A FUNCTIONAL ANALYSIS OF HLA-DQ USING TRANSGENIC MODELS
ARLENE JANE PRATER
A thesis submitted in partial fulfilment of the requirements of the University of London
for the degree of Doctor of Philosophy.
October 1991 - October 1995
Transplantation Biology Group, MRC Clinical Sciences Centre, Royal Postgraduate Medical School,
ProQuest Number: 10106930
All rights reserved
INFORMATION TO ALL USERS
The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest 10106930
Published by ProQuest LLC(2016). Copyright of the Dissertation is held by the Author.
All rights reserved.
This work is protected against unauthorized copying under Title 17, United States Code. Microform Edition © ProQuest LLC.
ProQuest LLC
789 East Eisenhower Parkway P.O. Box 1346
Abstract
Contents
Page no.
Title page 1
D edication 2
Abstract 3
C ontents 5
List of Figures 12
List of Tables 15
A bbreviations 16
Chapter 1: General introduction
1.1 MHC molecules 2 0
- MHC class I heterodimers 21
- MHC class II heterodimers 22
- Peptide binding groove 23
- Interaction of MHC molecules with the T cell receptor (TCR), CD4 and 24 CDS co-receptors
- Genes within the MHC region 26
- Polymorphism in the peptide binding groove 28
- Significance of the polymorphism within the a l / p l domain of MHC class II 31
molecules and implications of the different distribution of hypervariable regions between isotypes.
1.2 Tissue distribution of MHC class II 39
- Modulators of MHC class II expression 39
- Evidence for selective expression of class II MHC isotypes 40
1.3 Regulation of MHC class II gene expression 48 - Cis-acting regulatory sequences and trans-acting DNA binding proteins 48
- Regulatory sequences 49
- DNA binding proteins 52
- MHC class II deficient mutants 56
- Post-transcriptional regulation 57
- Evidence for HLA-DQ specific transcription events 58
1.4 MHC molecules and T cell repertoire selection 64 - Cell types that mediate positive and negative selection 64
- Altered ligand model of selection 68
- What is the evidence for the involvement of MHC/peptide complexes 68 in selection?
- Peripheral tolerance 71
1.5 Peptide presentation by MHC molecules 75
- Antigen processing and presentation 76
- Binding of peptides to MHC class II molecules 78
- Peptide elution experiments/techniques 79
- MHC class II binding motifs 80
- Tissue specific peptides 82
- MHC allele/isotype-specific binding 82
- The role of disease associated polymorphic residues in peptide binding 84
1.6 Autoimmunity 90
- Pathology of autoimmunity 90
- Glutamic acid decarboxylase (GAD) as an autoantigen in IDDM 103
Aims of this thesis 109
Chapter 2: Materials and Methods
2.1 M aterials 110
- Chemicals 110
- Enzymes 111
- Restriction Enzymes 112
- Polymerases 112
- Buffers 112
- Media 113
2.2 M ethods 117
Mice and in vivo techniques 117
- Generation of transgenics 117
- Mice 117
- Immunisation of mice 117
Cellular immunological in vitro techniques 118
- Cell lines 118
- Generation of T cell lines 118
- Cell preparations 118
- In vitro proliferation assays 119
- In vitro restriction assays 119
- FACS analysis 120
M olecular biological techniques 121 - Preparation of DNA fragments for micro-injection 121 - Restriction digest and isolation of DNA from LMP gel 121 - Ligation of cDNA construct into pDOl-5 vector 121
- Transformation of E. Coli 122
- Mini-preps 122
- Maxi-preps 123
- Isolation /purification of DNA from tails and cell lines 123
- PCR methods 123
-RT-PCR 124
- Southern blot analysis 124
- RNA isolation and purification 125
- Northern blot analysis 126
- DN A sequencing 126
- Western blotting 126
Chapter 3: Generation and characterisation of HLA-DQ3+ and HLA-DQ6+ transgenic mice
3.1 Introduction 128
- MHC class I transgenic mice and rats 129
- MHC class U transgenic mice 131
- Is there a need for a species-matched CD4 molecule? 137
- Vectors driving transgene expression 138
- Aims of chapter 3 141
3.2 Results 14 3
GENERATION OF HLA-DQ TRANSGENIC MICE 143
- Use of the pDOI-5 transgenic expression vector 144 - Sub-cloning of the HLA class II cDNA's into pDOl-5 145 - Sub-cloning of HLA-DQAl *0102: trouble-shooting and PCR strategy 147 - Preparation of the pDOI-5/ -DQ cDNA cassettes for microinjection 148
CHARACTERISATION OF THE TRANSGENIC LINES 156
- Development of strategies for PCR genotyping of transgenic lines 156
- Southern blot analysis 156
- RNA detection 158
- FACS analysis of cell surface expression 158
- Thymic expression of HLA-DQ transgenes 159
3.3 D iscussion 176
- Expression patterns in mice carrying HLA class II transgenes with 176 endogenous promoters
- HLA class II transgenes driven off the H-2Eo^ promoter in pDOl-5 178
Chapter 4: T cell repertoire changes in HLA class II transgenic mice
4.1 Introduction 181
- Superantigens 181
- Superantigen/MHC/TCR interactions 184
- Analysis of Vp/Sag/TCR/MHC interactions using transgenic mice 187
- MTV mediated selection in mice expressing a hybrid MHC class II molecule 188 - MTV mediated selection in HLA class II transgenic mice 189
- Aims of chapter 4 191
4.2 R esults 192
- HLA-DQ expression alters the TCR repertoire in C57BL/10 mice 193 - HLA-DQ3 and HLA-DQ6-specific alterations in the repertoire 194 - Repertoire changes in the HLA-DQw6/cDNA transgenic mice 194 - Switching promoters results in subtle changes in the effects of an HLA- DQ3 195 transgene on the mouse T cell repertoire
4.3 D iscussion 211
- TCR Vp specific negative selection in HLA-DQ transgenic mice 211
- HLA-DQ mediated deletion at the double positive and single positive stage 211 of T cell development
- HLA-DQ3 and -DQ6 specific alterations in the repertoire 213 - A lack of HLA-DQ mediated Mtv-specific deletion of V p ll+ T cells 216 - No apparent evidence for HLA-DQ mediated deletion in DQ6/cDNA mice 218 - TCR Vp specific positive selection in HLA-DQ transgenic mice 220 - MHC class II mediated positive selection of CD8+ T cells 223
Chapter 5: HLA-restricted responses in transgenic mice
5.1 Introduction 2 2 6
- Introduction 226
- Aims of chapter 5 232
- HLA-restricted epitopes of MBP in MS 233
- HLA-restricted epitopes of GAD in IDDM 236
- HLA-restricted epitopes of TPO in thyroid autoimmunity 238
5.2 Results 2 4 0
- T cell responses of HLA-DQ3/Gen (DQB1 *0302) and -DQ6/cDNA 240 (DQB1 *0602) transgenic mice to immunisation with a nominal peptide
immunisation with a panel of human GAD^s peptides.
- Responses of HLA-DQ6/cDNA (DQB 1 *0602) transgenic mice and controls 242 to immunisation with a panel of 17 human MBP peptides.
- Responses of HLA-DQ6/cDNA (DQB 1 *0602) transgenic mice and controls 243 to immunisation with a human MBP peptide 13-32
- Evidence that HLA-DQ6 specific binding in vitro of TPO peptide does not 244 necessarily correlate with HLA-DQ6 (DQB 1*0602) or (DQB 1*0604) restricted T cell responses in vivo
5.3 D iscussion 2 5 9
- HLA-DQ transgenic mice are capable of mounting transgene restricted 259 T cell responses
- An epitope of GAD^S spanning residues 505-519 is preferentially 260 presented by HLA-DQ3+ transgenic mice
- Changes in the level of T cell responsiveness to specific MBP peptides 262 associated with the introduction of an HLA-DQAl *0102/DQB 1*0602
transgenic heterodimer
- What mechanism could induce an auto-aggressive T cell response to 265 GAD or MBP?
