.:
’ (ISSN 0022-409X)
JOURNAL
OF
r
I.’ i .RANGE MANAGEM,ENT
PUBLICATION OF THE
SOCIETY iOR RANGE MANAGEMENT
.
JOURNAL OF
RANGE MANAGEMENT
Publtshed bimonthly-January March, MayJuly. September, November-by the Society for Range Management 2760 West Fifth Avenue
TABLE OF CONTENTS:
Vol. 32, No. 6, November 1979
Denver, Colorado 60204
Copynght 1979 by the ARTICLES
Soctety for Range Management 412
Managing Editor FLOYD E. KINSINGER
2760 West Fifth Avenue Denver, Cola. 80204 Editor
418
REX D PIEPER
Dep Animal and Range Science New Mexico State Unlvensty Las Cruces. N Mex 88003
422 430
Diets of Sheep, Angora Goats, Spanish Goats and White-tailed Deer under Excellent Range Conditions by F.C. Bryant, M.M. Kothmann, and L.B. Merrill Nematode Density and Biomass in an Annual Grassland Ecosystem by Diana W. Freckman, Don A. Duncan, and Jeanne R. Larson
Supplementary Feeding of Range Sheep by D.L. Michalk and D.G. Saville Evaluation of Fall Burning on Bighorn Sheep Winter Range by James M. Peek, Robert A. Riggs, and Jerry L. Lauer
Book Review Editor PAT 0 CURRIE
Science 8 Education Admln
Livestock and Range Research Statlon Route 1. Box 3
Miles City. Montana 59301 Copy Editor
PATRICIA G SMITH 2760 West Ftfth Avenue Denver, Colo 80204 Editorial Board 1977-79
JAMES 0 KLEMMEDSON. Tucson, Anz GEORGE W. SCOTTER. Edmonton, Alta M JOE TRLICA. Fort Collins. Co10 J ROSS WIGHT. Logan, Utah 1978-80
453
457 &3
Performance Evaluation of Water Harvesting Catchments by G.W. Frasier, K.R. Cooley, and J.R. Griggs
DONALD A KLEBENOW, Rena. Nev JAMES T NICHOLS, North Platte, Neb MICHAEL D PITT. Vancouver, B C RONALD E SOSEBEE. Lubbock, Tex 1979-81
467
ROD BOVEY. College Statton. TX NEIL FRISCHKNECHT. Provo. UT
470
Evaluation of Wire Fences for Coyote Control by Bruce C. Thompson Structural Changes in Chamise (Adenostoma fizscicuhtum) along a Fire-induced Age Gradient by Philip W. Rundel and David J. Parsons
Variation in Winter Levels of Crude Protein Among Artemisia tridenta Sub- species Grown in a Uniform Garden by Bruce L. Welch and E. Durant McArthur Residual Effects of Phosphorus and High Rates of Nitrogen on Shortgrass Hange- land by Frank Rauzi
MARTIN VAVRA. Union, OR
HAROLD GRELEN. Plnevllle, LA TECHNICAL NOTES
475 A Portable Burner for Evaluating Effects of Fire on Plants by Carlton M. Britton INDIVIDUAL SUBSCRIPTION IS by membershtp In the
Society for Range Management
and Henry A. Wright
LIBRARY or other INSTITUTIONAL SUBSCRIPTIONS, 476 Protection of Instrument Wires in the Field by Ray W. Brown and James M. on a calendar vear basis. are $22 00 oostoald to all Collins
wuntrles Payment from outslde the Unlteb States should 478 F *
be remitted In US dollars by Internat!onal money order or au-y Rings and Wildlife by John G. Stelfox and David Stelfox draft on a New York bank
433
437
442
445
449
Yield and N Uptake by Seven Perennial Grass Species as Affected by High Rates of N Fertilizer by L.E. Lutwick and A.D. Smith
Effects of Agricultural Terraces on the Reestablishment of Bluestem Grasslands by Thomas B. Bragg and Larry J. Stephens
Survival Patterns of Major Perennials in Salt Desert Shrub Communities of South- western Utah by Neil E. West
Impact of a White Grub (Phyllophaga ctinita) on a Shortgrass Community and Evaluation of Selected Rehabilitation Practices by Darrell N. Ueckert
Cattle Activities and Preferences Following Strip Application of Herbicide by H.B. Shaw and J.D. Dodd
BOOK REVIEWS
CHANGE OF ADDRESS notlces should be sent to the 4x0 .&hOIOg>J Of Free-RunfiirzX Domesric Animds by G.W. Arnold and M.L. Dudzinski; Managlng EdItor. 2760 West Fifth Ave Denver, CO 80204,
no later than the first day of the month of Issue Copies lost
Big G’ume ofNorth Ameriw: Ecology und Mumqememcompiled and edited by John L. due to a change of address cannot be replaced unless Schmidt and Douglas L. Gilbert.
adequate nottce IS given To assure uninterrupted service.
provtde your local postmaster with a Change of Address 4x1 Index of Volume 32 Order (POD Form 3575). lndlcatlng thereon to guarantee
forwardlng postage for second class mail POSTOFFICE Please return entlre journal with address change BUSINESS CORRESPONDENCE. concerning subscrip- tons, advertlslng, reprints,, back Issues, and related mat- ters. should be addressed to the Managmg EdItor, 2760 West Fifth Ave., Denver, CO 80204
EDITORIAL CORRESPONDENCE, concerning manu- scripts or other edltonal matters, should be addressed to the Edltor, Dep Animal and Range Sciences. New Mextco State Unlv Las Cruces 88003
INSTRUCTIONS FOR AUTHORS appear each year In the March Issue. copies of these InstructIons are avallable from the editor
Diets of Sheep, Angora Goats, Spanish
Goats and White-tailed Deer under Excellent
Range Conditions
F.C. BRYANT, M.M. KOTHMANN, AND L.B. MERRILL
Abstract
A study was initiated in August 1975 to illuminate the dietary interrelationships among four kinds of small ruminants on the Edwards Plateau of Texas. Sheep diets were dominated by grass followed by browse, goat diets were nearly equal in percent grass and browse, and deer spent most of their feeding time on browse followed by forbs. Livestock diets were not as high in forbs as expected, indicating that abundant grass due to excellent range condition may relieve livestock pressure on forbs, leaving more forbs for deer. Increased feeding time by deer on grass regrowth, after livestock were removed from the study area in July for a 4-month deferment, indicated that grazing systems were favorable to deer by providing regrowth forage. However, where livestock and deer graze in common, there are periods of competition even under excellent range conditions. These periods were fall and winter for browse, winter and early spring for forbs, and anytime immature grass was available. Relative to deer, sheep probably were most competitive for forbs and succulent grass, while goats, particularly the Spanish goats, were most competitive for browse. Similar annual trends for diets for all kinds of animals for all forage classes suggested competition was primarily for palatable green forage.
Common use grazing refers to the use of a range by more than one kind of animal during the same year. Its advantages lie in the abilities of different kinds of animals to more thoroughly cover the range and make efficient use of all forage species. Economic benefits have been derived from both game animals and livestock through common use grazing on the Edwards Plateau of Texas (Merrill and Young 1954; Huss and Allen 1969). This area supports one of the largest white-tailed deer (Odocoileus virginianus L.) concentrations in the world and is a major source of production from sheep, Spanish goats, and Angora goats. Consequently, knowledge of dietary interrelationships between livestock and deer is important to proper management of these rangelands.
