Copyright1998 by the Genetics Society of America
Detection of Deleterious Genotypes in Multigenerational Studies.
II. Theoretical and Experimental Dynamics with Selfing and Selection
Marjorie A. Asmussen, Laura U. Gilliland and Richard B. Meagher
Department of Genetics, Life Sciences Building, University of Georgia, Athens, Georgia 30602-7223
Manuscript received February 2, 1998 Accepted for publication March 10, 1998
ABSTRACT
A mathematical model was developed to help interpret genotype and allele frequency dynamics in selfing populations, with or without apomixis. Our analysis provided explicit time-dependent solutions for the frequencies at diallelic loci in diploid populations under any combination of fertility, viability, and gametic selection through meiotic drive. With no outcrossing, allelic variation is always maintained under gametic selection alone, but with any fertility or viability differences, variation will ordinarily be maintained if and only if the net fitness (fertility3 viability) of heterozygotes exceeds that of both homozygotes by a substantial margin. Under pure selfing and Mendelian segregation, heterozygotes must have a twofold fitness advantage; the level of overdominance necessary to preserve genetic diversity declines with apomixis, and increases with segregation distortion if this occurs equally and independently in male and female gametes. A case study was made of the Arabidopsis act2-1 actin mutant over multiple generations initiated from a heterozygous plant. The observed genotypic frequency dynamics were consistent with those pre-dicted by our model for a deleterious, incompletely recessive mutant in either fertility or viability. The theoretical framework developed here should be very useful in dissecting the form(s) and strength of selection on diploid genotypes in populations with negligible levels of outcrossing.
M
UTATIONS that are not lethal or do not produce difficult to predict the equilibrium frequencies reached or even whether a genetic polymorphism will be main-an obvious morphological phenotype are oftentained or lost in the population. used to categorize genes or gene functions as
nonessen-The case of a purely selfing population, however, is tial or redundant. However, this conclusion can seldom
fully analyzable, which provides us with a valuable theo-be justified without evolutionary data on allele
frequen-retical framework from which to infer the form(s) and cies across multiple generations (Gillilandet al. 1998).
strength of selection operating in such populations. This Arabidopsis is an ideal model organism for
multigenera-is because the expected genotypic and allelic frequencies tional population studies, both in the field (Mauricio
at a diallelic locus can be readily predicted through time 1998) and laboratory (Gillilandet al. 1998), because
in a selfing population under selection.Karlin(1969) of its small size, short life cycle, and small genome (
Mey-outlined a sophisticated dynamical analysis of such sys-erowitz 1989, 1994). Moreover, the negligible rate of
tems based on a nontraditional application of linear outcrossing (AbbottandGames1989), even when
in-algebra. This approach was akin to that used to determine florescences are in close physical contact (Snape and
the equilibria with all alleles present under the classical Lawrence 1971), allows a purely selfing model to be
selection model with constant viability selection on a used to analyze the observed frequencies of genotypes
multiallelic locus (Mandel 1959). Karlin’s method in this organism. The ability to apply this approximation
hinged on temporarily ignoring the normalizing factor is a tremendous advantage because a mixed-mating
se-that makes the new genotypic frequencies sum to one lection model for diploid populations, incorporating
and treating the simplified genotypic recursions as three both selfing and random outcrossing, is far more
com-independent linear equations (whereas technically plicated to analyze (Workman and Jain 1966; Weir
there are only two independent equations). He then 1970;KimuraandOhta1971;OverathandAsmussen
specified the dynamical solutions through time (but not 1998). In general, if there is outcrossing present, explicit
their explicit formulas) in terms of a triple product of time-dependent solutions are not possible for the
geno-3 3 3 matrices, whose entries were functions of the typic frequencies, even if the genotypes differ only in
eigenvalues and eigenvectors of the resulting three-vari-viability. The equilibrium analysis is also very complex
able linear transformation. A further complication of in mixed-mating systems under selection because it is
this approach is that the solutions for certain combina-tions of parameter values require computing the Jordan canonical decomposition of the coefficient matrix. Corresponding author: Marjorie A. Asmussen, Department of
Genet-Here we provide a much simpler derivation than that
ics, Life Sciences Building, University of Georgia, Athens, GA
30602-7223. E-mail: [email protected] proposed by Karlin (1969), which readily generates
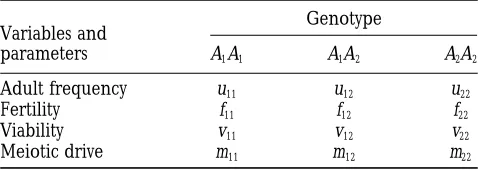
TABLE 1 are transmitted independently and with the same fre-quency of A1(m) and A2(12m) alleles, meiotic drive
Genotypic frequencies and selection parameters
can be described by the single parameter m, where
m115m2, m1252m(12 m), m225 (1 2 m)2, and 0#
Genotype
Variables and m# 1. Under Mendelian segregation, m51⁄
2, yielding
parameters A1A1 A1A2 A2A2 the familiar proportions m
115m2251⁄4 and m125 1⁄2. Derivation of purely selfing model: The changes in
Adult frequency u11 u12 u22
Fertility f11 f12 f22 adult genotypic frequencies from one generation to the
Viability v11 v12 v22 next in a purely selfing population are readily
deter-Meiotic drive m11 m12 m22 mined by working through a complete generation cycle.
