R E S E A R C H
Open Access
Threshold dynamics of a delayed
predator–prey model with impulse via the
basic reproduction number
Xiangsen Liu
1*and Binxiang Dai
2*Correspondence:
1Department of Mathematics,
North University of China, Taiyuan, P.R. China
Full list of author information is available at the end of the article
Abstract
In this paper, we study a delayed predator–prey model with impulse and, in particular, the existence of the predator-free periodic solution. We employ the approach and techniques coming from epidemiology and calculate the basic reproduction number for the predator. Using the basic reproduction number, we consider the global attraction of the predator-free periodic solution and uniform persistence of the predator. Our results improve the results by Li and Liu (Adv. Differ. Equ. 2016:42,2016), where they left the open problem of finding a threshold value that determines the eradication and uniform persistence of the predator. Furthermore, we give some numerical simulations to illustrate our results.
Keywords: Predator–prey model; Delay; Impulse; Basic reproduction number
1 Introduction
In the natural world, many species usually pass through a number of stages during their life cycle. So it is practical to introduce time delay into models of theoretical ecology. In par-ticular, it is often important to take into account the processes of gestation and maturation to make an abstract model more biologically realistic [2–4]. The characteristic property of population models with time delay is their oscillatory behavior: for a sufficiently large mat-uration period, an initially stable equilibrium becomes unstable, and the system exhibits sustained oscillations [2,5]. Additionally, impulsive differential equations have been ex-tensively used as models in biology, physics, chemistry, engineering, and other sciences with particular emphasis on population dynamics [6–9]. In [9] the authors discussed an impulsive predator–prey system with stage structure and generalized functional response. Sufficient conditions are established for the existence of a predator-free positive periodic solution and the permanence of the system. Numerical simulation shows that impulses and functional response affect the dynamics of the system.
authors considered the following delayed model with impulse:
wherex(t) andy(t) denote the population densities of prey and predator at timet, respec-tively,r,K,α,β,k,dare positive,ris the intrinsic rate of increase of the prey,Kis the carrying capacity of the prey,αis the predation coefficient of the predator, which reflects the size of the predator’s ability, β is the predation regulation factor (saturation factor) of the predator, dis the death rate of the predator,k(0 <k< 1) is the rate of convers-ing prey into predator,τ > 0 denotes a time delay due to the gestation of the predator,
x(t) =x(t+) –x(t),y(t) =y(t+) –y(t),T is the impulsive period,n∈N
+={1, 2, . . .}, and
p> 0 is the proportionality constant, which represents the rate of mortality due to the applied pesticide. The initial conditions for system (1.1) are
φ1(s),φ2(s)
∈C [–τ, 0],R2+, φi(0) > 0 (i= 1, 2). (1.2)
In [1], sufficient conditions for the global attraction of a predator-free periodic solu-tion are obtained by the theory of impulsive differential equasolu-tions, that is,T <T1∗. The conditions for the permanence of the system are investigated, that is,T>T2∗. Note that
T1∗<T2∗always holds. It is obvious that ifT∈(T1∗,T2∗), then we cannot determine whether the predator can persist or not. In the present paper, we give a thorough global dynam-ics of (1.1), which completely solves the question left in [1]. To do this, we employ the approach coming from epidemiology [13]. As far as we know, there are no papers em-ploying this approach in ecology. Throughout the present paper, roughly speaking, the basic reproduction numberR0 may be thought as the number of predators one preda-tor gives rise during its life, when introduced in a prey population [14]. A similar thresh-old value for the coexistence of a predator–prey system has previously been formulated and explained by Pielou [15], among others but, to the best of our knowledge, has not been termed a “basic reproduction number.” In ecology, many authors have investigated the autonomous predator–prey systems using the basic reproduction number [16,17]. For example, in [16] the authors considered a stage-structured predator–prey model with nonlinear predation rate. They discussed the stability of the system using the basic re-production number of the predator population. In contrast, there have been few papers discussing the nonautonomous, delayed, or impulsive predator–prey systems using the basic reproduction number (except for [18]). In [18] the authors considered an ecoepi-demiological model with Holling type-III functional response and time delay. They used the ecological and disease basic reproduction numbers to determine the persistence of the system. In this paper, using the basic reproduction number of the predator population and approach in [13], we wish to find a threshold value to determine whether the predator can exist or not.
we employ the approach coming from epidemiology and calculate the basic reproduction number for the predator. In Sect.4, using the basic reproduction number, we consider the global attraction of the predator-free periodic solution and persistence of the predator in (1.1). In Sect.5, we give some numerical simulations to illustrate our results. Finally, we give some concluding remarks.