- Determinant spread may expand the autoimmune repertoire during 266 the course of the disease
- A role for HLA-DQ and -DR as susceptibility alleles in autoimmunity 268 - Determinant capture in autoimmune pathogenesis 271
Concluding Remarks 2 7 2
References cited 2 7 5
List of Figures Page no.
1.1 MHC gene organisation 33
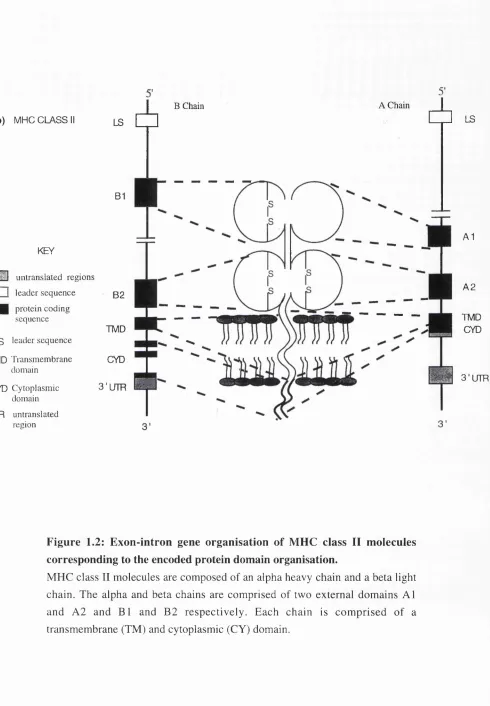
1.2 Exon-intron gene organisation of MHC class II molecules 35 corresponding to the encoded protein domain organisation.
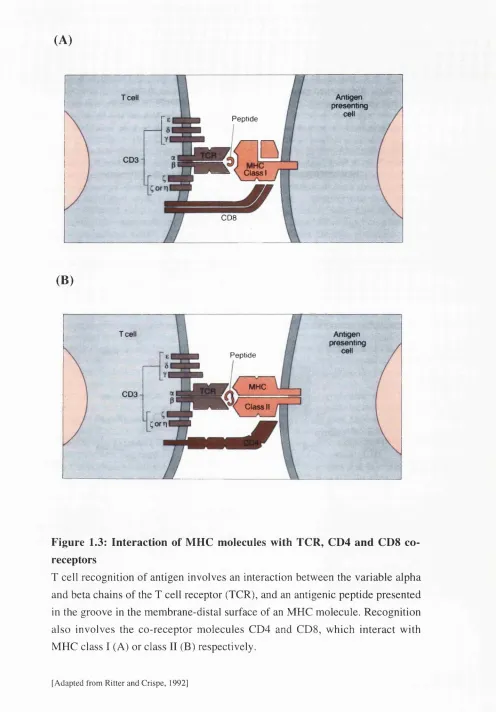
1.3 Interaction of MHC with TCR, CD4 and CDS co-receptors 36 1.4 Structure of the peptide binding groove of HLA-DR and HLA-DQ 37
showing the isotype-specific distribution of hypervariable regions.
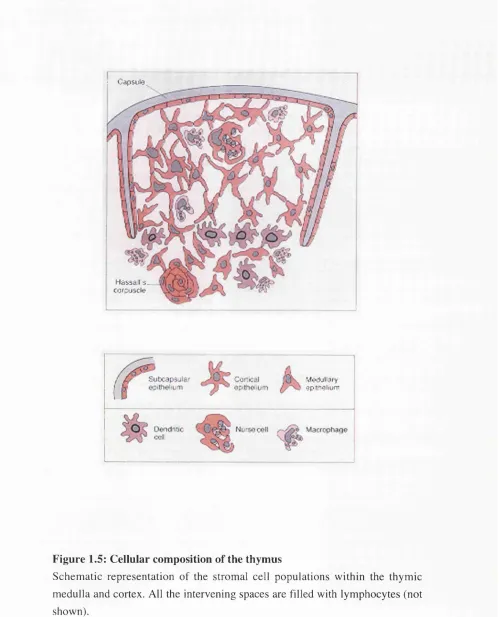
1.5 Cellular composition of the thymus. 46
1.6 Model of the MHC promoter 61
1.7 T cell maturation stages in the thymus 73
1.8 Summary of MHC class II antigen processing and presentation 86 1.9 A comparison of HLA-DR, -DQ and -DP peptide binding motifs 89 3.0 A schematic map of the pDOl-5 expression vector and sequences 151
of the sub-cloned HLA-DQ cDNAs.
3.1 A flow diagram (A-B) representing the steps involved in sub-cloning 154 of HLA-DQ cDNA (DQB 1*0602) into pDOl-5 and preparation for
micro-injection.
3.2 PCR amplification of the HLA-DQA 1*0102 cDNA to produce EcoR 1 155 restriction sites which were subsequently used to ligate HB20 into
pDOl-5 before preparation for micro-injection.
3.3.1 Diagrammatic representation of the primer pairs used in the typing of 161 transgenic mice.
3.3.2 Aligned amino acid sequences of HLA-DQ alleles showing location and 163 application of universal PCR primers used in typing of HLA-DQ+
transgenic mice.
3.4 A representative example of the detection of HLA-DQ transgenes 164 by PCR.
HLA-DQ6/cDNA transgenic mice and negative litter-mates.
3.6 Southern blot analysis of DNA extracted from tail biopsies of 166 HLA-DQ3/cDNA transgenic mice and negative litter-mates.
3.7 Southern blot analysis of DNA extracted from tail biopsies of 167 HLA-DQ3/Gen mice and negative litter-mates.
3.8 Northern blot detection of (A) HLA-DQA1*0301 and (B) 168 -DQB 1*0302 mRNA in DQ3/Gen transgenic mice and C57BL/10
negative littermates.
3.9 A representative example of RT-PCR detection of (A) HLA- 169 DQA1*0102 and (B) -DQB 1*0602 mRNA from DQ6/cDNA
transgenic mice and C57BL/10 littermates.
3.10 FACS analysis of cell surface expression of HLA-DQ3/Gen heterodimers 170 3.11 FACS analysis of cell surface expression of HLA-DQ3/cDNA 171
heterodimers
3.12 FACS analysis of cell surface expression of HLA-DQ6/cDNA 172 heterodimers
3.13 FACS analysis of the level of cell surface expression of HLA-DQ6 on 173 the positive control human B-LCL ‘PGF’
3.14 Immunocytochemical staining of an HLA-DQ3/Gen thymus 174 3.15 Western blot of thymi from HLA-DQ3/cDNA and -DQ6/cDNA 175
transgenic mice and negative control
4.0 A representative example of a FACS profile obtained after staining 198 mesenteric lymph node (MLN) cells from an HLA-DQ3/Gen transgenic mouse with anti-CD4/FITC, anti-CD8/FITC and anti-V pil/PE
antibodies
4.1 Analysis of raw data for TCR V p comparisons 199 4.2 Diagrammatic representation of the influence of the HLA-DQ6/Gen 202
transgene products on the mouse V p repertoire
transgene products on the mouse V (3 repertoire
4.4 Diagrammatic representation of the influence of the HLA-DQ3 transgene 209 products on the mouse V (3 repertoire
5.0 In vitro analysis demonstrating (A) preferential binding of TPO 245 peptide NP-7 (535-551) to an HLA-DQ6 molecule and (B) -DQ6 restricted stimulation of human self-reactive T cell clones from Grave’s thyroid infiltrates
5.1 T cell proliferative responses to ovalbumin in (A) HLA-DQ6/cDNA and 247 (B) -DQ3/cDNA transgenic mice and negative litter-mates
5.2 Restriction analysis of T cell lines from (A) HLA-DQ6/cDNA and (B) 248 -DQ3/cDNA transgenic mice primed with ovalbumin
5.3 Amino acid sequence of the human glutamic acid decarboxylase 249 (GAD65) N terminal (A) and GOGH terminal (B) peptides used in this
study
5.4 T cell proliferative responses of (A) B 10 and (B) HLA-DQ3/Gen 250 transgenic mice to a panel of 6 GAD^s peptides
5.5 An epitope of GAD spanning residues 505-519 is preferentially 251 presented by HLA-DQ3 + transgenic mice
5.6 Sequences of the overlapping human myelin basic protein (MBP) 253 peptides 111-126
5.7 T cell proliferative responses of HLA-DQAl *0102/-DQB1 *0602 254 transgenic mice (B) and negative littermates (A) after priming with a
panel of overlapping hMBP peptides
5.8 Responses of transgenic and non-transgenic mice to immunisation 256 with a peptide spanning residues 13-32 of human MBP
5.9 T cell proliferative responses of (A) C57BL/10 and (B) HLA-DQ6/cDNA 257 transgenic mice after immunisation with the TPO peptide NP-7
List of Tables Page no.
