 people. Alarmingly, the gonorrhea rate in the US increased in both 2010 and 2011 (Division of STD Prevention 2012). The increase occurred in all regions of the US and was most dramatic for the Northeast, but the overall rate remained the highest in the South (Figure 1.1). Because infections are often asymptomatic, especially in females, it is estimated that fewer than half of the gonorrhea cases that occur in the US are reported; thus, the actual burden is likely closer to 800,000 cases per year (Satterwhite, Torrone et al. 2013).
It is more difficult to estimate the world-‐wide burden of gonorrhea. Based on data collected in 2008, the World Health Organization (WHO) estimated that 106 million new gonococcal infections occur each year and that at any given time, 36.4 million people are infected with N. gonorrhoeae (2012). These estimates are considerably higher (11% increase) than the same estimates made for 2005. Although the increase is partially driven by population growth, it represents a 21% increase in the global incidence of gonorrhea.
Figure 1.1 Geographical distribution of gonorrhea infections across the United States
(adapted from the CDC Sexually Transmitted Diseases Surveillance Report (Division of STD Prevention 2012)).
1.2.2 Clinical manifestations
N. gonorrhoeae is an obligate human pathogen. It typically colonizes the genital tracts of men and women but can also infect other mucus membranes, including the anorectal canal, pharynx, and eye. Men typically develop symptoms including urethral discharge and a burning sensation while urinating within 3 to 7 days following exposure (Spence 1983). However, some men have less severe symptoms, and a small proportion of infected men may remain asymptomatic (Handsfield, Lipman et al. 1974, Spence 1983). Interestingly, heterosexual men that are co-‐infected with N. gonorrhoeae and Chlamydia trachomatis and men who have sex with men (MSM) and are infected with both gonorrhea and syphilis are more likely to be asymptomatic than men infected with N. gonorrhoeae alone (Bozicevic, Fenton et al. 2006). Rectal infections are common in MSM, and these infections are often asymptomatic (Miller 2006). Rectal infections can also be more difficult to detect by culturing, and infections are often missed if only the urogenital tract is sampled for nucleic acid amplification testing (van Liere, Hoebe et al. 2013).
Asymptomatic urogenital infections are more common in women, especially in cases where the infection has not yet ascended the genital tract. When symptoms are present, they include discharge and slight irritation (McCormack, Stumacher et al. 1977, Platt, Rice et al. 1983). Anorectal infections are also generally asymptomatic and most often occur secondary to endocervical infections due to the spread of the bacteria across the perineum (McCormack, Stumacher et al. 1977).
asymptomatic, and these women continue to pose a public health threat as carriers of gonorrhea. When symptoms are present, they include abdominal pain, pelvic pain, and fever. If this condition remains untreated, it can result in ectopic pregnancy and even infertility due to damage to the fallopian tubes (Weström, Joesoef et al. 1992, Walker and Sweet 2011).
In both sexes, a small number (0.5 to 3%) of untreated gonorrhea cases progress to disseminated gonococcal infections (DGI), a bacteremia that can result in chronic joint infections, arthritis, or a skin rash (Britigan, Cohen et al. 1985, Bardin 2003). DGI is more common in women than men, likely because asymptomatic mucosal infections are more common in women and because menstruation is a risk factor for DGI (Holmes, Counts et al. 1971, Bleich, Sheffield et al. 2012). Similarly, DGI is more common among MSM than among heterosexual men because anorectal gonorrhea is more likely to be asymptomatic than urethral gonorrhea (Holmes, Counts et al. 1971). Once DGI is diagnosed, it can be treated with antibiotics (Miller 2006).