Although we do not monitor the population size in this model, the frequency dynamics are most easily derived by considering the number of each genotype at the the explicit time-dependent solutions for the genotypic
successive life stages. This factor will be eliminated in the and allelic frequencies under all possible parameter
val-final step, where we calculate the new adult frequencies. ues. Our results provide a theoretical framework that
Letting N represent the total number of reproducing fully dissects the evolutionary dynamics under any
com-adults at the start of the current generation, the number bination of fertility, viability, and gametic selection on
of A1A1 zygotes is then, for instance, f11(u11N ) 1 a single diallelic locus in a purely selfing population,
m11f12(u12N )5( f11u111m11f12u12)N, where f11(u11N) is the with or without the asexual production of seeds by
apo-number of A1A1 zygotes produced by the selfing of mixis. This framework can also be applied with
reason-the u11N individuals who are A1A1, and m11f12(u12N ) is able accuracy to partially random mating populations
the number produced by the selfing of the u12N individu-in which the outcrossindividu-ing rate is negligible. The utility
als who are A1A2. Note that all progeny from A1A1 homo-of this approach is illustrated through application to
zygotes are like homozygotes, whereas a fraction m11 the multigenerational data on the act2-1 actin mutant
of the progeny from heterozygotes will be A1A1. The in Arabidopsis, presented in a companion manuscript
generation cycle concludes with viability selection, after (Gillilandet al. 1998).
which the number of new A1A1 adults is v11(f11u11 1
m11f12u12)N. Analogous formulas apply to A2A2 homozy-gotes. In contrast, A1A2 individuals can be produced POPULATION DYNAMICS WITH
only from other heterozygotes under pure selfing. The
SELECTION IN SELFING POPULATIONS
number of A1A2zygotes is thus simply m12f12(u12N ), and The expected genotypic and allelic frequencies at a the number that survive to become A1A2 adults is locus with two alleles (A1 and A2) can be predicted v12m12f12(u12N ).
through time under selfing, apomixis, and selection, Normalizing relative to the total number of new adults using a simple deterministic model. We first assume a (wN), we then find that the three genotypic frequencies purely selfing diploid population with discrete, nonover- in adults change from one generation to the next ac-lapping generations with no mutation or gene flow, cording to the following recursions:
which is large enough to preclude the effects of random
genetic drift. The genotypes are subject to constant fer- u9 115
( f11v11)u111( f12m11v11)u12
w (1)
tility selection and/or viability selection and possibly gametic selection via meiotic drive (deviation from
u9125
(m12f12v12)u12
w (2)
50:50 Mendelian ratios in gametes produced by hetero-zygotes). The frequencies of the three genotypes in adults (u) and the selection parameters ( f, v, m) are
u9225
( f22v22)u221( f12m22v22)u12
w , (3)
defined in Table 1, where subscripts ij refer to the alleles carried by the individual. The fertility parameters fij
de-where a prime (9) denotes a value in the next genera-note the average number of offspring produced when
tion, and the normalization factor is an AiAjindividual self fertilizes, while the viability
param-eters vij denote the average fraction of newly formed w5( f
11v11)u111f12(m11v111m12v121m22v22)u12
AiAjzygotes that survive to reproduce. 1( f
22v22)u22. (4) Meiotic drive is incorporated through the parameters
mij, which represent the average fraction of progeny that This factor (w) could be viewed as a measure of the
mean fitness in the population, although this interpreta-are AiAjwhen an A1A2individual selfs. In general, these
values can be any three nonnegative numbers that sum tion is complicated by the fact that the second term, weighted by the frequency of heterozygotes (u12), in-to 1 (m11 1m121 m225 1) to allow for the possibility
of differential or nonindependent transmission of the volves parameter values for all three genotypes. The corresponding iterative formula for the frequency p of alleles to male and female gametes. If the male and
p9 5u91111⁄2u912 values in the population whose interpretations are given below.