2 The existence of a predator-free periodic solution and boundedness of system (1.1)
In this section, we investigate the existence of a predator-free periodic solution of system (1.1). In this case, the predator population is entirely absent from the population perma-nently, that is,y(t) = 0,t≥0. System (1.1) yields
⎧ ⎨ ⎩
x(t) =rx(t)(1 –xK(t)), t=nT,
x(t) = –px(t), t=nT.
(2.1)
For system (2.1), we have the following result.
Lemma 2.1([1]) If(1 –p)erT> 1,then system(2.1)has the unique positive periodic solution
x∗(t) = x
∗
0
(1 –x∗0/K)e–r(t–nT)+x∗ 0/K
, t∈ nT, (n+ 1)T, (2.2)
which is globally asymptotically stable,where
x∗0=K((1 –p)e
rT– 1)
erT– 1 . (2.3)
According to Lemma2.1, we obtain the following result.
Theorem 2.1 If (1 –p)erT > 1,then system (1.1)has a predator-free periodic solution
(x∗(t), 0),where x∗(t)is shown in(2.2).
Next, we will show that all solutions of (1.1) are uniformly upper bounded.
Theorem 2.2 If(1 –p)erT> 1,then all solutions of(1.1)are uniformly upper bounded. Proof From the first and third equations of (1.1) we have
⎧ ⎨ ⎩
x(t)≤rx(t)(1 –xK(t)), t=nT,
x(t) = –px(t), t=nT.
Consider the following impulsive comparison system:
⎧ ⎨ ⎩
z1(t) =rz1(t)(1 –z1K(t)), t=nT,
z1(t) = –pz1(t), t=nT.
(2.4)
for given0> 0, there existsT1> 0 such that, fort>T1,
x(t)≤z1(t)≤x∗(t) +0≤
x∗0
(1 –x∗0/K)e–rT+x∗
0/K
+0M1. (2.5)
LetV(t) =kx(t–τ) +y(t). Then, fort>T1+τ, we have
V(t) =krx(t–τ)
1 –x(t–τ)
K
–dy(t)
≤krx(t–τ) –dy(t)
= (kr+kd)x(t–τ) –d kx(t–τ) +y(t)
≤k(r+d)M1–dV(t)
and
V (nT+τ)+=kxnT++y (nT+τ)+
=k(1 –p)x(nT) +y(nT+τ)
≤kx(nT) +y(nT+τ)
=V(nT+τ).
Consider the following impulsive comparison system:
⎧ ⎨ ⎩
V1(t) =k(r+d)M1–dV1(t), t=nT+τ,
V1(t) = 0, t=nT+τ.
It is clear that
lim
t→+∞V1(t) =
k(r+d)M1
d .
By the comparison principle, for0> 0, there existsT2>T1+τ such that, fort>T2, we get
V(t)≤V1(t)≤
k(r+d)M1
d +0M2. (2.6)
The proof is completed.
Thus the dynamics of system (1.1) can be analyzed in the following bounded feasible region:
Γ = x(t),y(t)|0≤x(t)≤M1, 0≤y(t)≤M2
,
3 The basic reproduction number of the predator
In this section, we introduce the basic reproduction numberR0of the predator for system (1.1) using the next generation operators approach in [19]. We linearize system (1.1) at (x∗(t), 0) and get
y(t) = kαx
∗2(t–τ)
1 +βx∗2(t–τ)y(t–τ) –dy(t)
=g(t)y(t–τ) –dy(t), (3.1)
where
g(t) = kαx
∗2(t–τ)
1 +βx∗2(t–τ). (3.2)
From (3.1) we obtain
y(t) =y(0)e–dt+e–dt t
0
g(s)y(s–τ)edsds.
Lety1(t) =g(t)y(t–τ). Then
y1(t) =y(0)g(t)e–d(t–τ)+e–d(t–τ)g(t)
t–τ
0
y1(s)edsds.