Previous studies on the Edwards Plateau have provided valuable insight into grazing habits of several kinds of animals
on Texas rangelands (McMahan 1964; Kothmann 1968;
Malechek 1970; Taylor 1972; McCollum 1972). This research was designed to complement previous studies. The objectives were: (1) to measure the forage available for grazing at regular intervals throughout 1 year; (2) to determine the botanical composition of diets of sheep, Angora goats, Spanish goats, and
The authors are assistant nrofessor. Ranee and Wildlife Management. Texas Tech University, and professors Department of Raige Science,
time of this research, Bryant was research assistant,
Texas A&‘&I University. At the Range Science. Texas A&M University.
Technical Article TA 14603 from the Texas Agricultural Experiment Station. Manuscript received November 11, 1978.
412
white-tailed deer at monthly intervals; (3) and to examine the dietary interrelationships among these four kinds of animals grazing on common range in excellent condition.
Study Area
The field research was conducted at the Texas A&M University Agricultural Research Station located on the Edwards Plateau 45 km southeast of Sonora, Texas. Historically, this resource area was a grassland steppe, but since being invaded by numerous woody perennials, it now contains a botanical mosaic of grasses, forbs, and browse (Krebs 1972). Topography is rolling with steep breaks along some drainages. Soils are stony clays and clay loams. Precipitation averages 61 cm with peaks in May and September.
The study area was located within a 33-ha pasture which was in excellent condition (L.B. Merrill, per. comm.). From 1948 to 1970, the pasture received light continuous grazing (16 animal units per 260 ha) with a 2: 1 :l ratio of animal units of cattle, sheep, and Angora goats, respectively. In 1970, it was root-plowed and seeded with both introduced and native grass species. Stocking rate was subsequently increased to 40 animal units per 260 ha and the Merrill grazing system (4-3; 12:4 mo) was applied.
Due to the long rectangular nature of the pasture (1.6 x 0.4 km), it was assumed that uniform grazing distribution could not be achieved by the different kinds of experimental animals used in the study. Consequently the study area was reduced to 8 ha by establishing a temporary enclosure on the west one-fourth of the pasture for 3 hr on the days when the experimental animals were released to graze. This tactic restricted the experimental animals used in this study to the 8 ha enclosure during diet sampling but did not prevent grazing of the area by the resident animals on the Merrill grazing system.
Selection of the study area based on (1) amounts and variety of grasses and forbs and (2) amounts, growth form, and age class of two important browse species which were plateau oak (Quevcus virginiana var. fusiformes (Small) Sarg.) and Vasey shin oak (Q. pungens var. vaseyana (Buckl.) C.H. Muller). Variety and abundance of the available forage generally allowed all animals to express dietary preferences. The browse stand consisted of 7 year regrowth following root plowing and had not been over-utilized to the extent that a browse line existed. Thus the browse stand had a relatively uniform growth pattern and was fairly productive of edible leaves and twigs. It was also within reach (less than 2 m tall) of all kinds of animals regardless of grazing behavior. This age class was also easier to quantify as to standing crop than were larger more mature trees.
Materials and Methods
Availability of herbaceous species and diets selected by the four kinds of animals were both sampled at monthly intervals for 1 year beginning August 20, 1975. The 2-day sampling for herbaceous species preceded diet sampling by no more than 4 days. Thus an
estimate of herbaceous species available for consumption was obtained just prior to diet sampling.
Estimates of standing crop of grass and forb species were obtained usingamodificationofthe techniqueproposed byGoebeletal. (1958) and modified by Durham and Kothmann (1977) and Bryant (1977). The area was sampled along randomly located transects by reading 100 plots (.25 m’) covered with a IO-cm grid to estimate cover. In a separate sample each species was clipped to determine weight per unit of cover. The IWplot sample size was sufficient as indicated by species: area curves (Oosting 1956).
All browse species were sampled for edible and total standing crop in August at the initiation of the study. Standing crop was estimated for plateau oak and Vasey shin oak again during November before frost, and in February before initiation of spring growth, and in May and July, 1976.
The sampling approach involved establishing the density and mean volume for each browse species and assuming these parameten were constant for I year. The oak densities were estimated using the corrected-point~distance technique of Laycock and Batch&r (1975). due to their aggregate growth pattern (i.e., clones). Densities of all other browse species were estimated using the point-centered quarter technique (Cottam and Curtis 1956). Regression equations were developed to predict the mean weight per plant using conical volume as the independent variable. Standing crop was estimated by multiplying density times weight per plant. A complete description of the technique was reported by Bryant and Kothmann (1979).
Diets of all four kinds of animals were sampled on 5 consecutive days each month. All study animals were fasted overnight prior to diet collections. Deer were grazed in the morning and livestock were grazed in the afternoon after an additional 6-hr fast. The four kinds of animals were treated similarly in that (I) all were maintained in a dly-lot, fed alfalfa hay during collection periods, and fed cottonseed cake and alfalfa hay between collection periods, (2) water and salt were always available, (3) all were fasted prior to each collection, (4) all were transported to and from the study area each collection day, and (5) all were allowed to graze for .75 to 1 .O hr each day. However, livestock were released for a 72.hr grazing period prior to the initial collection day of each month and deer were not. This allowed at least the sheep and goats to become familar with changes in vegetation, as recommended by Langlands (1967).
Livestock diets were obtained from four each of Angora goats, Spanish goats, and sheep fitted with permanent esophageal cannulae described by Taylor and Bryant (1977). Due to losses, eight Angora goats, eleven Spanish goats, and nine sheep were used to collect diet samples during the study. Each sampling day, the extrua samples were collected in screen-bottomed bags, thoroughly hand-mixed, and separated into two sub-samples. One was used for botanical and one for chemical analyses. Chemical analyses will be reported in another paper. The sub-samples for botanical analyses were composited by animals across days and by days across animals within each collection period. Botanical composition, based on percent of plant fragments, was determined for the three major forage classes (grass, forb, and browse) and for plant parts (leaf, stem, and head, or flower) within a forage class after Durham and Kotbmann (1977).
Twenty wild-trapped white-tailed deer fawns were raised and trained following the guidelines of Reichert (1972). Of the 20, only two survived that adapted to the rigors of transporting and handling and could still be allowed free range of the study area without the confines ofaleash (Fig. I). At project initiation one fawn was5 moold while the other was 4 mo. Thus the study utilized two deer through theirmostcritical life stage, weanling to yearling age (4 to 17 months). As an index to selection, the feeding minutes method was employed. The major limitations ofthis technique have been reported by Bjugstad et al. (1970). Feeding minutes has been found to overestimate browse and underestimate grass as diet constituents when compared with the esophageal fistula technique (Bryant 1977). Therefore, diet data from deer were excluded from statistical treatment.