The explicit analytical formulas for the genotypic fre-5( f11v11)u111 f12(m11v111 1⁄2m12v12)u12
w . (5) quencies after any number of generations t of selfing
and selection are now obtained simply by substituting Time-dependent solutions: Here we present a new
the solutions for x(t)
1 and x(t)2 from (11–14) into the rela-and complete dynamical analysis of this model (Karlin
tions in (6–8). The corresponding time-dependent solu-1969), which quickly generates the explicit
time-depen-tion for the frequency of the A1allele is dent solutions for the genotypic and allelic frequencies
under all possible parameter values. Our approach
capi-p(t)5 u(t)
111 1⁄2u(t)12 talizes on the fact that the system is two dimensional,
along with a change to the readily analyzed variables,
5 x(t)1 11⁄2 11x(t)
1 1x(t)2
for t50, 1, .... (15)
x15u11/u12and x25u22/u12, which are the ratios of the homozygote to heterozygote frequencies in the
popula-Dynamical and equilibrium behavior:The form of these tion. The quantities x1and x2represent two independent
solutions immediately reveals that the dynamical and variables that fully describe the three-genotype system
limiting behavior under this model depends on the via the relations
relative magnitude of the geometric terms a1 and a2 [defined in (12)], and whether their values are above,
u(t)11 5
x1(t) 11 x(t)1 1x(t)2
(6)
below, or equal to 1. A comprehensive analysis of the dynamics under all possible numerical orderings of a1,
a2, and 1 shows that there are precisely four types of
u(t)12 5
1 11 x(t)1 1x(t)2
(7)
evolutionary outcomes for the equilibrium genotypic frequencies in the population. These types of outcomes
u(t)12 5
x2(t) 11 x(t)1 1x(t)2
, (8) are given in Table 2 in terms of the values of a 1 and
a2, together with the corresponding conditions on the which hold in every generation t, where for any variable selection parameters. This classification both completes
z, z(t)represents its value in generation t. and simplifies that presented byKarlin(1969). The recursions for the two new variables, x1and x2, are Turning to the biological interpretation of these find-simple, independent, linear difference equations with ings, we see that the population dynamics are governed constant coefficients: by the relative magnitudes of composite fitness values for each of the three genotypes (Table 2, column 3). These delimiting values correspond to the relative rate
x915
1
f11v11
m12f12v12
2
x1 1
m11v11
m12v12
(9)
at which individuals survive and produce offspring with their own genotype. For homozygotes, which only bear
x925
1
f22v22
m12f12v12
2
x2 1
m22v22
m12v12
. (10) progeny like themselves, these values are simply their
net fitnesses ( f11v11,f22v22), corresponding to the product These both have the same general form, x9i 5 aixi 1 of their average number of offspring and the probability
bi. Their dynamical solutions in every generation t50, that they survive to reproduce. The value for
heterozy-1, ..., are immediately found to be gotes (m12f12v12), however, is discounted by the factor m12 because heterozygotes do not ordinarily breed true. In the special case of Mendelian segregation, for instance,
x(t)
i 5
5
(ai)t(x(0)i 2x*i )1 x*i if ai ?1
x(0)
i 1 tbi if ai51
for i5 1, 2,
the composite fitness value for heterozygotes is half their (11) net fitness (1⁄
2f12v12).
The results summarized in Table 2 show that under where x(0)
i 5 u(0)ii /u(0)12 is the initial value of xi, and the this model, a purely selfing population will converge
remaining terms,
to a polymorphic equilibrium with all three genotypes present (outcome 1) if and only if the discounted net
ai5
fiivii
m12f12v12
(12) fitness of heterozygotes exceeds the net fitness of both homozygotes (m12f12v12.f11v11,f22v22). These selective con-ditions are equivalent to the requirement that the net
fit-x *i 5
f12miivii
m12f12v122 fiivii
(13)
ness of heterozygotes (f12v12) exceed those of the homozy-gotes ( fiivii) by the factor 1/m12. With Mendelian
segre-and gation, this requires double overdominance ( f
12v12 . 2f11v11,2f22v22). The minimum level of overdominance
nec-bi5
miivii
m12v12
TABLE 2
Classification of equilibrium genotypic frequencies under fertility, viability, and gametic selection in a selfing population
Genotype Conditions
Outcome on a1and a2 Selection conditions A1A1 A1A2 A2A2
1 a1, a2,1 f11v11, f22v22,m12f12v12 uˆ11.0 uˆ12.0 uˆ22.0 2 a15a2$1 f11v115f22v22$m12f12v12 uˆ11.0 0 uˆ22.0
3 a2,a1$1 f22v22,f11v11$m12f12v12 1 0 0
4 a1,a2$1 f11v11,f22v22$m12f12v12 0 0 1
independent in male and female gametes [m12 5 2m unique equilibrium values (12 m)].