1.2.3 Gonorrhea and HIV co-‐infections
increased transmission and acquisition have been described. One of the first studies on the relationship between gonorrhea and HIV found that treating the gonococcal infection in co-‐ infected patients reduced viral RNA levels in the semen (Cohen, Hoffman et al. 1997). Women infected with gonorrhea have greater numbers of endocervical CD4+ T-‐cells that
can then be infected upon exposure to HIV (Levine, Pope et al. 1998), and infection with gonorrhea can enhance the transcription of HIV in T-‐cells infected with the virus, leading to increased viral expression and shedding (Chen 2003). Infection with gonorrhea also
increases the susceptibility of immune cells to HIV. Ding et al. found the exposure of CD4+
T-‐cells to N. gonorrhoeae increased HIV infection of these cells in an TLR2-‐dependent manner (Ding, Rapista et al. 2010), and co-‐infection with gonorrhea enhances the infection of dendritic cells by HIV (Zhang, Li et al. 2005). These studies show that an individual with gonorrhea is at an increased risk of contracting gonorrhea, and an individual with both gonorrhea and HIV has an increased capacity to transmit HIV. Thus, treating gonococcal infections is one strategy to decrease the spread of HIV.
1.3 Structural components of the gonococcal cell wall
Like all Gram-‐negative bacteria, N. gonorrhoeae possesses a cell wall consisting of an inner (cytoplasmic) membrane, a thin layer of peptidoglycan, and an outer membrane. The outer membrane forms a permeability barrier and is composed of lipooligosaccharide, phospholipids, integral membrane proteins, and structures that are necessary for genetic transformation, adhesion to host cells, and the import and export of nutrients. These outer membrane components include the type four pili, Opa proteins, and porins.
1.3.1 Lipooligosaccharide
Gram-‐negative bacteria are characterized by the presence of lipopolysaccharide (LPS; also known as endotoxin) in the outer leaflet of the outer membrane. LPS is composed of lipid A, which anchors LPS to the membrane, a polysaccharide core, and O-‐ antigen repeats that extend from the bacterial surface (Raetz and Whitfield 2002). Because N. gonorrhoeae has a shorter polysaccharide core and lacks the O-‐antigen repeats, the endotoxin of GC is called lipooligosaccharide (LOS) (Preston, Mandrell et al. 1996). Bacteria that produce LOS rather than LPS have cell surfaces that are more hydrophobic and are more likely to colonize the respiratory tract, genital tract, and other mucosal surfaces. In contrast, bacteria that produce LPS are more hydrophilic, and this property allows enteric bacteria to be more resistant to intestinal enzymes and bile (Preston, Mandrell et al. 1996).
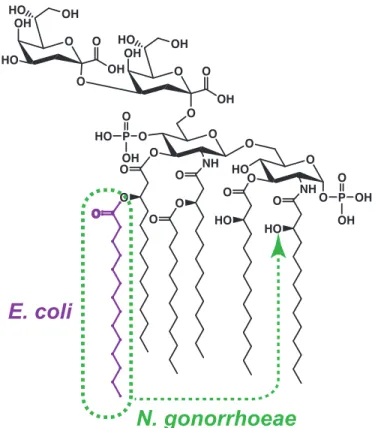
N. gonorrhoeae lipid A consists of two glucosamine molecules, each with 3 acyl chains, bound to two Kdo (2-‐keto-‐3-‐deoxyoctulosonic acid) residues (Figure 1.2). This lipid A moiety is the basic structure of LOS required for cell viability and is the active component of endotoxin. Two heptose molecules are attached to one of the Kdo residues, and 3
variable oligosaccharide side chains can then be attached to the heptose molecules by a series of glycosyltransferases. The oligosaccharides added to the chain include glucose, galactose, GlcNAc, and GalNAc, but their exact composition and order depend on the presence of specific glycosyltransferase enzymes (Preston, Mandrell et al. 1996).
frame-‐shift mutations that turn the genes on or off, leading to subsequent antigenic variations in the sugars added to the LOS core (Danaher, Levin et al. 1995, Yang and
Gotschlich 1996, Banerjee, Wang et al. 1998). Further variability in this region is conferred by the presence of multiple promoter regions in in the lgtABCDE gene cluster. The presence of multiple promoters allows for slight variations in transcription that make it possible for a single cell to express multiple LOS variations simultaneously (Burch, Danaher et al. 1997, Braun and Stein 2004).