Making the change of variables=t–δ, we get
y1(t) =y(0)g(t)e–d(t–τ)+g(t)e–d(t–τ)
t
τ
y1(t–δ)ed(t–δ)dδ
=y(0)g(t)e–d(t–τ)+g(t)
t
τ
y1(t–δ)e–d(δ–τ)dδ
=y10(t) +
t
τ
¯
K(t,δ)y1(t–δ) dδ,
where
y10(t) =y(0)g(t)e–d(t–τ)
and
¯
K(t,δ) =
⎧ ⎨ ⎩
0, δ<τ,
g(t)e–d(δ–τ), δ≥τ.
LetCTbe the ordered Banach space of all continuousT-periodic functions fromRto Requipped with the maximum norm · and the positive coneC+
T={ψ ∈CT|ψ(t)≥
0,t∈R}. Then we can define the linear operatorL:CT→CTby
L:v(t)→
+∞
0
¯
Following [19], the basic reproduction number of the predator is defined asR0r(L), the spectral radius ofL.
We see that the basic reproduction numberR0=r(L) can be obtained by solving the eigenvalue problem
Differentiating Eq. (3.3) and then integrating by parts, we have
R0u(t) =g(t)
This equation can be rewritten as
u(t)
Obviously, there is no explicit formula forR0whenτ= 0.
Remark3.1 Whenτ= 0, system (1.1) reduces to a periodic system of ordinary differential equations. The corresponding basic reproduction numberR0becomes
R0=
T
0 g(t) dt
dT .
4 The global dynamics of system (1.1)
In this section, we study the global dynamics of system (1.1) in terms of its basic repro-duction numberR0. To this end, we first introduce some lemmas for our main results.
For anyψ∈C([–τ, 0],R), letP(t)ψ=ut(ψ) be the unique solution of (3.1) satisfying
Consider the following linear equation with time delay:
wherea(t)∈CT, andb(t) is a piecewise left-continuousT-periodic function with discon-tinuous points of first kind atnTsuch thatb(t) > 0,t≥0. By applying the method of steps it is easy to verify that, for anyφ∈C+=C([–τ, 0],R+), system (4.1) has a unique contin-uous solutionu(t,φ) on [–τ, +∞) withu0=φ. LetPbe the Poincaré map associated with (4.1) onC+, that is,P(φ) =uT(φ). Then we have the following result.
Lemma 4.2 [13].Letμ=lnr(TP).Then there exists a positive T -periodic function v(t)such that eμtv(t)is a solution of (4.1).
For the predator-free periodic solution (x∗(t), 0) of (1.1), we have the following result.
Theorem 4.1 Assume that(1 –p)erT> 1.If R
0< 1,then the predator-free periodic solution (x∗(t), 0)of (1.1)is globally attracting,where R0is defined in(3.5).
Proof LetPbe the Poincaré map of the following perturbed linear equation:
y2(t) = kα(x
∗(t–τ) +)2
1 +β(x∗(t–τ) +)2y2(t–τ) –dy2(t). (4.2)
By Lemma4.1we see thatR0< 1 if and only ifr(P) < 1, wherePis the Poincaré map of (3.1). Sincelim→0r(P) =r(P) < 1, we may fix a small enough> 0 such thatr(P) < 1.
By Lemma4.2there is a positiveT-periodic functionv(t) such thateμtv(t) is a positive
solution of (4.2), whereμ=lnrT(P)< 0.
According to the proof of Theorem2.2, for given> 0, there existsT3> 0 such that, for
t>T3,
x(t)≤x∗(t) +, (4.3)
wherex∗(t) is defined in (2.2).
By the second equation of (1.1), fort>T3+τ, we get
y(t)≤ kα(x
∗(t–τ) +)2
1 +β(x∗(t–τ) +)2y(t–τ) –dy(t).
Choose a sufficiently large A> 0 such that y(t)≤Aeμtv
(t), t∈[T3,T3 +τ], where
Aeμtv
(t) is a positive solution of (4.2). By the comparison principle we have y(t)≤Aeμtv
(t), t≥T3+τ.
Sinceμ< 0,limt→+∞y(t) = 0.
Then for a sufficiently small1> 0 (1<αMr1, whereM1is shown in (2.5)), there exists
T4> 0 such that, fort>T4, we obtain
0 <y(t) <1.
From the first and third equations of (1.1), fort>max{T1,T4}, we have
⎧ ⎨ ⎩
x(t)≥x(t)(r–αM11–Krx(t)), t=nT,
Consider the following impulsive comparison system:
⎧ ⎨ ⎩
z2(t) =z2(t)(r–αM11–Krz2(t)), t=nT,
z2(t) = –pz2(t), t=nT.