Under the conditions producing outcome 1, the pop- uˆii5
bi
b1 1b2
5 miivii
m11v111 m22v22
for i 51, 2, (20) ulation will reach the unique equilibrium
where b1and b2are defined in (14). In this case, the final
uˆ115
x*1
11x*1 1 x *2
frequencies of the two homozygotes are in proportion to the product of their viabilities (vii) and the rate at which
they are produced by heterozygotes (mii).
uˆ125
1
11x*1 1 x *2 Alternatively, if the common net fitness of the homo-zygotes is higher than the discounted heterozygote value ( f11v115 f22v22. m12f12v12), as is true in the absence of
uˆ225
x*2
11x*1 1 x *2
, (16)
selection, there is an infinite number of the second type of polymorphic equilibria. This is because the final where x*1 and x*2, defined in (13), are in this case the
homozygote frequencies then depend on the initial ge-final, limiting values of the variables x1 and x2. After
notypic frequencies in the population, with substituting in their values, we obtain the exact
equilib-rium formulas for the genotypic frequencies
uˆii5
x(0)i 2x*i
x(0)1 1x(0)2 2x*1 2x*2
for i51, 2. (21)
uˆii5
f12miivii(m12f12v122 fjjvjj)
D for i? j51, 2 (17) After substituting the values for x*1 and x*2 from (13) and simplifying, we obtain the exact expressions for and
these equilibria in terms of the selection parameters and initial genotypic frequencies:
uˆ125
(m12f12v122 f11v11)(m12f12v122 f22v22)
D , (18)
uˆii5
where the common denominator is ( f
11v112m12f12v12)u(0)ii 1f12miiviiu(0)12
( f11v112 m12f12v12)(u(0)11 1u22(0))1 f12(m11v111m22v22)u(0)12
D5 f12m11v11(m12f12v122 f22v22)
(22) 1(m12f12v122f11v11)(m12f12v122f22v22)
for i5 1, 2. 1f12m22v22(m12f12v122f11v11) . (19)
Two final general points should be made about the two types of polymorphic equilibria reached in this sys-A second type of polymorphic equilibrium is also
pos-sible for purely selfing populations, with only the two tem (Table 2, outcomes 1 and 2). First, although the genotypes present at equilibrium depend only on the homozygous genotypes present. In this model,
heterozy-gotes will be eliminated monotonically, and the popula- relative values of the composite fitnesses of the three genotypes, the formulas in (17–22) demonstrate that tion will eventually split into two pure homozygous lines
if and only if the net fitnesses of the two homozygotes the exact final frequencies at a polymorphic equilibrium depend on the individual selection components because are equal and are at least the discounted value for
het-erozygotes (Table 2, outcome 2). A given population has they involve additional terms of the form f12miiviiand
miivii. Second, barring very special relationships among
either a single or an infinite number of such equilibria,
depending on the relative magnitudes of the composite the fitness parameters, the first type of polymorphic equilibrium will ordinarily be the only way to maintain fitnesses of the three genotypes. When the common net
fitness of the homozygotes exactly equals the discounted allelic variation if the genotypes differ in fertility or viability. The final two possible equilibrium states (Table value for heterozygotes ( f11v115 f22v225 m12f12v12), the
allele and loss of the other. The population will become Equation 22 shows that with pure selfing, the fre-quency of AiAihomozygotes will converge to
fixed for A1if the net fitness of A1A1individuals exceeds that of A2A2 individuals and is at least as high as the
discounted value for heterozygotes ( f22v22 , f11v11 $ uˆii5 u(0)ii 1
1
mii
m111m22
2
u(0)
12 . (25)
m12f12v12. Analogous conditions lead to the fixation of A2.
The four outcomes in Table 2 apply to the most bio- The final frequency of each homozygote equals its initial logically relevant case in which heterozygotes are ini- frequency (u(0)
ii ) plus a proportion of the initial
fre-tially present (u(0)12 .0) and further adult heterozygotes quency of heterozygotes (u(0)12) corresponding to the rela-can be produced (m12f12v12.0). If this is not the case, the tive rate at which that homozygous genotype is produced population remains as two homozygous lines, with their by heterozygotes (mii). The equilibrium frequency of allele relative frequencies in every generation t$1 given by A1, pˆ, will exceed its initial value (p(0)5 u(0)
11 1 1⁄2u(0)12) if and only if A1is preferentially transmitted by
heterozy-u(t)
11
u(t)
22
5
1
f11v11f22v22
2
t
1
u(0) 11u(0)
22
2
. (23) gotes (m11.m22). The A1allele will predominate at equilib-rium (pˆ . 1⁄
2) if and only if m11(1 2 2u(0)22) . m22(12 The frequencies of the A1A1genotype and the A1allele 2u(0)11), which compares unexpected cross-products in-in adults in-in any generation t 5 0, 1,..., then have the volving the initial frequencies of each homozygote and common value the rate at which the other homozygote is produced by heterozygotes. The dependence upon initial conditions under meiotic drive is illustrated in Figure 1, B and C.
u(t)11 5p(t)5
atu(0) 11 12u(0)11 1 atu(0)11
, where a5f11v11
f22v22 .