1.0 Phenotypic classification of the thymic micro-environment 47 1.1 A summary of the evidence presented in the text for HLA-DQ 63
specific gene regulation
1.2 Sequences of (A) H-2A^ and (B) H-2E^ associated peptides from 88 C3H/HeJ spleen
1.3 HLA and disease associations 108
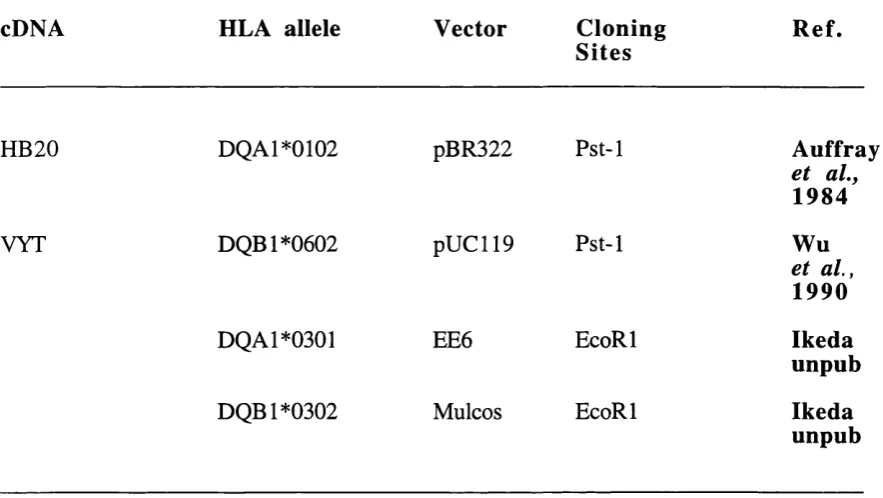
2.0 Antibodies directed against mouse and human MHC class II molecules 115 2.1 Antibodies directed against mouse v p , CD4 and CD8 molecules 116 3.0 Details of the original vectors, cloning sites and references for the 150
HLA-DQ cDNA clones used to generate HLA-DQ6/cDNA and -DQ3/cDNA transgenic mice
4.1 Strain distribution pattern of Mtv genomes in mice used in the present 197 study
4.2 Summary of v p deletion/Mtv specificity 197
4.3 TCR v p usage in HLA-DQ6/Gen transgenic mice and negative litter- 201 mates
4.4 TCR v p usage in HLA-DQ3/Gen transgenic mice and negative litter- 204 mates
4.5 TCR v p usage in HLA-DQ6/cDNA transgenic mice and negative 205 litter-mates
Abbreviations
AChR - acetylcholine receptor APC - antigen presenting cell AS - ankylosing spondylitis
BIO - C57BL/10
P21ÏI - beta 2-microglubulin BFA - brefeldin A
B-LCL - B-lymphoblastoid cell line BLS - bare lymphocyte syndrome bp - base pairs
CAT - chloramphenicol acetyl transferase CD - coeliac disease
CnTA - class II transactivator
CFA - complete Freund’s adjuvant
CLIP - class II invariant chain associated peptide CNS - central nervous system
CSF - cerebrospinal fluid CTL - cytotoxic T lymphocyte ddw - double distilled water dGuo - deoxyguanosine
EAE - experimental allergic encephalomyelitis BBV - Epstein Barr virus
ER - endoplasmic reticulum EtBr - ethidium bromide
FACS - fluorescence activated cell sorter FCS - foetal calf serum
FTTC - fluorescein isothyiocyanate FLU - influenza virus
FTOC - foetal thymic organ culture GAD - glutamic acid decarboxylase HA - haemagglutinin
HEL - hen egg lysozyme HLA - human leukocyte antigen HSP - heat shock protein HT - Hashimoto’s thyroiditis
li invariant chain IL-2,4,10 - interleukin-2,4,10
kb kilobases
kDa kilodalton
LCMV Lymphocytic Choriomeningitis Virus LMP low molecular weight protein/proteosome LTR-ORF - long terminal repeat open reading frame
mAh monoclonal antibody
MAG myelin-associated glycoprotein MBP myelin basic protein
MCC moth cytochrome C
MG myasthenia gravis
MHC major histocompatibility complex MLR mixed lymphocyte reaction
MOG myelin oligodendrocyte glycoprotein
MS multiple sclerosis
Mtv mammary tumour virus
NOD non-obese diabetic
OVA ovalbumin
pbg peptide binding groove PEL peripheral blood lymphocyte PBMC peripheral blood mononuclear cells
PCC pigeon cytochrome C
PCR polymerase chain reaction PLP proteolipoprotein
PPD purified protein derivative of M. tuberculosis
PV pemphigus vulgaris
RA rheumatoid arthritis
RFLP restriction fragment length polymorphism
rpHPLC - reverse phase high performance liquid chromatography
Sag superantigen
SA-PE streptavidin-phycoerythrin SEA Staphylococcal enterotoxin A SEE Staphylococcal enterotoxin B SEE systemic lupus erythematosis SSO sequence specific oligo
TAP transporter associated with antigen processing
TNF - tumour necrosis factor TPO - thyroid peroxidase
Abbreviations for amino acids
Amino Acid Three Letter One Letter Abbreviation Abbreviation
Alanine Ala A
Arginine Arg R
Asparagine Asn N
Aspartic Acid Asp D
Cysteine Cys C
Glutamine Gin Q
Glutamic Acid Glu E
Glycine Gly G
Histidine His H
Isoleucine He I
Leucine Leu L
Lysine Lys K
Methionine Met M
Phenylalanine Phe F
Proline Pro P
Serine Ser S
Threonine Thr T
Tryptophan Trp W
Tyrosine Tyr Y
Chapter 1 :
General introduction
1.1 MHC MOLECULES
MHC class I heterodimers
Classical MHC class I molecules are expressed at variable levels on most tissues and nucleated cell types except for corneal epithelium, the exocrine region of the pancreas, acinar cells of the parathyroid gland and villous trophoblast (Reviewed by Klein in 'The Natural History of the M H C , 1986). The gene products, class I antigens, consist of transmembrane glycoproteins expressed as heterodimers composed of an MHC encoded H (heavy) chain non-covalently associated with a non-MHC encoded invariant L (light) chain known as P2 microglobulin (p2m). The MHC class I H chain is composed of three extra-cellular domains a l , a2 and a3, a transmembrane region and a cytoplasmic tail, each encoded by separate exons (Sodoyer et aL, 1984; Strachan et al., 1984) Both human and murine HLA class I H chains have a molecular weight of 46Kd whilst the P2m L chains are 12 Kd. Each domain consists of approximately 90 amino acids. The alpha 2 and 3 domains contain intra-chain disulphide bonds enclosing loops of 63 and 86 aa respectively; the alpha 3 domain and P2m chain are composed of Ig-like single domains and are members of the Ig superfamily of genes. The a3 domain is highly conserved and this is thought to reflect the importance of this region’s potential interactions with the P2m molecule (Bjorkmanetal., 1987a, 1987b).
domain (Reviewed by Bjorkman and Parham 1990). These allele-specific polymorphisms determine the range of peptides each MHC molecule binds.