(4.4)
Since1< αMr1, it is obvious thatr–αM11> 0. According to Lemma2.1, we find that
system (4.4) has a globally asymptotically stable periodic solution
z2∗(t) = z
∗
2(0+)
(1 –rz∗2(0+)/(Kr1))e–r1(t–nT)+rz∗2(0+)/(Kr1)
, nT<t≤(n+ 1)T,
with
z2∗(0+) =Kr1((1 –p)e
r1T– 1)
r(er1T– 1) , r1=r–αM11.
By the comparison principle, forx(0+)≥z2(0+) andt>max{T1,T4}, we havex(t)≥z2(t), andz2(t) –z∗2(t)→0 ast→+∞. Meanwhile,z∗2(t) –x∗(t)→0 as1→0. Based on this analysis and (4.3), we see thatx(t) –x∗(t)→0 ast→+∞. Therefore the predator-free periodic solution (x∗(t), 0) of (1.1) is globally attracting. The proof is completed.
Theorem 4.2 Let(1 –p)erT > 1.If R
0> 1,then there exists q> 0such that every positive
solution(x(t),y(t))of (1.1)satisfies y(t)≥q for t large enough.
Proof LetMηbe the Poincaré map of the perturbed equation
¯
y(t) = kα(x
∗(t–τ) –η)2
1 +β(x∗(t–τ) –η)2y¯(t–τ) –dy¯(t). (4.5)
Sincelimη→0r(Mη) =r(P) > 1, we can fix a small positive numberηsuch thatr(Mη) > 1
andη<inft≥0x∗(t).
By Lemma4.2there is a positiveT-periodic functionvη(t) such thateμηtvη(t) is a positive
solution of (4.5), whereμη=
lnr(Mη)
T > 0.
Consider the impulsive comparison system
⎧ ⎨ ⎩
¯
x(t) =x¯(t)(r–αM1¯–Krx¯(t)), t=nT,
x¯(t) = –px¯(t), t=nT. (4.6)
where 0 <¯<αMr
1, andM1is shown in (2.5).
Using Lemma2.1, we see that system (4.6) admits a positive periodic solutionx∗¯(t), which is globally asymptotically stable, andlim¯→0x∗¯(t) =x∗(t). Hence we can fix a small
number¯∈(0,αMr
1) such that, fort≥0,
x∗¯(t) >x∗(t) – η
2. (4.7)
y(t) <γ fort>t0. It follows from the first equation of (1.1) that, fort>t0,
x(t)≥rx(t)
1 –x(t)
K
–αM1γx(t)
≥x(t)
r–αM1¯–
rx(t)
K
.
Therefore we have
⎧ ⎨ ⎩
x(t)≥x(t)(r–αM1¯–rxK(t)), t=nT,
x(t) = –px(t), t=nT.
By system (4.6) and the comparison principle there existst1>t0such that, fort>t1and
x(0+)≥ ¯x(0+),
x(t)≥ ¯x(t) >x∗¯(t) – η
2. (4.8)
Using (4.7) and (4.8), fort>t1, we get
x(t) >x∗(t) –η. (4.9)
From (4.9) and the second equation of (1.1), fort>t2t1+τ, we have
y(t)≥ kα(x
∗(t–τ) –η)2
1 +β(x∗(t–τ) –η)2y(t–τ) –dy(t). (4.10)
Choose a numberm1> 0 such that
y(t)≥m1eμηtvη(t), t∈[t1,t2], (4.11)
and
m1eμηtvη(t) <γ, t∈[t1,t2], (4.12)
whereeμηtv
η(t) is a positive solution of (4.5) andμη> 0. Using (4.10), (4.11), (4.12), and the
comparison theorem, there must bet3>t2such thatγ<y(t3) <¯, which is a contradiction. From the above claim we discuss the following two possibilities:
(H1) y(t)≥γ for all larget.
(H2) y(t)oscillates aboutγ for all larget.
Letq=min{γ
2,q2}, where
q2=
q1mint∈[0,T]vη(t) eμητmax
t∈[0,T]vη(t)
withq1=γe–dτ.