Fertility selection alone (m115 m225 1⁄4, m1251⁄2; vij ;
(24) 1): If a locus is subject only to fertility selection, with
Mendelian segregation and no viability differences Analysis of this dynamical solution (24) shows that in
among the genotypes, the geometric terms in (12) are the absence of heterozygotes, the frequency of allele A1
a15 2f11/f12and a25 2f22/f12. All four possible equilib-increases monotonically to 1 if A1A1 individuals have a
rium outcomes (Table 2) can occur in this case, de-higher net fitness than A2A2individuals (f11v11.f22v22),
pending upon the relative magnitudes of the fertility of decreases monotonically to 0 if A2A2 individuals have
heterozygotes (f12) and twice the fertility of the two the higher net fitness ( f11v11,f22v22), and remains at its
homozygotes (2f11, 2f22). The population will reach a initial value if both homozygotes have the same net
fully polymorphic equilibrium with all three genotypes fitness ( f11v115f22v22), as expected for selection on two
present (Figure 1D, outcome 1) if and only if heterozy-autonomous lines.
gotes produce over twice as many offspring as each Individual selection components:The preceding
analy-homozygote (f12.2f11, 2f22). Using (17–19), the limiting sis fully delimits the dynamical and limiting behavior in
genotypic frequencies are then purely selfing populations experiencing any
combina-tion of fertility, viability, and gametic seleccombina-tion. Because
of their distinctive features, it is also informative to ex- uˆii5
f12( f1222fjj)
2(f1222f11)( f122 f22)1 2(f1222f22)(f122f11) amine the evolutionary consequences of each of these
three selection components individually. For simplicity, for i?j51, 2 (26) we focus here on the most interesting biological case,
and in which adult heterozygotes are initially present, can
continue to be produced, and bear both heterozygous
uˆ125
( f1222f11)(f1222f22)
( f1222f11)(f122f22)1 ( f122 2f22)(f122f11) . and homozygous progeny (0,m12,1). Sample
trajec-tories for the genotypic and allelic frequencies under (27)
each form of selection are provided in Figure 1, with
These polymorphic equilibrium frequencies depend on Figure 1A showing the baseline dynamics in the absence
the fertility of heterozygotes ( f12) and the difference of selection.
between the fertilities of heterozygotes and
homozy-Gametic selection alone ( fij;vij;1): If a diallelic locus
gotes ( f122 fii), as well as the difference between the
is subject only to meiotic drive, with equal fertilities and
fertilities of heterozygotes and twice that of homozy-viabilities for the three genotypes, then the geometric
gotes ( f1222fii), but not directly on the fertility of the
terms in (12) are a15a251/m12.1. This ensures that
homozygotes. the population will always converge to the second type
Alternatively, a polymorphism will be maintained via of polymorphic equilibrium (Table 2, outcome 2)
con-a split into the two homozygous lines (outcome 2) if sisting of the two true breeding homozygous lines.
Con-the two homozygotes have equal fertilities that are at sequently, allelic variation will always be maintained
un-least half that of heterozygotes (f11 5 f22 $ 1⁄2f12). der meiotic drive in a purely selfing population, whereas
If f115f2251⁄2f12, then (20) shows that the final frequency a random mating population will usually become fixed
of both homozygotes will always be1⁄
2, whatever the exact for the preferentially transmitted allele (Hartl and
Figure1.—The impact of gametic, fertility, and viability selection on genotype and allele frequencies under our purely selfing model. Genotypic frequencies u(t)
(31) 1E), (22) shows that the final frequencies of the two
homozygous genotypes depend on both the fertilities
differ from those in (28) for fertility selection in that and initial frequencies of each genotype, with
the heterozygote fitness is replaced by the homozygote value in the terms involving the initial frequency of
uˆii5
2(2f112 f12)u(0)ii 1f12u(0)12 2(2f112f12)(u(0)11 1u(0)22)12f12u(0)12
for i51, 2.
heterozygotes (u(0)
12). The conditions for fixation are completely analogous under the two forms of selection. (28)
Under viability selection alone, the population will be-The population will become fixed for allele Ai(Figure come fixed for allele A
i (Figure 1I, outcome 3 or 4) if
1F, outcome 3 or 4) if and only if AiAiindividuals pro- and only if A
iAi individuals survive to reproduce more
duce more progeny than AjAjindividuals and produce often than A
jAjindividuals, and survive at least half as
at least half as many progeny as heterozygotes ( fjj , often as heterozygotes (v
jj, vii$1⁄2v12).