MHC class II heterodimers
Peptide binding groove
The a l and p2 domains associate to form the pbg of the class II molecule whose structure was initially predicted on the basis of conservation of sequence and distribution of polymorphic residues between MHC class I and class II molecules (Brown et al., 1988). X-ray crystallographic analysis has confirmed the major features of this prediction (Brown
et at., 1993; Stern et at., 1994). Overall the MHC class II structure is similar to that of class I molecules. With the exception of sequence alignment in strand 4 (residues a37- a43) of the a l domain, a hypothetical model of HLA class II based on the HLA-A2 structure is broadly correct. Differences include an altered position of the Ig-like p2 domain of HLA-DR 1 relative to the a3 of HLA class I, changes in the pbg so that it's ends are more open, peptides which bound as straight chains that projected out of both ends of the groove and more peptide binding pockets in the pbg (Brown et at., 1993).
tryptophan at position 167 and the salt bridge (from Glu55-Argl70) which 'close' one end of the HLA class I groove are absent in the HLA class II sequences.
The electron density in the peptide binding groove, representing a collection of endogenous peptides from the human B cells was straight with an extended conformation as expected. The data indicated that -15 peptide residues contact the HLA-DR molecule consistent with the mean length of the most abundant peptides bound to HLA-DR. The electron density was not weak in the middle as with HLA class I (which represents the peptides bulging out of the groove) but extended out of both ends of the pbg consistent with the observation that purified endogenous peptides from HLA class II vary in length from 12-24 amino acids and have ragged ends. One prominent peptide side chain appeared to bind in a hydrophobic pocket near residues a26, a31 and p86. The significantly smaller side chain (gly/val) at position 86 compared to the HLA class I homologue (tyr 171) helps create a deep pocket in the HLA-DR 1 groove surrounded by predominantly non-polar residues of both the a l and p i domains. This seems to be the position of a dominant peptide anchoring residue identified using substituted FLU peptides.
Interaction of MHC molecules with the T cell receptor (TCR), CD4 and CDS co receptors
clones whose receptors were derived from the same mouse strain and recognised the same peptide, but only one of which was alloreactive. Their findings demonstrated that, as previously predicted (Brown et a l, 1988), the CDR3 regions of both a and p chains were essential to all reactivities of the cloned T cell. However, they also demonstrated that interchanging the CDRl and CDR2 of the TCRa chain altered the fine specificity of MHC recognition such that a weak response became strong and vice versa. Further evidence for TCR/MHC interactions has come from MHC mutagenesis studies using characterised panels of moth cytochrome C (MCC) peptide specific T cells, MCC peptide analogues and H-2Ek mutant molecules (Jorgensen et a l, 1992). Residues in the H-2E^ a l and p i a - helices were mutated to amino acid residues with an opposite ionic charge or with an increase in the size of the side chain. The inhibitory residues were generally found to be centrally located in the a-helices, especially those shared by all three hybrids (a-65, p-64, -69 and -73/77) (Reviewed by Chien and Davis 1993).
residues play during interactions between CD4+ T cells and MHC class II molecules Sant
et al., (1993) generated interisotypic MHC class II p chains in which the p i domain was derived from H-2A and the P2, transmembrane and intracytoplasmic regions were from H-2E. The chimeric or wild type p chains were then transfected into L cells and formed a chimeric molecule with the wild-type a chain. The recombinant molecules were found to be defective in activation of normal T cell clones, deficient in the activation of primary alloreactive and superantigen-reactive T cells, but able to stimulate CD4" alloreactive T cell hybridomas. These results indicate that isotypic residues in the membrane proximal domains of MHC class II control CD4-linked accessory function.
Genes within the MHC region
The class I and II genes are the most polymorphic in the genome and many allelic forms have been identified. Nomenclature of the genes and their products has been classified in a series of workshops under the auspices of World Health Organisation (WHO). Each locus is identified by a suffix, ie. HLA-DQ, whilst additional terminology indicates whether the gene encodes an alpha or beta chain. The locus designation is then followed by an identity number (HLA-DQBl) and a specific allele sequence {HLA-DQB 1^^0601).
Non-polymorphic in man, this molecule shows limited variability in laboratory mouse strains: three alleles exist which differ by a single amino acid at position 85 of the protein sequence (Gates etaL, 1981).
Within the MHC class II region the H-2A alpha and H-2A beta genes pair to form the H- 2A heterodimer (homologous to human HLA-DQ), whereas the H-2 E alpha and H-2E
beta genes encode the H-2E molecule (the murine homologue of HLA-DR). The mouse gene homologous to the HLA-DP p chain, called PB (previously A B 5 ) is a pseudogene containing a 7 base pair deletion. No murine equivalent of the D P a chain has been found. Recently the homologue of HLA DOB , H -20B , (previously H-2AB2 ) was shown to produce a protein product and to form a heterodimer with O A . Some mice (b s f and q haplotypes) fail to express H-2E class II products. The b and s strains fail to make H -2Ea chains because of a deletion of 627bp in the promoter region. However, they express normal cytoplasmic levels of H-2Ep which can be utilised in chimeric H-2E molecules in F1 hybrids between b or s haplotypes and strains expressing H-2Ea. Mice of the f and q haplotypes fail to make both H-2Ea and H-2Ep chains. The H -2Ea chain defect appears to reside at the level of RNA splicing, whereas the -Ep defect has not yet been clarified (Mengle-Gaw and McDevitt 1983, and 1985; Mengle-Gaw etaL , 1984).
The HLA class II region spans approximately 1 kb with the -DP locus at the centromeric end, the -DR loci at the telomeric end and DQ situated between them (Hardy et a l, 1986; Trowsdale et aL, 1991). There is one HLA-DR alpha and some specificities have multiple
haplotypes. There are 5 group haplotypes designated HLA-DR 1, -51, -52, -8 and -53 (Bodmer et aL, 1995). Within HLA-DR 1 and -DR8 only one -DRB locus is expressed whereas there are 2 expressed in the -DR52 haplotype. To date 72 HLA-DR5 alleles have been sequenced. Five HLA-DQ genes can be detected in all haplotypes viz. HLA-DQAl, -A2, -Bl, -B2 and -B3. Recent studies have reported 14 alleles for H L A -D Q A l, 19 for
-D Q Bl but no evidence for expression of -DQA2, -B2 (DXl and DXB ) or -DQB3. Of the two pairs of HLA-DP genes encoded only -DPAl (8 alleles) and -DPBl (38 alleles) are expressed, -DP2 encoding pseudogenes (Bodmer et al,, 1995).
In addition to the MHC class II structural genes described above the class II region of the MHC contains genes involved in both the class I (endogenous) and class II (exogenous) antigen processing pathways (Trowsdale et at., 1990; Bahram et at., 1991; Geraghty et at,, 1992). In humans two genes, TAPI and TAP2 encode membrane transporters which form heterodimeric complexes whose role is to deliver antigenic peptides into the endoplasmic reticulum (ER), a necessary prerequisite for the loading and expression of MHC class I molecules (Spies et at,, 1992). Similarly, the LMP2 and LMP7 genes map to this region (el Kahloun et al., 1993) and are involved in the processing of peptides to generate fragments of suitable length which can then bind to MHC class I molecules. The mouse homologues of these genes also map to the H-2 region. Furthermore novel genes encoding the human HLA-DM and mouse H-2M molecules have been mapped to this region. These molecules affect antigen processing and presentation by MHC class II molecules. (Further detail in section 1.5, Antigen processing and presentation).
Polymorphism in the peptide binding groove
(presumably to serve the same function) whilst drawing attention to the variation that exists between isotypes and the possibility that these sequence and resultant structural changes may reflect subtly different roles for the various MHC class II isotypes.