We hope to show thaty(t)≥qfortlarge enough. Obviously, we only need to consider case (H2). Lettand¯tsatisfy
wheretis sufficiently large such that, fort∈[t,¯t],
x(t) >x∗(t) –η. (4.14)
Note that the function y(t) for t≥0 is uniformly continuous since its derivative is bounded for allt≥0. Hence there existsT(0 <T<τ is independent of the choice of
t) such thaty(t) >γ2 fort∈[t,t+T]. Let us consider the following three cases: Case (B1)t¯–t≤T. Theny(t) >γ2 for allt∈[t,t¯].
Case (B2)T<¯t–t≤τ.
By the second equation of (1.1) we have
y(t)≥–dy(t).
Using the comparison principle, fort∈[t,¯t], we get
y(t)≥y(t)e–d(t–t)
≥γe–dτq
1.
Therefore, in this case,y(t)≥q1fort∈[t,¯t]. Case (B3)t¯–t>τ.
Similarly to the discussion of case (B2), we obtainy(t)≥q1fort∈[t,t+τ]. Ify(t)≥q1 fort∈[t+τ,¯t], then our aim is reached. Suppose not. Then there existsT¯1> 0 such that
y(t)≥q1, t∈[t,t+τ+T¯1],
y(t+τ+T¯1) =q1, y(t) <q1, 0 <t– (t+τ+T¯1)1.
By (4.14) and the second equation of system (1.1), fort∈[t,¯t], we have
y(t)≥ kα(x
∗(t–τ) –η)2
1 +β(x∗(t–τ) –η)2y(t–τ) –dy(t).
Choose a numberm2=q1[eμη(t+τ+T¯1)maxt∈[0,T]vη(t)]–1> 0. It is clear that, fort∈[t+
¯
T1,t+τ+T¯1],
m2eμηtvη(t)≤q1≤y(t),
and fort∈[t+T¯1,t¯],
m2eμηtvη(t)≥m2eμη(t+T¯1) min
t∈[0,T]vη(t) =
q1mint∈[0,T]vη(t) eμητmaxt∈
[0,T]vη(t)
=q2> 0.
The comparison theorem implies that, fort∈[t+T¯1,¯t],
y(t)≥q2.
Thusy(t)≥q2fort∈[t,¯t].
5 Numerical simulation
In this section, we give phase portraits of system (1.1) to illustrate the above theoretical analysis using numerical simulations. We consider the following delayed model with im-pulse: cluded that ifT<T1∗, then the predator-free periodic solution is globally attractive, and if
T>T2∗, then the predator will persist. However, ifT∈(T1∗,T2∗), then they could not deter-mine whether the predator can persist or not. In the present paper, we completely solve this question.
By Eq. (3.3), for system (5.1), the following eigenvalue problem can be obtained:
g1(t)
+∞
0.5
u(t–δ)e–0.24(δ–0.5)dδ=R0u(t), u∈CT, (5.2)
whereR0andu(t) are the eigenvalue and eigenvector of Eq. (5.2), respectively, and
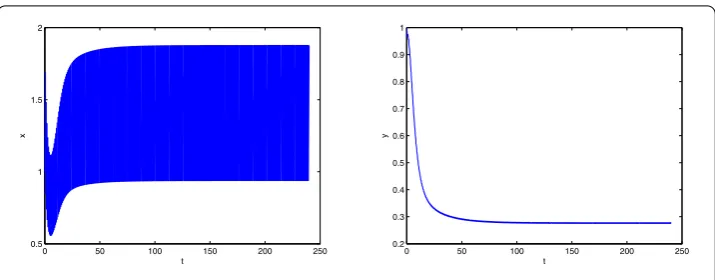
Figure 2Time series of the solution of system (5.1) withT= 0.7
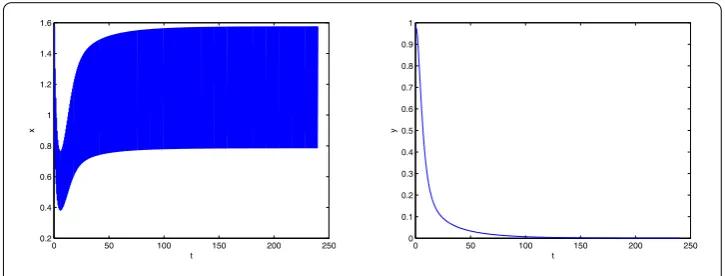
In system (5.1), letT= 0.6∈(T1∗,T2∗). We compute the basic reproduction numberR0 numerically to get R0≈0.8590 < 1. We also numerically compute the thresholdT∗≈ 0.6445. It is obvious thatT= 0.6 <T∗, that is,R0< 1. By Theorem4.1we obtain that the predator-free periodic solution is globally attractive. Figure1illustrates our theoretical results.