fii$1⁄2f12). Mixed selfing and apomixis model:A direct extension
Viability selection alone (m115 m225 1⁄4, m125 1⁄2; fij ; of this approach provides a full dynamical and
equilib-1): The behavior under viability selection alone, with rium analysis for populations that reproduce by a combi-Mendelian segregation and no fertility differences nation of selfing and apomixis (the asexual production among the genotypes, is similar to that under fertility of zygotes). Formally, we assume each individual selfs with selection alone. The geometric terms in (12) have the probability s and reproduces apomictically with probability analogous forms a1 5 2v11/v12 and a2 5 2v22/v12, and 12s. The only other difference from the original model thus all four equilibrium outcomes (Table 2) are again is that separate fertilities are allowed for the two forms of possible, with the delimiting values now the relative reproduction, with f
ijrepresenting the average number of
magnitudes of the viability of heterozygotes (v12) and progeny produced by selfing, as before, and F
ij
repre-twice the viability of the two homozygotes (2v11, 2v22). senting the number of progeny from apomixis. Paralleling fertility selection, all three genotypes will be Working through a complete generation cycle shows retained at equilibrium (Figure 1G, outcome 1) if and that the genotypic recursions under this mixed, selfing-only if heterozygotes survive to reproduce at over twice apomixis model are
the rate of each homozygote (v12.2v11, 2v22). The final values,
u9115
[sf111 (12 s)F11]v11u111(sf12m11v11)u12
w (32)
uˆii5
vii(v122 2vjj)
(v122 2v11)(v122v22)1(v1222v22)(v122v11) u9 125
[sm12f121(1 2s)F12]v12u12
w (33)
for i? j51, 2 (29)
and u9225
[sf221 (12 s)F22]v22u221(sf12m22v22)u12
w , (34)
uˆ125
2(v1222v11)(v1222v22)
(v122 2v11)(v122 v22)1(v1222v22)(v122v11)
, where the normalizing factor is now
(30) w5[sf111 (12s)F11]v11u11
however, are not equivalent to those in (26) and (27) 1[sf12(m11v111m12v121m22v22) for fertility selection. One difference is that with viability
1(1 2s)F12v12]u12 selection, the equilibrium genotypic frequencies
de-pend directly on the homozygote viabilities and not the 1[sf221(1 2s)F22]v22u22. (35) heterozygote viabilities, which is the reverse of the result
The time-dependent solutions for the transformed vari-for fertility selection. Most importantly, comparing (27)
ables, x1 5 u11/u12and x2 5 u22/u12, are equivalent to and (30) reveals that viability selection maintains twice
those in (11) with the constant factors now as many heterozygotes at equilibrium as does a
compara-ble level of fertility selection (fij5 vij).
Further differences are found for the second type of ai 5
[sfii1(12 s)Fii]vii
[sm12f121 (12s)F12]v12
(36) polymorphic equilibrium (outcome 2), consisting of the
two homozygous genotypes. The final frequencies of
x*i 5
sf12miivii
[sm12f121(12 s)F12]v122[sfii1(12 s)Fii]vii
the two homozygotes are again uˆii51⁄2if the viabilities of
the two homozygotes are equal and half that of heterozy- (37) gotes (v115v225 1⁄2v12); however, the final frequencies
and of the two homozygotes when v115 v22.1⁄2v12(Figure
1H),
bi5
sf12miivii
[sm12f121(12 s)F12]v12
5 sf12mii
sfii1 (12s)Fii
.
uˆii5
2(2v112 v12)u(0)ii 1 v11u(0)12 2(2v112 v12)(u(0)11 1u(0)22)1 2v11u(0)12
for i51, 2,
Note that the second formula for biin (38) is a simplifi- without selection. The three actin gene family members
belong to different, ancient, and conserved actin sub-cation that applies specifically to the case in (11) where
ai51. Substituting these new constant factors into (11) classes, and they are all strongly expressed in a distinct
temporal and spatial pattern (McDowell et al. 1996). and the resulting solutions for x(t)1 and x(t)2 into (6–8)
and (15) yields the dynamics of the genotypic and allele Although homozygous mutant plants (A2A2) in each of the three actin genes appeared morphologically normal frequencies under selection with any combination of
selfing and apomixis. and robust as adults, and appeared to have a normal seed set relative to the wild-type (A1A1), it is unlikely that such This new model has the same four possible
evolution-ary outcomes as the original, based on the relative mag- highly conserved genes could be fully redundant. The detailed experimental analysis of the act2-1 ge-nitudes of a1, a2, and 1 shown in Table 2. Biologically, the
actual outcome (and the rate at which it is attained) again notype frequencies (Gilliland et al. 1998) provides a valuable case study for the application of the theoretical depends on composite fitness values for the three
geno-types, with the homozygote values in the third column framework developed herein for detecting deleterious ge-notypes. Starting with a single heterozygote in the F0 gener-of Table 2 replaced by [sf111(12s)F11]v11and [sf221
(12 s)F22]v22, and the heterozygote value replaced by ation, these experiments followed the genotype and allele frequencies in large selfing populations (ca. 100 plants [sm12f121(12s)F12]v12. These new composite fitnesses
are a weighted average of those under selfing and apo- each) through the F3 generation. Because there were highly significant deviations in the F2and F3generations mixis. The selfing component is as before, the net fitness
under self-fertilization ( fiivii) for homozygotes and the from the genotypic frequencies expected under selective
neutrality, the null hypothesis of no selection is rejected discounted value (m12f12v12) for heterozygotes, while that
for apomixis is simply the net fitness of each genotype for the data set as a whole. Inasmuch as the F1progeny of the original heterozygote were consistent with Mendelian when it reproduces apomictically (Fijvij), since each
indi-vidual then breeds true. segregation ratios (1:2:1), meiotic drive does not appear to be the major factor. In the initial analysis here, we con-Inspection of the biological conditions associated with
each of the four evolutionary outcomes reveals two note- sequently focus on fertility and viability selection as the possible causes of the reduced frequency of the act2-1 worthy features of mixed selfing and apomixis. First,
under meiotic drive alone, allelic polymorphisms are allele under our model.