In addition to the varying expression of the glycosyltransferases, the structure of LOS can also be modified. The heptose II residue of LOS and the lipid A moiety itself can be decorated with phosphoethanolamine (PEA) (Cox, Wright et al. 2003, O'Connor,
Piekarowicz et al. 2006). The presence of PEA affects the ability of the bacterium to resist host defenses such as serum killing, complement binding, and cationic antimicrobial peptides (Lewis, Choudhury et al. 2009, Balthazar, Gusa et al. 2011, Lewis, Shafer et al. 2012). A N. gonorrhoeae mutant that lacks the PEA transferase (LptA) (Cox, Wright et al. 2003) and is unable to add PEA modifications to lipid A was less fit compared to wild-‐type in both the murine and human male urethra models of gonococcal infection (Hobbs, Anderson et al. 2013).
urethral infection or incubation with urethral epithelial cells in vitro reduces the infectivity of the gonococcus (Schneider, Schmidt et al. 1996, Harvey, Jennings et al. 2001).
Figure 1.2 Structure of Kdo2-‐lipid A. Compared to the structure of E. coli Kdo2-‐lipid A, the
structure of gonococcal Kdo2-‐lipid A differs in the location of one acyl chain.
1.3.2 Type IV pili
Type four pili (TFP) are phase-‐variable surface appendages in many Gram-‐negative bacteria that mediate multiple biological processes, including adhesion (Swanson 1973, Virji and Heckels 1984, Jonsson, Ilver et al. 1994), twitching motility (Swanson 1978, Henrichsen 1983), and DNA transformation (Sparling 1966) (Figure 1.3A). The presence or absence of pili can be observed by examining colony morphology under a dissecting
microscope. Colonies that express pili are small, shiny, and rounded, whereas colonies containing non-‐piliated cells are large and appear dull and flat (Kellogg, Peacock et al. 1963). The pili can be rapidly extended and retracted in an ATP-‐dependent manner. A transposon mutagenesis screen in N. meningitidis identified fifteen proteins that are essential for the biogenesis of TFP (Carbonnelle, Hélaine et al. 2004). Figure 1.3B shows a model of type four pilus biogenesis and the locations of many of these proteins, and Table 1.1 lists pilus biogenesis proteins and their functions.
may stabilize the PilE polymers (Jonsson, Nyberg et al. 1991, Wolfgang, van Putten et al. 2000, Morand, Bille et al. 2004). There is also evidence that PilC serves as an adhesion factor for interactions between the gonococcus and human epithelial cells; however, this is also controversial and may apply only to specific tissue types (Nassif, Beretti et al. 1994, Rudel, Facius et al. 1995, Rudel, Scheurerpflug et al. 1995, Kirchner and Meyer 2005). PilC is also required for the natural competence of the organism (Rudel, Facius et al. 1995).
The polymerization (extension) and depolymerization (retraction) of the filaments requires the activity of the ATPases PilF and PilT, respectively (Freitag, Seifert et al. 1995, Wolfgang, Lauer et al. 1998, Forest, Satyshur et al. 2004). The assembled filaments extrude from the cell through the outer membrane secretin PilQ (Wolfgang, van Putten et al. 2000, Chen and Dubnau 2004). PilQ forms SDS-‐resistant multimers; the formation of these multimers requires the presence of the lipoprotein PilW (Collins, Davidsen et al. 2001, Carbonnelle, Hélaine et al. 2004, Szeto, Dessen et al. 2011). In Pseudomonas aeruginosa, a recently defined association of PilM, PilN, PilO, and PilP referred to as the “alignment complex” (Tammam, Sampaleanu et al. 2013) was shown to bridge the periplasm to link the inner membrane components of TFP assembly with the PilQ secretin. These proteins (PilMNOP) are all required for piliation in N. gonorrhoeae, and similar associations are suggested by the work presented in Chapter 2.