Significant differences in the botanical composition of livestock diets were determined by analysis of variance. Differences in monthly and annual means were evaluated with Duncan’s multiple range test
(Dixon and Massey 1969). Due to the unbalanced nature of the data. a computerized general linear models program was used in analysis of variance (Barr et al. 1976).
Results and Discussion
Forage Availability
Standing crop of all 3 forage classes declined from August to February due to utilization, leaf fall, and decomposition of mature forage (Fig. 2). Grass growth exceeded utilization in March, April, and July, while forb growth exceeded utilization mApri1, May, and July. Forbstandingcropdeclined in Junedue to heat stress that dried up many of the succulent cool-season forbs. Grasses and forbs increased during July in response to 28 cm of rainfall received during the month, and because all livestock were removed on July I, 1976, to allow the pasture 4 mo of rest. Standing crop of herbage and edible browse in July.
1976, did not reach levels measured in August 1975, following a spring deferment.
Diet Composition
Sheep
801 G-G GRASS ?_,
GRASS
Standing Crop
5001 0
400-
FORBS
Standing Crop
300-
KG/HA -
200-
“ap
100 f 0 \ P \ 1
-\o- /
6 I I III1 1 II'
4oL--___-
Aug Sep Ott Nov Dee Jan Feb Mar Apr May Jun JLFig. 2. Standing crop of grasses, forbs, and edible browse from August 1975 to July 1976, on the study area at the Sonora Research Station.
A S 0 N D J F M A M J
MONTH
Fig. 3. Percentage of grass, forb, and browse fragments in sheep diets at monthly intervals throughout the year.
and selection of forbs during winter in Kothmann’s study. Other researchers have also reported that forb use by sheep increased as forb availability increased (Cook et al. 1967; Buchanan et al. 1972).
Sheep in this study appeared to shift their diet among the forage classes as the seasons changed (Fig. 3). During the growing season, grasses and forbs dominated the diets. From August through December, when grasses and forbs were mature, browse began to replace the herbaceous components of the diet. The dietary proportion of browse peaked in November and December. Kothmann (1968) reported the peak percent of browse in sheep diets occurred in January on bothgood and poor condition ranges. In this study, sheep compensated for a shortage of browse in January by increasing their consumption of grass from 39 to 54%.
Sheep were consistent in their use of grass. Percent grass in the diets did not differ significantly (P> .05) for 10 of the 12 months. Grass appeared to provide the base for the diet while forbs were selected opportunistically. Percent forbs in the diets closely paralleled the trend for availability of forbs. Browse was selected primarily during periods when herbage was mature, even though herbage comprised more than 90% of the available forage. This assumption was supported by the increased percent of browse in sheep diets during the June heat stress period. Leaf material comprised 96% of the sheep diets throughout the year (Table 1). Sheep utilized grass stems only in early fall and late spring while browse stems were consumed during winter. Dudzinski and Arnold (1973) reported leaf material in sheep diets was consistently greater than 93%) while Kothmann (1968) observed sheep diets to consist of more than 86% leaf material.
Table 1. Mean annual diet composition (percent) for livestock and deer grazed on the Texas A&M University Research Station at Sonora, Texas.
Kind of animal
Forage class Plant part
Grass Browse Forb Leaf Stem Flower
Sheep Angora goats Spanish goats White-tailed deer*
60” 22a lgQ 96a 3” 1”
4gb 40b 12b 96” 3” 1”
45c 42b 13b 95”
8 61 31 - “_”
t! -
nbc Means in the same column followed by the same letter are not significantly different (P<.O5).
* Deer were not included in mean separation.
Angora Goats
Angora goat diets contained similar amounts of grass (48%) and browse (40%) (Table 1). Malechek (1970) reported similar annual means for grass and browse components (50 and 39%, respectively) under light, continuous grazing. Annual trends reported by Malechek (1970) were remarkably similar to those found in this study. Angora goat diets from this study contained
12% forbs compared to 11% forbs reported by Malechek, even though drought conditions existed during Malechek’s study and forbs were not as available. Further, McMahan (1964) noted tame Angora goats increased their use of forbs when grazed in an exclosure that contained large amounts of forbs. It appears, as with sheep, excellent condition range also reduces the impact Angora goats have on forbs. The high protein diet fed to the experimental animals while in confinement may be partly responsible. However, Regelin et al. (1977) found no difference in forage preferences of tame, free ranging mule deer than those fed protein supplements.
Percentages of grass and browse in the diets were relatively constant throughout the year (Fig. 4). Major deviations from the annual means occurred in December for grass, and December and July for browse. Angora goat diets tended to shift back and forth from grass to browse.
Forbs increased in Angora goat diets in the spring (Fig. 4). The percentage of dietary forbs was higher in spring than fall even though availability in both seasons was similar. Mature forbs were obviously not as palatable. Malechek (1970) also found more forbs in Angora goat diets during spring, however, he reported higher percentages of forbs in diets during autumn than found in this study.
Leaf material comprised 96% of Angora goat diets (Table 1 ), which agrees closely with Malechek (1970). The highest percent of stems in the diets (7%) occurred during January and these were primarily grasses.
Spanish Goats
Grass in Spanish goat diets throughout the year was only slightly higher than browse with 45 and 42%, respectively. In July, percent of grass in diets was significantly higher than for the lowest month, December (Fig. 5). Browse was significantly lower in July than December. Percent grass and browse in the diets did not differ significantly among the other 10 months. Forbs were a minor component of the diets except in spring and summer.
a01
ii
IG-G GRASS I \
;
60:Gyxy
\
g
F 40-y=‘G,GjiTT
I ‘d
B _:
\/
\ \
0. B
.\/ 00.
0 \ ‘\B,’ \ I=
\...”
20- G .f .F.. . . . . .* ‘\
r.l
F
E .F’ . * \
5 *.
. . .*
*. ..F” ‘0
- *.
“F . . . . . . F . . . . . . F...
‘..+.... ..F.... : :* ..* *
I 1 1 I I I I I 1 I I I A S 0 N D J F M A M J J
MONTH
70 G-G GRASS
h 1 0 0- - % BROWSE
. . IG
F.*.***.F FORB
L F.
*..* *. 5 lo- ’ F. *..*
.* .F.. . . .** ‘.‘F.“’ *. .*
‘F.. . . “F...F :
L
II 11 11 ‘1 I”
A S 0 N D J F M A M J J
MONTH
Fig. 5. Percentage of grass, forb, and browse fragments in Spanish goat diets at monthly intervals throughout the year.
In the fall as grass availability declined, Spanish goats continued to select consistently high percentages of grass. Forbs did not contribute significantly to the diets from October through February. Forbs increased in diets during spring in response to greater availability, while browse in diets declined. Browse was lowest in the diets in July when Spanish goats selected actively growing grasses.
Spanish goat diets contained 95% leaf material (Table 1). Stem fragments were relatively constant in diets throughout the year for all forage classes. In May, 7% of Spanish goat diets was Texas cupgrass (Eriokhloa sericea (Scheele) Munro) seed- heads.
Deer
Overall, deer spent more time feeding on browse (61%) than any other forage class (Table 1). Forbs were intermediate (3 1%) while grasses were lowest (8%). Deer consistently fed on browse and forbs. Except for July, grasses apparently were of little importance to them (Fig. 6).