The mechanism involved in generating the polymorphism of MHC genes is still the subject of debate. Random point mutations could contribute to the generation of allelic products, however, different MHC class I and II products differ at multiple residues, unlike other allelic proteins such as immunoglobulins which generally differ at single residues (Benoist et al,, 1983a). The mechanism postulated to account for the generation of multiple nucleotide substitutions in MHC alleles has been termed gene conversion/copy substitution. Donor gene sequences present elsewhere in the genome are thought to be copied and substituted in a non-reciprocal fashion onto recipient genes. For example, nucleotide sequence comparisons of H-2EB and the H-2AB t>ml2 chain from the bm l2 mutant (see section 1.4) have been taken as evidence that the bm l2 mutation was produced by the transfer of a stretch of 14-44 nucleotides from the exon encoding the H-2EB ^ first domain to the equivalent region of H-2AB (McIntyre and Seidman, 1984; Mengle- Gaw et al,, 1984). Other mechanisms, such as gene duplication and deletion and differential RNA splicing (Kappes et al,, 1984; Schenning et al,, 1984) could also contribute to the generation of diversity among MHC molecules.
Several independently isolated copies of HLA-DRA cDNA and genomic clones have been sequenced (Marsh and Bodmer 1995). So far only 1 polymorphism has been detected in the coding region, a conservative substitution of a Leu for a Val at position 217 (p 217). Similarly, of the 8 HLA-DPA alleles sequenced the polymorphism detected is very limited and does not cluster into any obvious regions of the a l domain. This contrasts sharply with the clustering of variability found in the a l domain of HLA-DQA alleles. HLA-DQA
Direct allelic comparison of the sequences of HLA-DRB 1, 3 and 5 published to date highlights 3 broad polymorphic regions. A highly variable and sharply delineated region at position 9-13, a broader and less polymorphic region around positions 26-37 and an area between residues 67-77. The low level of variability of the HLA-DPA a l domain is a feature that is shared with the adjacent HLA-DPB genes. Limited substitutions can be found between amino acids 8-11 and 55-57. Considered as a whole HLA-DP contains only a third as many substitutions as HLA-DQ whether nucleotide or amino acid substitutions are considered. H LA-D Q genes show extensive polymorphism in 4 discernible regions in the membrane distal domain between residues 26-30, 45-57, 66-77 and 84-94. Sequence variability is greatest between amino acids 66-77. In summary, most polymorphism occurs within the HLA-DR beta and -DQ alpha and beta chains. The areas of polymorphism do not coincide between these two isotypes, the NH2 terminal sequence is variable in HLA-DR and invariant in -DQ, whereas both molecules are variable at the COOH end of the p i domain, -DQ is polymorphic between amino acids 85-93 -DR is not. The high degree of variability detected in the HLA-DQp chain coincides with a highly polymorphic a chain. These regions of polymorphism in the first exon lead to distinct regions of HLA-DR and -DQ specific hypervariability in the peptide binding groove, (figure 1.4).
residues 11-15, 53-59 and 69-77 (Benoist et al., 1983a). Although these regions have relatively large amounts of inter-allelic variation there are no positions at which the amino acids are different for each of the sequenced haplotyes.
H-2E beta alleles for the haplotypes k, d, b, u and s have also been sequenced and compared (Mengle-Gaw and McDevitt 1983, and 1985; Mengle-Gaw et at., 1984). The variability amongst these alleles is again largely restricted to the membrane distal (pi) domain. Discrete regions of variability have been identified between residues 2-13, 27-34, 67-75 and 86-92. Ap alleles of k, b and d haplotypes (Choi et at., 1983; Malissen et at.,
1983; Larhammer et at., 1985) also show distinct regions of hypervariability between residues 8-17, 60-66 and 85-89.
Significance of the polymorphism within the a l / p i domain of MHC class II molecules and implications of the different distribution of hypervariable regions between isotypes.
(a) HIA Complex
I
— III —I
H
\ B C
\
\ \
DP DQ DQ DRB DRA
/ /
/
/
/ 9 E
E A G F
C4B C4B C2 TNFA TNFB
(b) H-2 Complex
h H h
1 [
- I I
'
I - I I I H 1--- 1 --- 11 Qa/T Ia ____ I 11 c o m p l e x 1
1 / / l / J / /
1 1 1 1
K A B A A EB E A /
/ /
/ /
/ B
\ D L Q1
\
\
\
\
QIOTIa
C4 C2 TNFA TNFB
(c) MHC Class II Gene organisation
/ / W
DP DNA
L M P - 2 T A P - 1 L M P - 7 T A P - 2
DMA DMB DOB DOB DQA
Mouse
DRB DRA
Figure 1.1: MHC gene organisation.
(a) A simplistic view of the organisation of the human HLA complex. (b) A simplistic view of the murine H-2 complex.
(c)A more detailed view of the organisation of genes within the human and murine MHC class II region. The class II region encodes genes involved with class I antigen processing and presentation ie. LMP 2 and 7 and TAP 1 and 2 and class II antigen processing and presentation ie. HLA-D M A and HLA-DMB. Pseudogenes are not illustrated.
(b ) MHC C L A S S II
KEY
I B untranslated regions I I leader sequence H protein coding
sequence
LS
B1
B 2
LS leader sequence
TTMD
TMD Transmembrane domain
CYD
CYD Cytoplasmic domain
3'LTTR
LTTR untranslated region
A C hain
I
B Chain
LS
A l
A 2
TMD CYD
I 3 'U T B
Figure 1.2: Exon-intron gene organisation of MHC class II molecules corresponding to the encoded protein domain organisation.
(A )
Antigw presenting
cell Tcell
Peptide
CD3-C lassl
CD8
(B)
Tcell Antigen
presenting cell
Peptide
MHC
CD3-C lasslf ;o r n
Figure 1.3: Interaction of MHC molecules with TCR, CD4 and CD8 co receptors
T cell recognition of antigen involves an interaction between the variable alpha and beta chains of the T cell receptor (TCR), and an antigenic peptide presented in the groove in the membrane-distal surface of an MHC molecule. Recognition also involves the co-receptor molecules CD4 and CDS, which interact with MHC class I (A) or class II (B) respectively.
(A) 5P 7 3 i83 165 59 69 73 47
93 34 Q
29
,35E 9 2 0
,153 • « C h a i n
6E52 o p c h a i n
No o f d ifferent amino a c id s a t a site
16 E 29
' 87 E 83
88
'A 4 9 ' 62 68, fBO 72 45 46 22
C o n s e rv e d am ino acid re s id u e s a re indicated
20
(B)
92 93 ,4 321
7 3
i83
>65
47
0 3 4 29
« C h a i n 82
'29
P c h a i n
No Of differen t amino a c id s a t a site
Figure 1.4: Structure of the peptide binding groove of HLA-DR and HLA-DQ show ing the iso ty p e-sp ecific d istr ib u tio n o f hypervariable regions
(A) HLA-DR structure [based on the crystal structure of class I] showing polymorphic residues. Colours indicate the degree of polymorphism at each residue. Dashed boxes represent residues omitted due to their lack of homology with class I sites.
(B) HLA-DQ structure [based on the crystal structure of class I] showing polymorphic residues. Colours indicate the degree of polymorphism at each residue. Dashed boxes represent residues omitted due to their lack of homology with class I sites.
1.2 TISSUE DISTRIBUTION OF MHC CLASS II
W ith the development of | monoclonal antibodies (mAh) to individual HLA determinants it become possible to distinguish between HLA-DP, -DQ and -DR and to study MHC class II distribution in detail. The expression of MHC class II on lymphoid cells is a dynamic process. MHC class II expression can vary with physiological compartment, stage in differentiation and state of activation - all determining factors influencing T cell development and response. In most cases, however, MHC class II expression by many cell types (including B cells, macrophages and dendritic cells) is low or absent unless induced by some stimulus, usually cytokines (Houghton et al.,
1984; Collins et al., 1984; Pober et al., 1983).