Hamrick, Dempsey et al. 2001). These copies, or minicassettes, lack the N-‐terminal conserved region and contain increasing numbers of amino acid substitutions within the hypervariable C-‐terminal domain. Each minicassette is flanked by regions of repeats that match sequences found upstream and within pilE (Haas and Meyer 1986, Howell-‐Adams and Seifert 2000), and these sequences are important for recombination. Gene conversion occurs at a frequency of approximately 10-‐3 in vitro (Serkin and Seifert 1998), and variants
appear rapidly following infection (Hamrick, Dempsey et al. 2001). It is possible that
selective pressure during infection may increase the frequency of gene conversion to above that measured in vitro. Some recombination events can result in a deletion in the
expression locus, and these deletions result in the phase variation from an “on” (piliated) to an “off” (nonpiliated) colony morphology (Segal, Billyard et al. 1985).
A. B.
Figure 1.3 Type IV pili of N. gonorrhoeae. A. Scanning electron micrograph of piliated FA19 cells (magnification: 100,000×). B. Model showing the location of components involved in the biogenesis of TFP. The pilus is primarily composed of PilE subunits and the PilC adhesin and extrudes through the PilQ secretin in the outer membrane. The protein PilW is
required for the stability of the secretin. The pilin subunits are processed by the pre-‐pilin peptidase PilD and polymerized through the activity of PilF. Pilus retraction is mediated by the PilT ATPase. The proteins PilM, PilN, PilO, and PilP are required for piliation, as
discussed in Chapter 2 of this dissertation (Wolfgang, van Putten et al. 2000, Tammam, Sampaleanu et al. 2011).
Table 1.1 Proteins involved in type four pilus biogenesis
Protein( Func,on(
PilE% Pilin%
PilF% ATPase,%filament%polymeriza6on%(extension)%
PilT% ATPase,%filament%depolymeriza6on%(retrac6on)%
PilD% Pre=pilin%pep6dase%%
PilG% Pilus%assembly%protein%
PilX% Minor%pilin%
PilW% Stabiliza6on%of%PilQ%
PilM% Required%for%TFP%biogenesis%%
PilN% Required%for%TFP%biogenesis%%
PilO% Required%for%TFP%biogenesis%%
PilP% Required%for%TFP%biogenesis%%
PilQ% TFP%secre6n%
1.3.3 Opacity-‐associated proteins
The opacity-‐associated proteins (Opa) are antigenic-‐ and phase-‐variable outer-‐ membrane adhesins that are involved in the association of pathogenic Neisseria with host cells and contribute to the ability of the bacteria to evade the immune system. These proteins were named for the opaque appearance of Opa+ colonies on agar viewed under a
dissecting microscope with a substage light supply (Swanson 1978). Gonococcal cells recovered from infected individuals are Opa+, even when the infecting inoculum contained
only Opa-‐ cells (James and Swanson 1978, Jerse, Cohen et al. 1994). N. gonorrhoeae strains
typically encode 11 Opa proteins (Bhat, Gibbs et al. 1991), and a single cell may express any number of these proteins simultaneously. The Opa genes are constitutively transcribed, and phase variation occurs at the translational level. Phase variation is due to the presence of pentameric repeats (CTCTT) in the coding sequence (Stern, Brown et al. 1986); the number of these repeats varies due to slip-‐strand mispairing that can move the coding regions of the Opa genes in or out of frame (Murphy, Connell et al. 1989).