Other researchers have reported similar findings on the importance of browse and forbs in deer diets, along with the seeming unimportance of grass (McMahan 1964; Drawe 1968; Everitt and Drawe 1974). In contrast, McCollum (1972)
100’
HE. / 0‘ .\B
/ \ G-G GRASS
/ \
_ 80- / \ E--B BROWSE
B’
\
2 F*..** F FOR0
Q, Be’- ‘0
L” I
\
1’ \
.F.. x
*.
/ \ !; ‘.,. F
60- B---B ,
\ / ‘B :
..F’..” .*
F \ .:’
\::’ ! : \
.a.. ; \
“F .. \
. . . . t;
5 20-
II II 11 1 ’ 1 ’ f 1
A S 0 N D J F M A M J J
MONTH
Fig. 4. Percentage of grass, forb, and browse fragments in Angora goat diets at monthly intervals throughout the year.
Fig. 6. Percentage of time deer fed on grass, forbs, and browse at monthly intervals throughout the year.
found high use of grass by deer in winter and spring on the Edwards Plateau. In South Texas, deer used grass extensively only in winter (Drawe and Box 1968). Deer in this study were young and were not under nutritional stress. Older deer subjected to nutritional stress may in fact spend more time feeding on grass. However, since young animals require high protein levels (Ullrey et al. 1967) and grass is relatively low in protein (Fraps and Cory 1940), insignificant use of grass by young deer is nutritionally to their advantage.
The amount of time deer fed on forbs closely paralleled availability, especially during May, June, and July. Feeding minutes on browse was inversely related to availability, suggesting deer were forced to use browse rather than preferring :+
Il.
Deer diets responded to changes in grass availability only in July (Fig. 6). This was attributed to removal of livestock from the pasture in July and the regrowth of grasses. Deer did not have to compete with livestock for the nutritious grass regrowth available at that time.
Johnsongrass (Sorghum halepense (L.) Pers.), Texas winter- grass (Stipa Zeucotricha Trin. and Rupr.), and Carex spp. were the most important grass and grass-like species grazed by the deer during the study. Cane bluestem (Bothriochloa barbinodis var. barbinodis (Lag.) Herter) was important only in July when deer spent 8% of their feeding time on that species.
From August through October, Texas snoutbean (Rhynchosia texana Torr. and Gray), orange zexmenia (Zexmenia hispidu H.B.K.), copperleaf (Acalypha Zindheimeri Muell. Arg.), Texas virgins bower (Clematis drummondii Torr. and Gray), and Indianmallow (Abutilon incanum (Link.) Sweet) accounted for 80% of the total time deer spent grazing forbs. From November through March, primary forbs selected by deer were upright prairie-coneflower (rosettes) (Ratibidu columnar-is (Sims) D. Don.), Mexican sagewort (Artemisia Zudoviciana
Var. mexicana
(
WiZZd.) Fern. ), Texas virgins bower, and prickly lettuce (Lactuca serriola L.). From April through July, Texas snoutbean, Indianmallow, orange zexmenia, velvet bundle- flower (Desmanthus velutinus Scheele), chickenthief mentzelia (Mentzelia oligosperma Nutt.),
copperleaf, dayflower (Com- melina angustifolia Michx.), and milkvetch (Astragalus spp.) made up 80% of the deer’s forb grazing time. Deer selected from an average of 30 species of forbs per month, the highest number occurring in May (38 species).Of all the previously mentioned forb species, only prickly lettuce, day flower, and velvet bundleflower were similarly listed as important to deer in the studies of McMahan ( 1964) and McCullum ( 1972). Since McMahan ( 1964) did not report availability, it is difficult to judge the opportunity for selection afforded his deer. The important species listed in this study were unavailable in most, if not all, of the pastures sampled by McCollum (1972). Deer in this study were allowed a greater opportunity to express their preference for forbs than were deer in other studies on the Edwards Plateau.
Plateau oak dominated the deer’s browsing time in most months (Fig. 6). A notable exception was in March when deer spent 4 1% of their total time feeding on Vasey shin oak. This corresponded to the time it was actively producing tender, new growth twigs. Vasey shin oak normally initiates new leaves in April and use was expected to be high (Malechek 1970). However, a freeze in late March killed new growth leaves of browse plants. Interestingly, decadent leaves of Texas persimmon (Diospyros texana Scheele) from the previous year’s growth were highly selected in March, as deer spent 12%
of their time on them. Sugar hackberry (Celtis Zuevigata Willd.) was relatively important in spring. In August, March, April, and July, deer selected from 9 to 10 different browse species. McCollum (1972) and McMahan ( 1964) also found deer to rely on oaks as their primary source of browse.
Dietary Interrelationships Grass
All four kinds of animals exhibited similar trends for percentage of grass in their diets. Competition for grass was probably greatest between sheep and Angora goats since their diets were most similar with respect to percent grass. Diets of all four kinds of animals responded to both new growth and regrowth of grass as proportions increased in early spring and again in July. Therefore, it appears that competition was keen among livestock and particularly between livestock and deer when there were small amounts of high quality grass available. The competitive use of nutritious grass regrowth by cattle cannot be discounted because they are known to use large amounts of grass (Taylor 1972). However, their influence may be facilitative since they may remove mature growth thus making the regrowth more available. There was probably no competition among the four kinds of animals for dry grass because it was always abundant.
Forbs
The trends for forbs in diets and standing crop suggest competition was greatest in winter when forbs were scarce. Deer were particularly vulnerable during this period due to their apparent inability to use dry grass and to their high requirement for crude protein. This lack of winter forbs due to utilization and short term drought helps explain the poor antler development on yearling males harvested on the Sonora station the following autumn (Pat Reardon, per. comm.). Critical competition probably occurred in March as livestock grazed forbs to the extent that utilization apparently equaled net primary pro- duction. Later in spring, forbs increased in livestock and deer diets but availability also increased, suggesting competition was somewhat reduced compared with winter. Competition for mature forbs during late summer and fall was negligible since availability was high and forbs were only moderately important to diets of deer and livestock. Sheep appeared to be slightly more competitive with deer for forbs than were Angora or Spanish goats. Intensive management for deer suggests that sheep and goats could be restricted to small portions of the total deer range during this critical winter-early spring period to reduce their competition with deer. Future research should examine the economic practicality and possible trade-offs in sheep and goat management vs. deer management.
Browse
Trends of percent browse in diets were most similar among sheep and Angora goats, while trends for deer and Spanish goats were similar. All four kinds of animals competed for a limited quantity of edible browse in late autumn and early winter. In late winter when availability of browse was lowest, livestock consumed less browse, shifting primarily to dry grass. Deer, however, did not shift from browse to dry grass in late winter. Thus, the high proportions of browse in livestock diets prior to late winter, coupled with even small proportions in late winter, may have created a severe competitive influence on deer by reducing the availability of a forage class they so highly depended upon. Spanish goats continued to select higher levels of browse later in winter than sheep or Angora goats. ina:czt:ng
that Spanish goats may be more competitive with deer for edible browse during late winter. In addition, ongoing research at the Sonora station suggests Spanish goats may use more browse than Angora goats when pastures are in poor range condition (Charles A. Taylor, per. comm.).