LetT= 0.7∈(T1∗,T2∗). We numerically compute the basic reproduction numberR0≈ 1.1250 > 1. It is clear thatT= 0.7 >T∗, that is,R0> 1. By Theorem4.2we obtain that the predator will become permanent. Figure2illustrates our theoretical results.
6 Conclusion
In this paper, we mainly discuss the extinction and permanence of the predator for system (1.1). Using the basic reproduction number coming from epidemiology, we may find the threshold valueR0such that ifR0< 1, then the predator is extinct, whereas ifR0> 1, then it will persist. Thus we improve the results of [1]. As far as we know, this is the first paper employing this approach of [13] in ecology.
Acknowledgements
We would like to thank the anonymous referees very much for their valuable comments and suggestions.
Funding
The research was supported by the National Natural Foundation of China (11271371, 51479215, 11571324).
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
Both authors contributed equally to the writing of this paper. Both authors read and approved the final manuscript.
Author details
1Department of Mathematics, North University of China, Taiyuan, P.R. China.2School of Mathematics and Statistics,
Central South University, Changsha, P.R. China.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Received: 14 May 2018 Accepted: 19 November 2018
References
1. Li, S.Y., Liu, W.W.: A delayed Holling type III functional response predator–prey system with impulsive perturbation on the prey. Adv. Differ. Equ.2016, 42 (2016).https://doi.org/10.1186/s13662-016-0768-8
4. Murdoch, W.W., Briggs, C.J., Nisbet, R.M.: Consumer-Resource Dynamics. Princeton University Press, Princeton (2003) 5. Ruan, S.G.: On nonlinear dynamics of predator–prey models with discrete delay. Math. Model. Nat. Phenom.4,
140–188 (2009)
6. Hui, J., Zhu, D.: Dynamic complexities for prey-dependent consumption integrated pest management models with impulsive effects. Chaos Solitons Fractals29, 233–251 (2006)
7. Liu, B., Zhang, Y., Chen, L.S.: The dynamical behaviors of a Lotka–Volterra predator–prey model concerning integrated pest management. Nonlinear Anal., Real World Appl.6, 227–243 (2005)
8. Zhang, H., Georgescu, P., Chen, L.S.: An impulsive predator–prey system with Beddington–Deangelis functional response and time delay. Int. J. Biomath.1, 1–17 (2008)
9. Shao, Y.F., Li, Y.: Dynamical analysis of a stage structured predator–prey system with impulsive diffusion and generic functional response. Appl. Math. Comput.220, 472–481 (2013)
10. Liu, X., Ballinger, G.: Boundedness for impulsive delay differential equations and applications to population growth models. Nonlinear Anal., Theory Methods Appl.53, 1041–1062 (2003)
11. Leonid, B., Elena, B.: Linearized oscillation theory for a nonlinear delay impulsive equation. J. Comput. Appl. Math.161, 477–495 (2003)
12. Yan, J.: Stability for impulsive delay differential equations. Nonlinear Anal., Theory Methods Appl.63, 66–80 (2005) 13. Bai, Z.G.: Threshold dynamics of a time-delayed SEIRS model with pulse vaccination. Math. Biosci.269, 178–185
(2015)
14. Garrione, M., Rebelo, C.: Persistence in seasonally varying predator–prey systems via the basic reproduction number. Nonlinear Anal., Real World Appl.30, 73–98 (2016)
15. Pielou, E.C.: Introduction to Mathematical Ecology. Wiley-Interscience, New York (1969)
16. Paul, G., Hsieh, Y.H., Zhang, H.: A Lyapunov functional for a stage-structured predator–prey model with nonlinear predation rate. Nonlinear Anal., Real World Appl.11, 3653–3665 (2010)
17. Bate, A.M., Hilker, F.M.: Predator–prey oscillations can shift when diseases become endemic. J. Theor. Biol.316, 1–8 (2013)
18. Xu, R., Tian, X.H.: Global dynamics of a delayed eco-epidemiological model with Holling type-III functional response. Math. Methods Appl. Sci.37, 2120–2134 (2014)
19. Bacaër, N., Guernaoui, S.: The epidemic threshold of vector-borne diseases with seasonality. J. Math. Biol.53, 421–436 (2006)