The structure of the Opa proteins has not been solved, but a two-‐dimensional model has been predicted based on protein sequences and epitope binding of monoclonal
The Opa proteins can be divided into two categories based on the host factors to which they bind. A small number of Opa proteins bind to heparin sulfate proteoglycans (HSPGs) (Chen, Belland et al. 1995, van Putten, Duensing et al. 1998). However, the majority of Opa proteins bind to members of the carcinoembryonic antigen cell adhesion molecule (CEACAM) family (Wang, Gray-‐Owen et al. 1998, Virji, Evans et al. 1999). These receptors are found on a variety of cell types and can influence the immune response to gonococcal infection (Liu, Feinen et al. 2011).
1.3.4 Peptidoglycan
Peptidoglycan (PG) is a polymer of carbohydrate and amino acids that is sandwiched between the inner and outer membranes of Gram-‐negative bacteria. This essential macromolecule confers structure and shape to the cell and protects it from osmotic stress. PG is composed of alternating N-‐acetylglucosamine (GlcNAc) and N-‐ acetylmuramic acid (MurNAc) sugars linked with a β-‐1,4 glycosidic bond. In N.
PG is constantly remodeled as bacterial cells grow and divide, and peptidoglycan fragments are released from the cell. This remodeling process requires the activity of endopeptidases and amidases that hydrolyze the peptide chains and lytic transglycosylases (LTs) that hydrolyze the glycan backbone (Johnson, Fisher et al. 2013). During remodeling, the LTs cleave the glycosidic bond between the GlcNAc and MurNAc residues of the glycan strand by catalyzing the formation of 1,6-‐anhydro bonds within the MurNAc residues (Rosenthal 1979, Chan, Hackett et al. 2012). Peptidoglycan fragments released during remodeling may contribute to the pathogenesis of gonorrhea. For example, when gonococcal PG monomers are applied to human fallopian tubes in organ culture, the ciliated cells are damaged and sloughed off of the mucosa, similar to what has been observed in patients suffering from PID (Melly, McGee et al. 1984).
1.3.5 Porin
Porins are abundant proteins that form water-‐filled channels through the outer membranes of Gram-‐negative bacteria to allow for the diffusion ions across the membrane (Young, Blake et al. 1983, Nikaido 1994). In addition to their general channel function, neisserial porins also interact with host cells and are involved in evading the immune system. Porins can insert into the host cell membrane during infection and arrest
membrane-‐spanning channel, the fully assembled porin contains three channels (Judd 1989, Nikaido 1994, Derrick, Urwin et al. 1999).
The gonococcal porin is encoded by the porB gene, and a given strain possesses only one of two porB alleles that encode either PorB1A or PorB1B (Knapp, Tam et al. 1984,
Carbonetti, Simnad et al. 1988). The exact sequences of the two alleles can differ between strains, but depending on the strains compared, the alleles demonstrate 65 to 80%
homology (Judd 1989). The PorB1A proteins are slightly smaller than PorBIB proteins, with
apparent molecular masses of approximately 35 kDa compared to ~37 kDa for PorB1B
proteins (Gotschlich, Seiff et al. 1987). Some of the differences between the two PorB proteins and within each subclass have been exploited in various serotyping methods (Knapp, Tam et al. 1984, Sarafian and Knapp 1989).
Strains carrying PorB1A are most often associated with disseminated gonococcal
infection (Cannon, Buchanan et al. 1983). These strains are more invasive than PorB1B
strains (van Putten, Duensing et al. 1998) and are also more resistant to killing by human serum (Schoolnik, Buchanan et al. 1976, Carbonetti, Simnad et al. 1990). Two mechanisms by which PorB1A interferes with complement binding have been described. It can bind C4b
binding-‐protein, a component of the classical complement pathway, through interactions with the N-‐terminal surface-‐exposed loop (Ram, Cullinane et al. 2001). It can also resist complement attack by binding factor H, a regulatory protein in the alternative complement pathway, with its fifth surface-‐exposed loop (Ram, McQuillen et al. 1998). Together, these properties of PorB1A may explain its increased ability to disseminate from the initial site of
In contrast to the invasiveness of strains possessing PorB1A, strains carrying PorB1B
more often cause localized infections. This allele is also associated with increased antibiotic resistance. PorB1B strains demonstrate a lower baseline susceptibility to penicillin and
erythromycin compared to strains with PorB1A (Woodford, Bindayna et al. 1989,
Carbonetti, Simnad et al. 1990). However, PorB1B is able to develop mutations that can lead
to resistance to penicillin, tetracycline, and other antimicrobials (Olesky, Hobbs et al. 2002).