After exploring various combinations of fertility always maintained via a split into the two homozygous
lines because then a15 a25 1/[12 s(12 m12)].1. parameters, we found that the multigenerational data on the act2-1 mutant are consistent with fertility selec-In fact, the equilibrium frequencies are independent
of the relative proportions of apomixis and selfing and tion alone, where the fertility of the A1A2heterozygotes are only slightly reduced ( f125 0.8), while that of the always equal those in (25) for purely selfing populations.
The second point arises from the fact that if there are A2A2homozygous mutant is half ( f225 0.5) that of the wild type ( f11 5 1), as shown in Figure 2A. The chi-any fertility or viability differences, genetic variation will
ordinarily be maintained if and only if [sm12f12 1 (1 2 square values for the genotypic frequencies in the F1, F2, and F3 generations are all less than 2, showing that
s)F12]v12.[sf111(12s)F11]v11, [sf221(12s)F22]v22. The
presence of apomixis thus serves, as expected, to facilitate these data do not deviate significantly from the pre-dicted values (P 50.38–0.75 with 2 d.f.).
the maintenance of genetic variation, because the
apo-mictic component is satisfied by simple overdominance Viability selection alone gives an even better fit to the experimental results (Figure 2B) when the viability of (F12v12.F11v11, F22v22) as opposed to the
“super-overdomi-nance” condition for the selfing component. The final the A1A2 heterozygotes is slightly reduced (v1250.87) and the viability of the A2A2homozygous mutant is only limiting values for the polymorphic equilibria under
selfing and apomixis can be obtained by substituting the a bit lower (v225 0.7) relative to that of the wild type (v1151). The chi-square values for the three genotypes values from (37) and (38) into (16), (21), and the first
relation in terms of b1and b2in (20). are all below 1.2 (P50.57–0.90). In theory, the simplest interpretation of this experimental data would be that only the homozygous mutants have a reduced fitness.
ARABIDOPSIS ACTIN MUTANTS: A CASE STUDY
However, when a simple recessive model was explored, where only the A2A2genotype had reduced fertility (not This mathematical framework provides a valuable tool
for dissecting the selective forces at work in large diploid shown) or reduced viability (Figure 2C), the model did not fit the experimental results as well as models with populations with negligible rates of outcrossing. An
imme-diate application is furnished by the companion article directional selection and partial dominance (P50.13– 0.94).
(Gillilandet al. 1998), in which we demonstrated that
mutant alleles for three distinct plant actin genes in Arabi- Two practical points should be made about these numerical calculations. First, they are based on an infor-dopsis (act2-1, act4-1, and act7-1) were significantly reduced
in these experiments can be well explained by our sim-ple selection model. Formal estimates of the selection components are obtainable by collecting and analyzing data from further generations via a maximum likelihood estimation procedure based on the model (see discus-sion). The second practical note is that the projected genotypic frequencies and the comparisons with empiri-cal data presented here were all empiri-calculated and plotted using Delta Graph, version 4.0.1, which runs on a Macin-tosh computer (Deltapoint, Inc., Monterey, CA). A copy of the program is available through our web site (www.genetics.uga.edu) by requesting software for de-tection of deleterious genotypes.
DISCUSSION
A formal dissection of selection components is greatly facilitated in diploid populations that lack appreciable outcrossing, such as those for the model organism Arabi-dopsis. This simpler genetic structure allows the develop-ment of precise analytic formulas for predicting the evolu-tionary consequences of selection in diploid, selfing populations, with or without apomixis. By focusing on the readily analyzable dynamics of the ratio of homozy-gote to heterozyhomozy-gote frequencies, we easily derived ex-plicit time-dependent solutions for the genotype and allele frequencies at diallelic loci under any combina-tion of fertility, viability, and gametic seleccombina-tion through meiotic drive. In the absence of outcrossing, such selec-tion may maintain all three genotypes, only the two homo-zygotes, or only a single homozygous line, depending on the relative magnitudes of composite fitness values for the three genotypes. The delimiting values are a weighted average of the net genotypic fitnesses (fertility3 viabil-ity) under selfing and apomixis, with the selfing compo-nent for heterozygotes discounted by the frequency with which heterozygotes breed true when they self fertilize. Interestingly, although fertilities and viabilities play an equivalent role in determining which genotypes are re-tained or lost, the two types of selection yield different frequencies at polymorphic equilibria; viability selection alone, for instance, maintains twice the frequency of heterozygotes found under comparable levels of fertility selection in selfing populations.