Conclusions
These data indicate deer are favorably affected by proper grazing management which led to excellent range conditions on the study area. The ample grass availability may act to reduce the impact sheep and goats have on forbs, a forage class highly important to deer. Further, grazing systems increased the availability of grass regrowth to deer thus supplying a nutritious forage with reduced competition from livestock. It cannot be overlooked, however, that even though the pasture was in excellent condition, there were critical times of the year where competition occurred for certain forages. If deer are a primary management consideration, proper stocking of sheep and goats during critical times of the year on Texas ranges cannot be over- emphasized.
While significant variations occurred among the diets of the four kinds of animals, the overall similarity in trends for the selection of grass, forbs, and browse was surprising. Perhaps the competition among kinds of animals can best be summarized in terms of competition for green forage. Species preferences
were evident indicating that there would be a degree of com- plementarity .
Literature Cited
Barr, A.J., J.H. Goodnight, J.S. SalI, and J.T. Helwig. 1976. A user’s
guide to S.A.S. 76. SAS Institute Inc. Raleigh, N. Car. 329 p. Bjugstad, A.J., H.S. Crawford, and D.L. Neal. 1970. Determining forage
consumption by direct observation of domestic grazing animals. In Range and Wildlife Habitat Evaluation-A Research Symposium. USDA Forest Serv. Misc. Pub. 1147. 220 p.
Bryant, F.C. 1977. Botanical and nutritive content in diets of sheep, Angora goats, Spanish goats and white-tailed deer grazing a common pasture. Ph.D. Diss., Texas A&M Univ., College Station. 92 p.
Bryant, F.C., and M.M. Kothmann. 1978. Variability in predicting edible
browse forage from crown volume. J. Range Manage. 32:144-146. Buchanan, H., W.A. Laycock, and D.A. Price. 1972. Botanical and nutri- tive content of the summer diet of sheep on a tall forb range in soyth- western Montana. J. Anim. Sci. 35:423-430.
Cook, C.W., L.E. Harris, and M.C. Young. 1967. Botanical and nutritive content of diets of cattle and sheep under single and common use on mountain ranges. J. Anim. Sci. 26: 1169-l 174.
Cottam, G., and J.T. Curtis. 1956. The use of distance measures in phy- tosociological sampling. Ecology 37:45 1460.
Dixon, W.J., and F.J. Massey. 1969. Introduction to statistical methods. McGraw-Hill Book Co. 637 p.
Drawe, D.L. 1968. Mid-summer diets of deer on the Welder Wildlife Refuge. J. Range Manage. 21:164-166.
Drawe, D.L., and T.W. Box. 1968. Forage ratings for deer and cattle on the Welder Wildlife Refuge. J. Range Manage. 21:225-228.
Dudzinski, M.L., and G.W. Arnold. 1973. Comparisons of diets of sheep and cattle grazing together on sown pastures on the southern tablelands of New South Wales by principal component analysis. Aust. J. Agr. Res. 24:899-912.
Durham, A.J., Jr., and M.M. Kothmann. 1977. Forage availability and
cattle diets on the Texas coastal prairie. J. Range Manage. 30: 103-106. Everitt, J.H., and D.L. Drawe. 1974. Spring food habits of white-tailed
deer in the south Texas plains. J. Range Manage. 27:15-20.
Fraps, G.S., and V.L. Cot-y. 1940. Composition and utilization of range vegetation of Sutton and Edwards counties. Tex. Agr. Exp. Sta. B-586.33 p. Goebel, C. J., L. Debano, and R.D. Lloyd. 1958. A new method of deter- mining forage cover and production on desert shrub vegetation. J. Range Manage. 11:244-246.
Huss, D.L., and J.V. Allen. 1969. Livestock production and profitability comparisons of various grazing systems, Texas Range Station. Tex. Agr. Exp. Sta. B-1089. 14 p.
Kothmann, M.M. 1968. The botanical composition and nutrient content of the diet of sheep grazing on poor condition pasture compared to good condition pastures. Ph.D. Diss., Texas A&M Univ., College Station. 60 p. Krebs, C.J. 1972. Ecology: The experimental analysis of distribution and abundance. Harper and Row, Pub. New York, Evanston, San Francisco, London. 694 p.
Langlands, J.P. 1967. Studies on the nutritive value of the diet selected by grazing sheep. II. Some source of error when sampling esophageally fis- tulated sheep at pasture. Anim. Prod. 9:167-176.
Laycock, W .A., and C.L. Batcheler. 1975. Comparison of distance-
measurement techniques for sampling tussock grassland species in New Zealand. J. Range Manage. 28:235-239.
Malechek, J.C. 1970. The botanical and nutritive composition of goat diets on lightly and heavily grazed ranges in the Edwards Plateau of Texas. Ph.D. Diss., Texas A&M Univ., College Station. 61 p.
McCollum, J.M. 1972. The botanical composition of the diet of white-
tailed deer. M.S. Thesis, Texas A&M Univ., College Station. 51 p.
McMahan, C.A. 1964. Comparative food habits of deer and three classes
of livestock. J. Wildl. Manage. 28:798-808.
Merrill, L.B., and V.A. Young. 1954. Results of grazing single classes of livestock in combination with several classes when stocking rates are constant. Tex. Agr. Exp. Sta. P.R. 1726. 6 p.
Oosting, H.J. 1956. The study of plant communities. W.H. Freeman and Co. 440 p.
Regelin, W .L., R.M. Bartman, D.W. Reichert, and P.H. Neil. 1977. The influence of supplemental feed on food habits of tame deer. USDA Forest Serv. Res. Note R.M. 316. 4 p.
Reichert, D.W. 1972. Rearing and training deer for food habit studies. USDA Forest Serv. Res. Note R.M. 208. 7 p.
Taylor, C.A. 1972. The botanical composition of cattle diets on a 7-pasture high-intensity low-frequency grazing system. M.S. Thesis, Texas A&M Univ., College Station. 60 p.
Taylor, C.A., and F.C. Bryant. 1977. A durable esophageal cannula for
sheep and goats. J. Range Manage. 30:397-398.
Uhrey, D.E., W .G. Youatt, H.E. Johnson, L.D. Fay, and B.L. Bradley. 1967. Protein requirements of white-tailed deer fawns. J. Wildl. Manage. 3 1:679-685.
Nematode Density and Biomass in an Annual
Grassland Ecosystem
DIANA W. FRECKMAN, DON A. DUNCAN, AND JEANNE R. LARSON
Abstract
The nematode community structure was examined in grazed and ungrazed annual-plant rangeland on the US/IBP Grassland Biome San Joaquin Site located in the foothill-grasslands of central California. Nematode numbers and biomass were estimated from early growth to mature stages of the annual-plant vegetation. Nematode density was greater on the grazed area, predominately forbs, than on the ungrazed, mainly grass, area. A lower than normal precipitation appeared to be a limiting factor of nematode population density. The nematode trophic structure differed between the two sites, fimgivores and microbivores predominating on the grazed and ungrazed sites, respectively. Indications are that the critical factors controlling nematode density and community structure on this annual grassland are not grazing but soil moisture and temperature.