1.4 Infection
1.4.1 Adhesion and invasion
Of the structural components described in the previous section, TFP, LOS, Opa, and porin are involved in mediating the attachment to and invasion of epithelial cells by
tissues, including primary male urethral epithelial cells, the pilus has been shown to interact with the α1β1 and α2β1 integrins (Edwards and Apicella 2005).
Following the initial TFP-‐mediated contact with host cells, Opa proteins are
required for tight binding to host cells. Gonococci express multiple Opa proteins, and these different Opa proteins bind to one of two types of mammalian receptors: carcinoembryonic antigen cell adhesion molecules (CEACAMs) or the heparin sulfate proteoglycans (HSPGs). Opa variants that bind to HSPGs are less common than those variants that bind CEACAMs. In some cell lines, Opa binding to HPSGs is sufficient for invasion (Chen, Belland et al. 1995); however, the majority of Opa proteins bind to CEACAMs (Dehio, Gray-‐Owen et al. 1998, Gray-‐Owen 2003). The CEACAM family receptors are differentially expressed by human tissues (Thompson, Grunert et al. 1991). Of the at least 12 CEACAM receptors, CEACAM1, CEACAM3, CEACAM5, and CEACAM6 are known to serve as receptors for the Opa proteins. Epithelial cells express CEACAM1, CEACAM5, and CEACAM6. Neutrophils express CEACAM3 and smaller amounts of CEACAM1 and CEACAM6. CEACAM6 is also found on monocytes, and Opa proteins also interact with CEACAM1 on T and B
lymphocytes (Wang, Gray-‐Owen et al. 1998, Sadarangani, Pollard et al. 2011).
interactions between Opa proteins and the CEACAM1 receptor on these cells may result in immune suppression. The binding of Opa proteins to CEACAM1 can suppress the activation and proliferation of CD4+ T cells (Boulton and Gray-‐Owen 2002). In B cells, Opa binding can
inhibit antibody production and induce cell death (Pantelic, Kim et al. 2005).
Using microscopy to observe interactions between the gonococcus and host cells, a number of labs have described the gross morphological changes in host epithelial cells during infection. Bacterial cells initially attach in clusters of 10 to 100 diplococci on the surface of the cells (Griffiss, Lammel et al. 1999), prompting the extension of the host cell microvilli towards the bacteria (Ward and Watt 1972, Shaw and Falkow 1988) and the formation of cortical plaques (Merz, Enns et al. 1999). Later in infection (6-‐18 h), the bacteria spread across the epithelial surface as individual diplococci, become tightly associated with the host membrane, and are subsequently engulfed by the host cells (McGee, Johnson et al. 1981, Shaw and Falkow 1988, Griffiss, Lammel et al. 1999).
Following internalization, gonococci are observed in both the cytosol and within vacuoles. At approximately 40 hours post-‐infection, the bacteria egress from the epithelial cells and colonize the subepithelial tissues (McGee, Johnson et al. 1981, Apicella, Ketterer et al. 1996, Mosleh, Boxberger et al. 1997).
1.4.2 Bacterial defenses against the host immune response
gonorrhoeae possesses multiple mechanisms to resist killing by the innate immune system and to suppress the adaptive immune response.