Modulators of MHC class II expression
Many cytokines are capable of inducing MHC class II expression (Balkwill and Burke 1989). They generally act on specific cells and can function independently or in concert with other cytokines. The molecular response to fFN-yis the best studied. The latter is a potent inducer of MHC class II expression in macrophages and cells of the non-haemopoietic lineage (Houghton et al., 1984; Collins et a l, 1984; Pober et a l,
al., 1986). Other inducers include TNF-a and GM-CSF for monocytes and IL-10 for B cells (Pfizenmaier et at., 1987; Alvaro-Garcia et at., 1989; Go et al., 1990) In general TN F-a acts in synergy with IFN-y at the level of transcription (Arenzana- Seisdedos eta l., 1988). In addition, recent evidence suggests that while TN F-a has a stimulatory effect early after treatment it can also be inhibitory with prolonged treatment (Zimmer and Jones 1990). A number of other agents have also been shown to downregulate class II expression. IFN-y is an antagonist for IL-4 in B cells (Rousset et al., 1988), whilst glucocorticoids are inhibitory for expression in both macrophages and B cells (Dennis and Mond 1986). Prostaglandins may also be inhibitory under certain circumstances (Snyder etal., 1982).
Evidence for selective expression of class II MHC isotypes
There have been few attempts to analyse the expression of the individual MHC class II isotypes on specific cell populations, most work focusing on the expression of HLA- DR as being representative of all human class II molecules. However, MHC class II genes are never expressed equally. The different levels of expression of HLA-DR, -DQ and -DP occur during haemopoietic progenitor cell differentiation. HLA-DQ expression, in particular, is not always concordant with the other HLA class II molecules expressed early in progenitor cells. The regulatory mechanisms which control the locus specific expression of the class II genes are not fully understood, however, such discordant expression of DQ implies that regulatory mechanisms which are specific to this isotype must exist. The following is an outline of isotype specific expression of MHC class II molecules on various cell types.
HLA-DQ differs from that of -DR: the former is confined to thymic epithelial cells and to endothelial cells of the small intestine during the first 26 weeks of intrauterine life, whilst -DR attains its full tissue distribution after 26 weeks (Natali et al,, 1984). Studies in mice have revealed that the major difference between the two species is the appearance of MHC class 11+ epidermal dendritic cells; in man these appear in intrauterine life, whilst in mice they are only detectable in 5-day old neonates (Natali et al,, 1984). In both species, as well as in rabbits, rats and guinea pigs MHC class 11+ cells are first detected in the thymus at an early stage (Rouse et al,, 1979; Jenkinson et al,, 1980; Jenkinson etal,, 1981; Natali etal,, 1981a,b).
In the myelomonocytic lineage, MHC class II molecules have been detected on monocytes (Winchester eta l.,191^), normal myeloblasts (Winchester etal., 1977) as well as committed stem cells for granulocytes and macrophages (Cline and Billing, 1977; Sieff etal., 1982). The earliest cells showing monocyte/macrophage markers are detected at week 4-6 of fertilisation and at this stage are HLA-DR"; as the cells seed to the thymic cortex, lymph, spleen and bone marrow they become -DR+. Interestingly, HLA-DQ molecules are not detectable on the stem cells of any lineage, including those destined to differentiate into monocytes. As with B cells there appears to be a progression from HLA-DR+/DQ" to -DR+/DQ+ as the macrophages mature, terminally differentiated Langerhans and Kupffer cells for example expressing both HLA-DR and -DQ antigens (Janossy et al., 1986).
MHC class II expression by T cells shows considerable inter-species variation, unlike B cells and dendritic cells which constituitively express MHC class II molecules in all species studied so far (Kaufman et al., 1984; Kappes and Strominger 1988). Resting T cells are generally MHC class II negative and reports of extremely low numbers of HLA-DR+/DP+/DQ" resting T cells may be due to the presence of a few activated cells (Brown et al., 1984). Natural killer cells do not express detectable levels of MHC class II molecules. In humans resting T cells are negative whereas cells activated with anti-CD3 mAh have been shown in one study to express -1x10"^ HLA- DR, SxlO'^'DQ and 5xlO^ DP molecules per cell, however, the levels are still significantly below that of other human accessory cell populations (Robbins et al.,
H LA -D R" populations were observed. These patterns of HLA class II isotype expression were similar in CD2+, CD4+ and CD8+ subgroups of T cells. All equine and rat T cells express MHC class II molecules upon activation whereas in mice the evidence is controversial (Lorber et al., 1982; Singh et a l, 1984; Crepaldi et at.,
1986). Early work using cloned lines of alloreactive cytolytic and helper T cells which were examined for the expression of H-2A molecules using FACS analysis suggested that alloreactive mouse T cells could adsorb MHC class II molecules from the stimulating cell population (Lorber et at., 1982). However, evidence does exist that points towards the presence of endogenously synthesised MHC class II molecules on activated T cells (Evans et al., 1978; Ko et al., 1979). HLA-DR, -DQ and -DP molecules are differentially expressed on T cells after stimulation (Robbins e ta l.,
1988). In addition, the level of expression would appear to vary with stage of cell cycle. In recent years the most comprehensive study of MHC class II expression on T cell sub-sets during maturation has been that of Dutia et al., (1993) and Hopkins et al.,
(1993), although this focuses on sheep it gives an indication of the sequence of events. The results of this study showed that the expression of MHC class II by T cells is dependant upon the age and location of the cell. In foetal blood CD8+/yô cells were HLA class II negative and the expression on CD4+ cells was very low. As the cells mature they progress from being HLA-DQ"/DR" to -DQ"/DR+ and then -DQ+^DR^ so that all blast cells expressed a uniformly high level of -DQ and -DR with no differential expression of the class II products. Within the afferent lymph and node HLA-DQ+/DR+ cells typified recently activated cells; whereas resting and recirculating memory T cells in the efferent lymph and node were -DQ"/DR+
MHC class II expression in the thymus
-DR. Ishikura et al., (1987) attempted to clarify the thymic expression of these antigens by using well characterised mAbs They demonstrated that there were different distributions of these antigens in the human thymus. In the cortex both HLA-DR and -DQ were strongly expressed on thymic epithelial cells, with simultaneous expression of both antigens. On the other hand in the medulla HLA-DQ antigens had a more limited pattern of expression than -DR. Double staining revealed small numbers of dual positive (HLA-DR+"DQ+) cells in the medulla, which looked like thymic epithelial cells (TEC), indicating that at least some medullary TEC were double positive, although the presence of small numbers of HLA-DR'DQ“ and -DR+DQ" cells could not be excluded. The preponderance of HLA-DR over -DQ was consistently found in ex vivo tissues from individuals aged between 10 weeks - 2 years and could be attributed to very weak/no expression of -DQ antigens on dendritic cells and/or macrophages.
Furthermore, evidence now exists for isotype-specific distribution of an MHC class II molecule (H-20) in the mouse thymus. Karlsson etal., (1991) identified a novel class II molecule when they precipitated H -20 (previously H-2Ap2). Electrophoretic analysis demonstrated that the H-20P chain pairs with a novel a chain and that the heterodimer associates with the invariant chain. Preliminary work indicated that H -20 had a more limited thymic distribution than H-2A. H-2A was present throughout the medulla and on cortical epithelial cells. H-20p, in contrast, was limited to aggregates of cells in the medulla. Assessing the distribution of MHC molecules on thymic epithelium is difficult due to ambiguous results caused by staining of the dense network of MHC class 11+ bone-marrow derived cells, however, this problem can be overcome by examining a bone marrow chimeric thymus. In this case all the bone- marrow derived cells are of donor origin and the host MHC class II is restricted to the epithelial cells. Using bone marrow chimeras and MHC transgenic mice Surh et al
I Capsu^
H assa I â _
corcuscie
MedJiia/y
epreliurr
Figure 1.5: Cellular composition of the thymus
Schematic representation of the stromal cell populations within the thymic medulla and cortex. All the intervening spaces are filled with lymphocytes (not shown).