Yuen (1966), Schmitt and Norton (1972), Schmitt (1973), Stanton (1974), Yeates (1974), and Smolik and Rogers (1976) have studied the function of nematodes in belowground grassland ecosystems; however, knowledge of nematodes from the California annual grassland ecosystem is almost non- existent. Recent investigations in the mixed-grass prairie of South Dakota (Smolik 1974) provided evidence that nematodes constituted a significant pathway of energy flow, and that the application of nematicides significantly reduced nematode populations resulting in large increases in herbage production. Also, phytophagic nematode biomass was significantly higher in an ungrazed pasture than in a grazed pasture. A preliminary study in the shrub-steppe area of south-central Washington (Smolik and Rogers 1976) revealed large numbers of soil-dwelling nematodes but no consistent differences in nematode density or biomass between grazed, ungrazed, and burned areas.
In 1972, the annual grassland site of the United States International Biological Program (US/IBP), Grassland Biome, was established at the U.S. Forest Service’s San Joaquin Experimental Range, 40 km northeast of Fresno, California. For a 3-year period, data were collected on abiotic, producer, consumer, and decomposer components of the ecosystem (Duncan 1975). A preliminary soil sampling program was undertaken in 1974 to estimate the importance of nematodes at the San. Joaquin site. Soil samples were taken on two dates (March 13 and May 1, 1974) in the grazed and ungrazed
Authors are assistant research nematologist, Department of Nematology, University of California. Riverside; research range scientist and range conservationist, Pacific Southwest Forest and Range Experiment Station, Forest Service, U.S. Department of Agriculture, Berkeley, California, stationed at Fresno, California.
Authors wish to acknowledge the assistance of Sandy Barnard, Bambi Jones, and George McIlece. This paper reports on work supported in part by National Science Foundation Grants GB-41233X and BMS73-02027-A02 to the Grassland Biome. U.S. International Biological Program, for “Analysis of Structure. Function and Utilization of Grassland Ecosystems.”
Manuscript received June 18, 1978.
418
portions of the US/IBP site, refrigerated and air-freighted to Dr. J.D. Smolik. Plant Science Department, South Dakota State University, Brookings. Nematodes were extracted by the Christie-Perry (1951) method, and a correction of 73% for extraction efficiency was used. Results of this preliminary effort indicated nematodes could be important components in the functioning of the annual grassland ecosystem. Numbers of nematodes were considerably higher in the May sample than in March on the ungrazed area, with an average of 5 to 12 X
106/m2 to a depth of 60 cm for the two dates. l Biomass varied from about 623 to 1,309 mg/m2 (dry weight).
This limited study was conducted in 1976 to further investigate the importance of nematodes in a grazed and ungrazed annual grassland ecosystem. Importance in an ecosystem is often determined by measuring parameters such as nematode density and biomass at different seasons and making estimates of nematode metabolism and productivity.
Study Area
The San Joaquin Experimental Range is representative of the granitic soil area in lower foothill annual grasslands in central California. The area is the annual plant-oak woodland type, with scattered trees and brush. The herbaceous vegetation is made up almost entirely of annual plants. The nematode study area was within the US/IBP site, which is an open grassland area. The study area for nematode investigations was a uniform area of Ahwahnee coarse, sandy loam soil, moderately deep, on a 10% slope with northern exposure. The site was the open rolling slope described by Bentley and Talbot (195 I), which is intermediate in herbage production between the swale or gentle slope sites and the rocky, steep slope sites comprising most of the area on the Experimental Range. Yearlong grazing by cows and calves constituted the grazed treatment, which was separated from the ungrazed area by a barbed wire fence (Fig. 1).
The ungrazed area had not been grazed since 197 1.
Methods
Two plots of 1 square foot (30.48 cm2) each in the grazed and the ungrazed areas were selected at random for each of five sampling dates from January 2 1 to May 24, 1976. The annual vegetation in these plots during this period ranged from early vegetative in January to mature stages in May. Aboveground biomass was determined by clipping the vegetation to ground level for both grazed and ungrazed plots inside a square-foot frame at each sampling period. Current year vegetation was placed in a labeled paper bag, oven dried at 6O”C, and total dry weight determined. The vegetation was then sorted into species and groups: Bromus mollis, B. diandrus, Vulpia megalura, Erodium spp.
Trijolium spp. , Lotus purshianus miscellaneous legumes, and miscel- laneous forbs other than legumes. Litter from the prior years’ growth
’ Smolik, J.D. 1975. Unpublished results.
tom, Dorylamida, excluding Xiphinema spp, Longidorus spp., and
Trichodorus spp.; phytophagic nematodes, Tylenchida; and fungi- vote, Aphelenchus, Aphelenchoides, and Difylenchus spp. (Table IL
A fifth group was composed of nematodes which were damaged or unidentifiable. Numbers of nematodes were corrected for extraction efficiency of 26% and density of nematodes was calculated as number/mz. Body weights of the nematode population were determined by length and width measurements of 250-300 randomly selected nematodes and calculations were according to Andrassy (1956). Biomass was expressed as mg/mz. Results are based on analysis of four of the five samples, February to May. Extracted nematodes from January samples wx destroyed accidentally.
Results and Discussion
Precipitation during the study period was below normal. with
0
” t “a exception of the month of February (Table 2). and the monthly f ((, mean air temperatures followed the long-term means.
“ Herbage yield was considerably less than for the prior 3 years
on the same area. The herbage yield was highest, 359 g/m’, on the ungrazed area in April (Table 2). In terms of plant growth
Table 2. Herbage yield, composition, litter, and monthly precipitation at five sampling dates in a grazed end ungrazed grassland at the San Joquin Exprimental Range, 1976.
After the current year’s vegetation and litter from previous years ““grazed (g/m’) 106.0 163.0 211.0 359.0 326.0 were removed, three soil cores were taben from each plot. The soil Grasses(B) 99.8 IW.0 1ca.O 99.6 100.0 tubes, 5 cm in diameter, were driven into the soil to a depth of60 cm FcrbS (?a) 0.2 0.0 0.0 0.4 0.0 with a power driver. Two soil cores were used for nematode analysis
and the other for determination of soil gmvimetric water. Each core Litter
was divided into IO-cm segments, and soil temperatures were Grazed (8irn2) 326.0 162.0 205.0 219.0 344.0 measured in the center of each soil core segment with calibrated Ungrazed(gim2) 964.0 1182.0 679.0 500.0 477.0 thermometers. Then each segment was placed in a labeled plastic hag Precipitation
and sealed All soil core samples were collected between 9:oO a.m. ,975.1976(an) 0.41 13.64 4.17 2.84 0.05 20.70
and noon. 1934.1975 mean 8.53 8.76 7.42 4.90 1.40 22.48
Each IO-cm soil core segment for nematode analysis was mixed and (cm) a 50.cm3 subsample was placed on a Baamann funnel for 48 h. The
extracted nematodes were counted and identified as one of fourtrophic
groups based on feeding habits or relationship to known trophic group: stages, the January and February sampling dates represent early microbivores, mainly Cephalobidae and Rhabditida; omnivore-preda- slow growth; the March date was at the beginning of rapid gmwth, and the April date was near peak production. Virtually
Table 1. Tiwonomic list of nematodes at the San Joaquin Experimental all the annual plants were mature and dry on the May date. The
Range. effects of prior grazing plus the unfavorable weather conditions
during the growing season probably combined to result in yields at all sampling dates on the grazed area which were lower than
Plant Feeders Microbiwres: on the unerazed area.