One way by which the innate immune system kills bacteria is through activation of the complement system. Activation of complement can lead to bacterial lysis, opsonization, and phagocytosis. However, several gonococcal surface structures are able to bind
components of the complement system, preventing its activation and evading lysis and phagocytosis. Serum resistance in gonococci can be variable or stable. Variable serum resistance is mediated by the binding of factor H to sialylated LOS, and serum resistance can be restored to sensitive strains by adding 5’-‐cytidinemonophospho-‐N-‐acetyl neuramic acid (CMP-‐NANA), a sialyl residue donor, to the media (Parsons, Andrade et al. 1989, Ram, Sharma et al. 1998). Factor H is a cofactor for the cleavage of C3b to the inactive iC3b. When GC are bound to factor H, nearly all of the C3b that they are associated with becomes converted to iC3b, thus preventing complement-‐mediated killing. Stable serum resistance is mediated by the binding of complement components to porin. Loop 5 of PorB1A binds factor H, and loop 1 binds to C4b-‐binding protein (Ram, McQuillen et al. 1998, Ram,
Cullinane et al. 2001). These interactions inhibit all three complement activation pathways. Following infection with N. gonorrhoeae, neutrophils are recruited to the site of the infection. Neutrophils phagocytose bacteria and typically kill them via the production of reactive oxygen species (ROS). However, some gonococci are able to resist killing by neutrophils and are even able to replicate within the phagocytes (Simons, Nauseef et al. 2006, Criss, Katz et al. 2009, Johnson and Criss 2011). GC possess multiple mechanisms to resist oxidative stress. They produce superoxide dismutase (SOD), an enzyme that
al. 2006); the hydrogen peroxide can then be neutralized by conversion to water and oxygen by catalase (KatA) and cytochrome c peroxidase (CCP) (Archibald and Duong 1986, Turner, Reid et al. 2003, Stohl, Criss et al. 2005). Superoxide and hydrogen peroxide can also be quenched by the manganese ion (Mn2+), and gonococci import Mn2+ into the
bacterial cytoplasm via the MntABC transporter (Tseng, Srikhanta et al. 2001).
The ability of N. gonorrhoeae to vary its surface structures allows the gonococcus to evade antibody-‐mediated immune responses. There is a growing body of evidence that N.
gonorrhoeae actively suppresses cell-‐mediated immunity as well. A study conducted using
the mouse model of gonococcal infection showed that infection with N. gonorrhoeae increased the production of TGF-‐β, and this cytokine subsequently enhanced the Th17 response while inhibiting the differentiation and proliferation of Th1 and Th2 cells. These T-‐helper cells are involved in activating both CD8+ cytotoxic T-‐cells and antibody-‐
producing plasma cells (Feinen, Jerse et al. 2010, Liu, Feinen et al. 2011, Liu, Islam et al. 2012). As described above, interactions between neisserial Opa proteins and CEACAMs expressed on activated CD4+ T cells also suppress the T-‐helper cell response (Boulton and Gray-‐Owen 2002), and Opa-‐CEACAM interactions on B cells can suppress the antibody response (Pantelic, Kim et al. 2005).
1.4.3 Murine model of gonococcal infection
colonized, but they then cleared the infection upon entering the post-‐ovulatory stages (Streeter and Corbeil 1981, Johnson, Tuffrey et al. 1989). To circumvent this issue, the current model of murine gonococcal infection, primarily developed by Ann Jerse’s group, uses estradiol treatment to make the genital tract of female BALB/c mice amenable to colonization (Taylor-‐Robinson, Furr et al. 1990, Jerse 1999). In this model, mice are injected with water-‐soluble estradiol on days -‐2, 0, and 2 of the infection and inoculated with N. gonorrhoeae on day 0. GC can be recovered on vaginal swabs for approximately 12 days post-‐infection until the infection is naturally cleared (Jerse 2011).