Table 1.0: Phenotypic classification of the thymic micro-environment
Cell type Phenotype R eferences
Cortical epithelial cells MHC class II hi Boyd et a l, 1977; de Maagd et a l, 1985; Wekerle et a l, 1980
Medullary epithelial cells MHC class n variable Bofill e ta l, 1985; F a.netal, 1983; Jenkinson et a i, 1981; Surh et
a l, 1993; van Ewijk et a l, 1980; von Gaudecker et a l, 1986
Macrophages MHC class n hi Boyd e ta l, 1993
Dendritic cells/ Interdigitating cells
MHC class n hi Boyd et a l, 1993
B cells MHC class II + Boyd e ta l, 1993
1.3 REGULATION OF MHC CLASS II GENE EXPRESSION
Polymorphisms between MHC class II molecules have been found to extend outside the coding regions to include variation in the promoters. The cloning of MHC class II genes coincided with the development of new techniques to examine gene regulation and this has lead to a rapid accumulation of information in recent years. While cytokine mediated attenuation and post-transcriptional controls are operative, studies have revealed that transcriptional control is the most important regulatory mechanism for MHC class II antigens, accounting for many of the complex expression patterns detected by immunological methods.
Cis-acting regulatory sequences and Trans acting DNA binding proteins
The transcriptional initiation system is composed of discrete DNA units (cis-acting sequences) which act as ligands for specific DNA binding proteins (trans-acting binding proteins) (Maniatis et a l.y 1987). These cis sequences can act independently or form co
operative units (Lin et al., 1990). Positive or negative regulation may be determined by the type of proteins which bind, the specific sequence, or its location in the promoter complex (Levine and Manly, 1989). Cis-acting elements have been described by examining the consequences of deletions on the in vivo activation of a reporter gene or its authentic transcript produced in vivo or in vitro. The nature of trans-acting DNA binding proteins is deduced largely from DNA binding assays and correlative expression in various cell types (Benoist and Mathis, 1990).
Regulatory sequences
Thirty base pairs from the start site of MHC class II transcription is a TATA box. Although not functionally significant in H-2Ea this element is essential for the correct transcript of DRA to be produced. Mutation or deletion abrogates expression in vivo and leads to the production of inaccurate transcripts in vitro. It may be that the TATA element serves as a focus for the binding of initiation factors ensuring that transcription proceeds from the correct point (Reviewed in Glimcher and Kara 1992).
The HLA-DRA octamer sequence 5 -ATTTGCAT-3' is a perfect match to those found in the promoter regions of Ig H and L chains, the human histone H2B gene and human and xenopus U1 and U2 small nuclear RNA genes (Parslow etaU, 1984; Mattaj etaL, 1985). Its importance is unclear because among both human and murine MHC class II genes,
Generally located -40 to -90 base pairs upstream of the start site of transcription the Y box is a 10 base pair motif (5'-CTGATTGGCC-3') defined by its conservation between the promoter regions of all class IIM H C genes. Y box elements from all class IIM H C genes have similar properties. Substitution mutagenesis in in vitro transcription analyses show that the Y box is necessary for correct expression in B cells and for IFN-y induction (Tsang et aL, 1990; Hume and Lee 1990;). The Y box may also determine the accuracy of transcriptional initiation since site-specific mutation in vivo (Sherman et aL, 1989b) and competition for Y box binding proteins in vitro cause transcription to initiate from a cryptic start site (Zeleznik-Le et aL, 1991).
Approximately 19-20 base pairs upstream from the Y box is an X box, a 14base pair motif whose sequence is generally more divergent than Y, (by 3-4 bases). The 19-20 nucleotide intervening X /Y spacer is highly conserved in length, although of variable sequence. As a conserved distance separates the two elements, it may be that their activities are linked. For instance it has been proposed that this spacer region is responsible for the alignment of the X and Y boxes on the helix in a position which allows correct conformation for binding with regulatory proteins (Dorn etal., 1987b; Vilen e ta l, 1991).
An X box sequence ( 5-CCCTAGCAACAGATG-3 ) is identified by its conservation between all class II MHC genes. Mutational analysis suggests that the HLA-DRA X box is necessary for expression in B cells and for IFN-y induction (Sherman et aL, 1989b; Tsang et aL, 1990) Site-specific competition for X box binding proteins in an in vitro
transcription assay causes transcripts to initiate from a cryptic start site similar to competition experiments for Y box binding proteins (Zeleznik-Le et aL, 1991) This suggests that an intact X box is required for accurate transcriptional initiation. HLA-DQB
and Strominger 1988 and 1989). The relevance of these elements in controlling the expression of other class II MHC genes has been less thoroughly studied.
The X2 box sequence (5 -ATGCGTCAT-3') overlaps with the 3' end of the consensus X box and the 5' end of the spacer region. X2 is homologous to the activator protein (AP-1) sites of other genes (Hume and Lee 1989; Tsang et aL, 1990). Not all MHC class II genes have an AP-1 like element; some have CRE (cyclic amp responsive elements) like elements, while others have neither. A 3 base pair point mutation in X2 reduces promoter function in B cells whether assayed by in vivo expression of a reporter gene (Lion et aL,
1990) or in vitro transcription (Zeleznik-Le e ta l, 1991). In contrast, another study found no effect on B cells using a broad X2 mutation. X2 may also have a moderate positive effect on HLA-DR expression in HLA-DR+ T cells and fibroblasts (Tsang et aL, 1990).
At -110 to -117 base pairs is a pyrimidine rich region (P region) between the X and Y boxes (Tsang et aL, 1990). This P region has weak positive function as defined in deletion mutagenesis, however, it considerably enhances transcription in heterologous promoter constructs in conjunction with the X or, X plus Y, elements (Tsang et aL, 1988). The P region does not appear to bind a specific protein per se, although certain X box binding proteins contact residues at the 3' end of this element (Reith et aL, 1988; Caiman and Peterlin 1988). Therefore it may serve to stabilise binding of these X box binding proteins.
The W box has been called by many names including W (for HLA-DRA, -DQB and H-2Eb), S (for all MHC), Z and Z2 (for HLA-DRA) and H (for H-2Aa and H-2Ea) (Boss and Strominger 1986; Tsang et aL, 1988; Benoist and Mathis 1990; Tsang et aL,
et aL, 1990; Tsang et aL, 1990), Within -136 to -118 base pairs of the W region several interesting motifs exist. Two of these, the S-like sequences (GGACCC = SRV2/Z, GGACAC = SRV1/Z2), are shared between all MHC class II genes and were first described by Servenius et aL, (1987). The third motif is identified as an inverted sequence which shows homology to the HLA-DRA X box (Cogswell et aL, 1990; Cogswell et aL,
1991). Each of the above elements may control expression in B cells whereas IFN-y induction occurs predominantly through SRV2/Z (Tsang et aL, 1990). While only HLA-DRA has a repeated SRVl/Z sequence, inverted X-box like sequences are found just 5' to S boxes in HLA-DQB, H-2Eh and H-2Aa. SRV2/Z homologous sequences are also found in the HLA-DQB W box, H-2Eb W box and H-2Aa H box (Benoist and Mathis 1990). An additional element, with silencer activity, the V box, has recently been detected in the
HLA-DRA promoter, upstream of the W box. This element shows sequence homology with W and XI (Cogswell et aL, 1990) and encompasses a DNase 1 hypersensitive site in MHC class I expressing cells (Gonczy etal., 1989).
DNA binding proteins
The Y box contains an inverted CCAAT sequence capable of being recognised by at least two of the multiple CCAAT-binding proteins identified to date (Dorn et aU,1987a; Hooft van Huijsduijnen et al., 1987; Didier et al., 1988; Hooft van Huijsduijnen et al.,
1990; Zeleznik-Le et al., 1991). A cDNA clone encoding YB-1 was obtained by probing a library with double-stranded oligos containing the Y box (Didier etal., 1988). YB-1 has an estimated molecular weight of 35.4Kd and is encoded by a mRNA of 1.5Kb. Although YB-1 is found in both MHC class II positive and negative cells , it may act as a negative regulatory factor since the levels of YB-1 mRNA are generally inversely correlated with the levels of MHC class II expression in several cell lines (Didier et al., 1988).