The combination of prior use and abnormally low rainfall resulted in an unusual difference in botanical composition during the study and may have been responsible for differences in nematode density and biomass data obtained from 1974 to 1976. Vegetation on the ungrazed areas was almost entirely grasses (mainly Bromus diandrus and B. mollis), while on the grazed area it was mostly forbs (Erodium was the most abundant) for all but the first sample date (Table 2). Dry conditions resulted in very little legume growth in either treatment. Litter accumulation was much greater on the ungrazed area at all times.
February, when it was over 12% at the 0- 10 cm depth and approximately 10% at lower depths. The soil was relatively dry on all subsequent dates (Fig. 2) and was less than 1% water at the 0- 10 cm depth on the final sampling date in May.
Soil temperatures, expressed as a mean of temperatures at all depth increments, were always 1 to 2°C higher on grazed area (Fig. 2). Seldom was there more than 1” difference between the 0- 10 and SO-60 cm core segments in either treatment during the morning sampling periods. The largest difference between the top and bottom segments was on May 25, when the soil temperatures in the ungrazed area were 26.5”C, respectively in the grazed area.
Table 3. Nematode density and biomass in grazed and ungrazed grassland at O-60 cm depth at San Joaquin Experimental Range, 1976.’
Nematode density Biomass
(X 106/m2) _ ~~idm*)
Sample date Grazed Ungrazed Grazed Ungrazed
February 0.94 1.82 38.3 72.0
March 4.19 3.13 175.4 125.9
April 4.81 3.85 193.4 152.9
May 2.82 1.99 112.8 79.3
Mean 3.19 2.69 130.0 107.5
I Corrected for extraction efficiency.
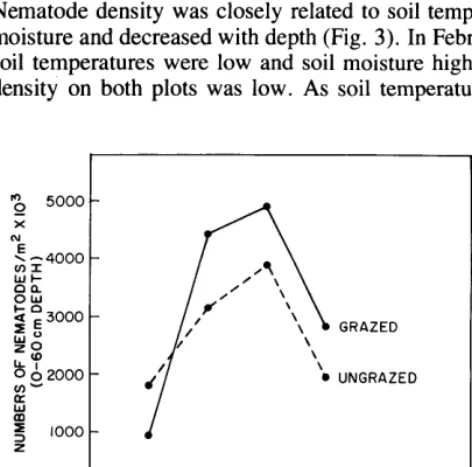
Total nematode density and biomass were highest on the grazed plot (Table 3, Fig. 2) at all sampling periods but February. The mean number of nematodes for the four sampling periods, corrected for extraction efficiency, was 3.2 x 1 06/m2 at
O-60 cm depth in the grazed plot and 2.7 X 10”/mg at O-60 cm depth in the ungrazed plot. Nematode biomass was 130 mg/m2 and 108 mg/m2 at 0 -60 cm on the grazed and ungrazed plots, respectively, and was at maximum in April on both plots. Nematode density was closely related to soil temperature and moisture and decreased with depth (Fig. 3). In February, when soil temperatures were low and soil moisture high, nematode density on both plots was low. As soil temperatures became
m 0 5000 t
GRAZED
UNGRAZED
TEMPERATURE OC
FEE MAR APR MAY
Fig. 2. Total nematode numbers and soil moisture and temperature in grazed and ungrazed grassland at the San Joaquin Experimental Range, California, 1976.
. GRAZED GRASSLAND
l FUNGAL FEEDERS X MICROBIVORES 0 OMNIVORE-PREDATORS 0 PLANT FEEDERS
5 80 t
UNGRAZED GRASSLAND z
1
O-IO IO-20 20-30 30-40 40-50 50-60
DEPTH (Cm)
Fig. 3. Vertical distribution of nematode trophic groups on grazed and un- grazed grassland at the San Joaquin Experimental Range, California, 1976.
warmer and moisture levels declined to 4% to 5% in March and April, there was an increase in numbers of nematodes. Maximum nematode metabolism, production, and energy turnover probably occurred during this period when the range of soil moisture and temperature was more optimal for nematode activity. Nematode activity and density may have been minimal in May because some nematodes are known to enter an inactive state of cryptobiosis, which is brought about by unfavorable environmental conditions such as low soil moistures. Freckman (1978), working in desert soils, found the nematode community to be cryptobiotic when soil moisture was below 2.7%, and it is possible similar conditions occurred during this study. Cryptobiosis enables nematodes to survive until favorable conditions of moisture and temperature return.
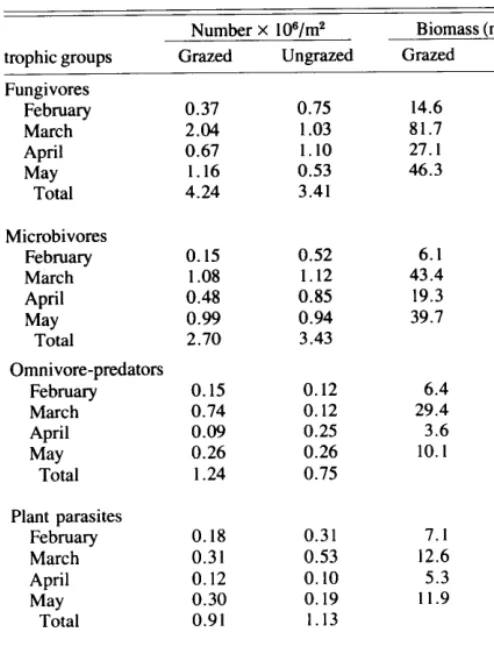
A comparison of this more comprehensive study with the 1974 survey shows a reversal in numbers of nematodes on grazed and ungrazed sites. In March, 1974, there was a greater number (7.35 x 106/m2, O-60 cm depth) and biomass (0.918 g/ma, O-60 cm depth) of nematodes in the ungrazed grassland. Results of the March 1976 sampling indicated a greater nematode density (4.19 X 1 06/m2, 0 - 60 cm depth) and biomass (0.175 g/m2, O-60 cm depth) on the grazed grassland. The trophic structure of the March, 1974, ungrazed and grazed area was respectively 80 and 60% saprovores, whereas in March,
1976, fungivores and microbivores represented the nematode community in the ungrazed area by 33 and 36% and in the grazed area by 49 and 26%, respectively (Table 4). Nematodes in all trophic groups were most abundant at the 0- 10 cm depth, and numbers declined with increasing depth on both grazed and
Table 4. Monthly numbers and biomass of nematode trophic groups cor- rected for extraction efficiency in grazed and ungrazed grassland at San Joaquin Experimental Range, 1976.