The development of the mouse model of gonorrhea made it possible to study the infection in vivo without using the human male urethral challenge mode. However, it does have some limitations. Many of the host proteins that the bacterium interacts with in its human host, including the pilus receptor CD46 and Opa-‐interacting proteins CEACAM1, 3, 5, and 6, are either not expressed by mice or have species-‐specific amino acid changes that prevent recognition by the bacterium. The murine complement proteins factor H and C4bp-‐ binding protein also fail to bind to GC (Jerse 2011). Nevertheless, the murine model of gonorrhea has been used to study the immune response to GC and to evaluate the fitness cost or advantage of different strains of GC (Wu and Jerse 2006, Song, Condron et al. 2008, Warner, Shafer et al. 2008, Feinen, Jerse et al. 2010, Hobbs, Anderson et al. 2013).
In chapter 3 of this dissertation, we describe the use of the murine model of
gonorrhea to evaluate the in vivo activity of a novel class of therapeutics against gonorrhea.
1.5 Genetic transformation
1.5.1 Natural competence
N. gonorrhoeae is one of the few naturally competent bacterial species (Johnsborg, Eldholm et al. 2007) (other naturally competent bacteria include Streptococcus pneumoniae and Haemophilus influenzae). The gonococcus is capable of acquiring DNA from its
environment and recombining it into its chromosome, provided that the exogenous DNA contains a Neisseria-‐specific 10-‐bp uptake sequence and has regions of homology
(generally >80%) with existing chromosomal DNA (Goodman and Scocca 1988, Elkins, Thomas et al. 1991). The ability of the gonococcus to take up and integrate exogenous DNA into its chromosome provides a means for the bacteria to undergo genetic variation and to become resistant to antibiotics and has aided the development of tools for the genetic manipulation of the organism. In contrast to the majority of the other naturally competent bacteria, N. gonorrhoeae remains competent for transformation throughout all phases of growth (Sparling 1966, Biswas, Sox et al. 1977).
Although it occurs at reduced efficiency, some strains can also be transformed with DNA that lacks the DUS; however, the mechanism behind this non-‐specific uptake remains unclear (Duffin and Seifert 2010).
1.5.2 Sources of DNA
Because the gonococcus is an obligate human pathogen, the sources of DNA for transformation are other co-‐infecting strains of gonorrhea, the closely related pathogenic species, N. meningitidis, and the commensal species N. lactamica, N. cinerea, and N.
flavescens (Hamilton and Dillard 2006). Notably, transformation with DNA from
commensal Neisseria species contributes to the formation of an altered PBP allele that confers increased resistance to penicillin in N. gonorrhoeae (Spratt, Bowler et al. 1992) and mosaic PBP alleles that confer resistance to the expanded-‐spectrum cephalosporins,
ceftriaxone and cefixime. DNA is released from these donor bacteria via autolysis or type IV secretion (Hamilton and Dillard 2006).
Seifert 2001). This is in contrast with DNA donation that occurs due to the lysis and death of donor cells, which primarily occurs during the stationary phase of in vitro growth.
Autolysis occurs when cells are exposed to suboptimal growth conditions, including nutrient limitation (Hebeler and Young 1975). This process likely relies on the activity of amidases and LTs to breakdown the cell wall (Hebeler and Young 1975, Chan, Hackett et al. 2012). The DNA released due to the autolysis of the donor cells has been shown to be a viable source of DNA for transformation and can be taken up by recipient cells (Norlander, Davies et al. 1979).
1.5.3 DNA binding and uptake
The mechanism that mediates the initial binding of double-‐stranded DNA to the cell surface is unknown, but it occurs independently of the DUS and may involve Opa proteins and minor pilus proteins (Hill 2000, Aas, Løvold et al. 2002, Aas, Wolfgang et al. 2002). Once the DNA has made non-‐specific contact with the cell, the DUS binds to an unknown protein on the cell surface. The factors required for uptake of the DNA into the cell have been better characterized. Studies in the 1970s observed that piliated cells transformed much more readily than non-‐piliated gonococci (Biswas, Sox et al. 1977), and many of the components of the TFP have been shown to be essential for competence (Rudel, Facius et al. 1995, Wolfgang, Lauer et al. 1998, Zhao, Tobiason et al. 2005).