The HLA-DRA Y-element binding protein YEBP results in a high molecular weight shifted band when nuclear extracts are incubated with oligonucleotides containing the Y- element (Zeleznil-Le et al., 1991). Méthylation interference analysis of a shifted band corresponding to the YEBP/DNA complex reveals that YEBP contacts the Y element within the CCAAT homology (Sherman et al., 1989b). YEBP has yet to be cloned, but it is very likely to be the human homologue of NF-Y (Hooft van Huijsduijnen et al.,
1990) [see below]. The functional role of YEBP has been examined using an in vitro
transcription system. In such a system which had been depleted of YEBP activity by specific competitor oligonucleotides addition of affinity purified YEBP could restore activity. More recent studies have demonstrated that the binding of this factor to the Y element is regulated in a cell cycle specific manner. For example, NIH 3T3 cells synchronised by serum starvation were released and YEBP-binding activity measured at different stages of the cell cycle. The results showed maximum activity in G l. MHC genes are all cell cycle regulated but it remains to be seen if YEBP regulates all isotypes in this manner (Zeleznik-Le et al., 1991).
Amino acid homology among mammalian NF-Y proteins is extensive. Mouse and human clones are 98.6% identical at the DNA level, although the mouse form contains 6 amino acids not found in the human molecule (Hooft van Huijsduijnen et aL, 1990). More recently it has been proposed that NF-Y binding to the Y box renders the X2 and XI elements accessible to their specific protein ligands (Wright e ta l, 1994).
Investigations into the control of HLA class II gene expression in Bare Lymphocyte Syndrome (BLS) patients who fail to express cell surface HLA class II (see below) led to the identification of RF-X (Reith et aL, 1988), an X hox binding transcription factor whose activity was missing in these patients. It was initially defined as severà'gel mobility shift complexes formed with a large oligonucleotide spanning from -140 to -70 base pairs of the HLA-DRA promoter that were present in extracts from normal B cells but absent in extracts from patients with type II or III BLS (see below, “Class II deficient mutants”). A cDNA encoding RF-X was isolated by screening a phage expression library with an X box binding site probe (Reith et at,, 1989; Kohr et a l, 1990) and this confirmed the findings of footprinting and méthylation interference analysis.
The NF-X binding protein was first described as an X box binding protein of the H-2Ea upstream regulatory region (URR), but it is believed to interact with the URR of other MHC class II genes based on its characteristic formation of a doublet/triplet of high molecular weight in gel mobility shift analyses (Benoist and Mathis 1990; Hume and Lee 1990). Although the contact points of NF-X are similar to RF-X,differences exist: N- FX can bind to short oligonucleotides whereas RF-X requires longer oligonucleotides that include W and Y elements for formation of stable complexes. In addition NF-X from BLS lines can bind X whereas RF-X cannot (Reith et aL, 1988).
hXBP-1, an X2 binding protein, was cloned from a phage expression library using an oligo corresponding to the mouse H-2Aa X element plus the X-Y interspace region (Liou
human HLA-D RA X element (although not the -DQA, -DQB or -DRB X2 region). Mutation analysis confirmed that the protein contacts residues within the 3' region of the X element and the 5' end of the interspace region. Other results suggest that hXBP-1 may complex with Fos similar to the Jun protein (Liou et aL, 1990).
An oligonucleotide corresponding to the X2 element of HLA-DRA was used to clone an X2-binding protein from a phage expression library . The resulting gene turned out to be c-Jun. Several experiments suggest that jun in a complex with Fos (called AP-1) binds to the X2 element of HLA-D RA. For instance, footprinting of the H LA -D R A URR with bacterially expressed Jun and Fos leads to the protection of -96 to -90 base pairs which corresponds to the X2 site. Furthermore, a complex formed between Hela nuclear extracts and X2 had identical mobility to one formed with AP-1 site and was blocked or supershifted by anti-Jun/ anti-Fos antibodies respectively. AP-1 distribution and relation to other isotypes remains controversial (Andersson and Peterlin 1990).
MHC class II deficient mutants
The analysis of MHC class II gene regulation has benefited from the existence of mutants lacking class II expression. These mutants are either in vitro transformed cell lines from patients with type II (which lack class II gene expression) or type HI (lack class I and II gene expression) Bare Lymphocyte Syndrome [BLS] or are experimentally derived in vitro
by mutagenesis and selection for MHC class II loss (Hume et al., 1989). Most mutants lack class II MHC protein and RNA expression from all three alleles (HLA-DP, -DQ and -DR), despite the presence of intact HLA genes (Long et at., 1984; Accolla et at., 1985; Lisowska-Grospierre et at., 1985; de Preval et al., 1985). Some mutants retain a low level of expression of a single locus, suggesting some discordant regulation (Levine et al.,
1985; Yang et al., 1988; Hume et al.,1989). The class II MHC genes in the in vivo and
in vitro derived mutant cell lines can be reactivated by somatic cell fusions with MHC class II positive cells (Accolla et al., 1985; Salter et al., 1985; Hume et al., 1989) treatment with phorbol esters, or in some cases through the expression of activated Ras gene products (Hume et a l, 1987). The majority of mutations have been mapped to regions outside the MHC class II locus and for these reasons it is generally believed that the mutations affect trans-acting regulatory factors (Koch et al., 1988; Hume et al., 1989; Doyle et al., 1990). For example, Steimle et al., (1993) isolated a novel gene using an MHC class II negative mutant cell line which functions as a transactivator of MHC class II gene expression and restores expression of all MHC class II isotypes in mutant cells, even correcting the defect in cells from patients with BLS. They named this gene CET A (class II transactivator) and identified a splice variant in BLS which accounts for the mutant class IT phenotype. CUT A has since been shown to be necessary for IFN-y induction of MHC class II genes (Steimle et al., 1994) and for the activation of genes required for antigen presentation such as HLA-DM (Chang and Flavell, 1995).
(Accolla, 1983; Caiman and Peterlin, 1987). One of these, RJ2.2.5 is derived from the B lymphoma Raji, and has been used to demonstrate that fusion with a mouse MHC class 11+ B lymphoma restores class II MHC expression, indicating that the defective trans acting factor (CETA) in RJ2.2.5 is highly conserved between mouse and man (Accolla et ah, 1985). Indeed, stable integration of mouse genomic DNA will restore MHC class II expression (Guardiola et ah, 1986). Through the use of a panel of mouse/human hybrids containing different mouse chromosomes, the locus responsible for restoring MHC class II expression in this case has been mapped to chromosome 16 and called aIr-1 (Accolla et a l, 1986).
Post-transcriptional regulation
Fellous, 1988) and : Osteosarcoma cells (Amaldi et a l, 1989) was demonstrated to be transcriptional, but the fold increase in the levels of mRNA exceeded the increase in the transcription rate, thus implicating a role for post-transcriptional events. Other data suggests that instances where the rate of RNA increase is greater than the rate of transcription can be explained by the build up due to the long half-life of RNA, leading to a large increase in the amount of RNA detected despite only a modest increase in transcription rates (Kern et a l , 1989). De Lerma Barbaro and co-workers (1994) provided evidence for post-transcriptional regulation of HLA class II gene expression. They found that in hybrids constructed by the fusion of human and mouse B cells, HLA- DR and -DP molecules could be detected at the cell surface, but the cells were -DQ negative. However, investigation of mRNA revealed that normal levels of HLA-DQa and -DQp transcripts were present. These findings led the authors to conclude that the lack of cell surface expression of HLA-DQ must be caused by an isotype-specific post- transcriptional mechanism.
Evidence for HLA-DQ specific transcription events
The MHC class II region represents a family of linked genes (HLA-DQ, -DP and -DR) and it is generally accepted that the expression of these various isotypes is co-ordinately regulated. There are several reasons for this assumption e.g., both B cells and non-immune cells when stimulated with IFN-y show a co-ordinately timed increase in the level of mRNA of all MHC class II genes (Fertsch et a l , 1987; Paulnock-King et a l , 1985). Furthermore, MHC class II mutant regulatory cell lines exhibit the combined loss of expression of all MHC class II genes (Accolla, 1983; Long et a l , 1984). These observations strongly suggest that a trans-acting factor regulates the function of many, or all, MHC class II genes.