troDhic groups
Number x 106/m2 Biomass (mgjm2)
Grazed Ungrazed Grazed Ungrazed
Fungivores February March April May Total Microbivores February March April May Total Omnivore-predators February March April May Total
Plant parasites February March April May Total Unidentifiable February March April’ May
0.37 0.75
2.04 1.03
0.67 1.10
1.16 0.53
4.24 3.41
0.15 0.52
1.08 1.12
0.48 0.85
0.99 0.94
2.70 3.43
0.15 0.12
0.74 0.12
0.09 0.25
0.26 0.26
1.24 0.75
0.18 0.31 0.12 0.30 0.91 0.31 0.53 0.10 0.19 1.13 0.12 0.33 1.55 0.07
14.6 28.3
81.7 41.1
27.1 43.7
46.3 21.1
6.1 20.5
43.4 44.6
19.3 33.8
39.7 37.8
6.4 5.1
29.4 5.3
3.6 9.7
10.1 10.1
7.1 12.7
12.6 21.5
5.3 3.7
11.9 7.4
4.0 5.3
8.2 13.4
138.1 62.0
4.8 2.9
I Ninety-five percent of all April data represents nematodes were damaged when the refrigerator malfunctioned.
from O-IO cm depth which
ungrazed areas (Fig. 3). Omnivore-predators were the most numerous trophic groups at the 0 - 10 cm and 40 -50 cm depths. The percentage of plant feeders was greater at the 0 - 10 cm depth (52%) on the grazed plot than on the ungrazed plot (32%) and at the lo-20 cm depth on both plots; plant feeders were the most abundant trophic group.
On the basis of this study and the 1974 study, it would appear that the interaction of soil moisture and temperature appears to be a more important factor controlling the nematode populations and trophic structure than grazing. For example, the 1974 data were taken in a year of abundant rainfall, and soil moisture was not a limiting factor. Total numbers of nematodes were much greater in the March sampling in 1974 than in 1976 on both grazed and ungrazed plots. The highest nematode density in
1974 occurred on the ungrazed areas, where higher organic debris and soil microfloral activity probably provided a greater abundance and quality of food sources for the fungivorous and microbivorous nematodes. However, in 1976, although low soil moisture (+2.5%) limited total nematode density later in April and May, soil in the 0- 10 cm depth grazed area was always several degrees warmer (28.5”C) than the ungrazed soil (265°C) (Fig. 2). A temperature optimal for nematode activity and reproduction was reached much sooner in the grazed area in March when moisture was not limiting than in the ungrazed area. This may explain why the numbers and biomass were higher in the grazed area than in the ungrazed area in 1976. Evidence from two previous studies on the importance of nematodes on grazed and ungrazed grasslands have been
JOURNAL OF RANGE MANAGEMENT32(6), November 1979
inconclusive. Smolik (1974) found a greater nematode biomass on ungrazed pastures and Smolik and Rogers (1976) found no differences in nematode density and biomass between grazed, ungrazed, and burned shrub-steppe areas. This San Joaquin study further indicates that grazed or ungrazed grasslands are not the sole determinants of nematode density and biomass. Other factors influencing the nematode community appear to be abiotic (i.e., soil moisture and temperature).
Grazing may influence nematode populations indirectly by causing a shift in plant vegetation. For example, in 1974, grasses were predominant with 70% grass on the grazed plot and 88% grass on the ungrazed plot. By March, 1976, the grazed area consisted of only 19.5% grass, whereas the ungrazed area was 100% grass (Table 2). Forbs were the predominant vegetation in the grazed area. Other differences in nematode density between the 1974 and 1976 plant growing seasons at the San Joaquin site could be in part due to extraction methods, extraction efficiencies, and taxonomic and trophic groupings.
Conclusions
Results of this study, although of an exploratory nature and obtained in a season of unusually low rainfall, indicate soil-dwelling nematodes are an important component in the annual grassland ecosystem and deserve further, more detailed study, particularly on different sites with different soils and vegetation. In comparison, soil microarthropod biomass ranged from 53 -22 mg/m2 for 0- 10 cm depth samples taken in April and May, 19732. This was lower than the nematode biomass of
117 -40 mg/m2 for a similar depth in April and May, 1976. Smolik’s (1974) South Dakota studies showed more nematodes on grazed rangelands, while a Washington study (Smolik and Rogers 1976) revealed little difference between grazed and ungrazed areas. These 1976 California tests indicated more nematodes on a grazed area where the vegetation was mostly forbs than on an adjacent ungrazed area with almost exclusively grass vegetation. Nematode community structure varied with fungivores > microbivores > omnivore-predators > phyto- phages in the grazed grassland and microbivores = fungivores > phytophages > omnivore-predators for the ungrazed grassland.
Nematode numbers and biomass in 1976 were much lower than found in preliminary tests in 1974, when more than twice as much rainfall fell in the months of March and April. However, relative seasonal nematode densities were very similar to the usual seasonal pattern for aboveground live plant biomass of the annual plants at the San Joaquin site. Nematode numbers were low in winter when soil temperatures were low, even with relatively abundant soil moisture. Numbers rose and peaked in April with rising soil temperatures and intermediate soil moisture supplies. As temperatures rose more and soil moisture rapidly declined in May, nematode numbers dropped.
Literature Cited
Andrassy, I. 1956. The determination of volume and weight of nematodes. Acta. Zool. Acad. Sci. Hung. 2:1-15.
Bentley, J.R., and M.W. Talbot. 1951. Efficient use of annual plants on cattle ranges in the California foothills. U.S. Dep. Agr. Circ. No. 870. 52 p. Christie, J.R., and V.G. Perry. 1951. Removing nematodes from soil. Proc.
Helminthol. Sot. Wash. 18:106-108.
Duncan, D.A. 1975. The San Joaquin site of the Grassland Biome: Its re- lation to annual grassland ecosystem synthesis, p. 9-15. In: The California Annual Grassland Ecosystem: Symp. Proc. Amer. Sot. Agron., Anaheim, Calif., Jan. 30, 1975. Inst. Ecol., Univ. Calif., Davis.
2 Burdick, D.J. 1979. Unpublished results.
Freckman, Diana W .1978. Ecology of anhydrobiotic soil nematodes. p. 345- 357. In: Crowe and Clegg (eds.) Dried biological systems. Academic Press.
Inc.
Schmitt, D.P. 1973. Population fluctuations of some plant parasitic nematodes in the Kalso Prairie, Iowa. Proc. Iowa Acad. Sci. 80:69-7 1.
Schmitt, D.P., and D.C. Norton. 1972. Relationships of plant parasitic nema- todes to sites in native Iowa prairies. J. Nematol. 4:200-206.
Smolik, J.D. 1974. Nematode studies at the Cottonwood Site. US/IBP Grass- land Biome Tech. Rep. No. 25 1. Colorado State Univ., Fort Collins. 80 p.
Smolik, J.D., and L.E. Rogers. 1976. Effects of cattle grazing and wildfire on soil-dwelling nematodes of the shrub-steppe ecosystem. J. Range Manage. 29:304-306.
Stanton, N.L. 1974. Soil organisms of Thunder Basin Grassland. Atlantic- Richfield Co. Environmental Impact Analysis, Univ. Wyoming, Laramie.
Yeates, G.W. 1974. Studies on a climosequence of soils in tussock grass- lands. No. 2. Nematodes. N.Z.J. Zool. 1:171-177.