The selective conditions that retain genetic diversity in selfing populations are very different from those in outcrossing populations. With or without apomixis,
Figure2.—Fertility or viability selection alone can account script (Gillilandet al. 1998). Selection parameters were cho-for the decline of the act2-1 allele from Arabidopsis popula- sen which most closely approximated the observed genotypic tions. The genotypic frequencies u(t)11, u(t)12, and u(t)22are plotted frequencies, where ACT25A1and act2-15A2. Unless indi-as a function of generation (t) for a large selfing population cated otherwise, fitness parameters were fij5 vij51, m115 for six generations, with the F0generation being a single het- 0.25, m125 0.5, and m22 50.25. (A) Fertility selection with erozygote (u(0)
selfing will always preserve allelic variation under mei- the actin mutants. The power to detect and estimate such selection, however, will depend critically on having otic drive alone, whereas any random mating will usually
lead to fixation for the preferentially transmitted allele. an adequate sample size and multigenerational data. In our preliminary analysis of the act2-1 data, the best-On the other hand, selfing hinders the maintenance of
genetic variation if there are any fertility or viability fitting selection parameters for the observed genotypic frequencies for ACT2 were obtained by an informal differences; in such cases, a genetic polymorphism will
ordinarily be preserved under selfing if and only if the exploration of the parameter space. This ad hoc approach nonetheless provided an impressive fit to the three gen-net fitness of heterozygotes is substantially higher than
those of both homozygotes, as opposed to the simple erations of data available to date, assuming either fertil-ity or viabilfertil-ity selection alone. It is difficult to tell, how-overdominance in fitness needed in fully random
mat-ing populations. Selfmat-ing also makes it more difficult to ever, whether this good fit will be maintained in subsequent generations (Figure 2) or whether the act2-1 preserve allelic variation when combined with either
apomixis or outcrossing, or both (OverathandAsmus- mutant is subject to more than one selection compo-nent. Before finalizing our conclusions on the form(s) sen1998). Under complete selfing and Mendelian
seg-regation ratios, permanent genetic diversity usually re- and strength of selection acting in this system, it will be important to continue these experimental Arabidopsis quires a twofold fitness advantage for heterozygotes,
with the required level of overdominance ameliorated populations for several additional generations and then analyze the complete data set by using formal statistical by apomixis and magnified by meiotic drive.
The results here apply directly to fairly large, isolated estimation procedures (Weir1996).
To directly estimate all three components of selection populations with no outcrossing, but they can also
pro-vide valuable insight into the behavior when there is a (fertility, viability, and gametic) in our model, we would ideally have population data from all three life stages negligible rate of random mating. The accuracy of this
approximation depends on the number of generations (adults, new zygotes, and gametes). In practice, how-ever, most experimental population genetic studies ex-followed, since any outcrossing will ordinarily preclude
the second equilibrium outcome above, in which the amining diallelic loci have only adult data with 2 d.f. for each generation, as presented in Gillilandet al. population ultimately splits into the two homozygous
lines. Practically speaking, however, the final frequency (1998) for the mutant actin alleles in Arabidopsis. With this limitation, we can estimate two of the fitness param-of heterozygotes with insignificant rates param-of outcrossing
should be so low that they are effectively lost from such eters (Table 1) and thus one of the selection compo-nents, based on data from any two generations (one of populations. In addition, over the relatively small
num-ber of generations followed in most empirical applica- which may be the initial population). For the fertility and viability components, this requires normalizing the tions, the expected trajectories for the genotypic
fre-quencies should be essentially unaffected by a low rate three fertilities (or viabilities) relative to a value of 1 for A1A1, for example, which is legitimate because the of outcrossing.
The practical utility of this theoretical framework is genotypic recursions in (1–4) are unaffected if each fertility (or viability) parameter is altered by the same affirmed by a case study of the act2-1 actin mutant in
highly selfing, experimental populations of Arabidopsis. constant factor.
A direct extension of this reasoning indicates that, Several critical conclusions can be drawn from this
sim-ple application of our generalized selfing model. First, in principle, all three selection components could be estimated from adult data, provided that independent the data are well fit by either fertility or viability selection
alone, with directional selection against an incompletely samples are available from three or more generations beyond the F0(each with two degrees of freedom). By dominant mutant (a reduced fitness for A1A2 and a
greater reduced fitness for A2A2 relative to A1A1 wild adapting established methods based on the fit to equilib-rium frequencies (e.g., Asmussen et al. 1989; Goodis-type). Second, under the two best-fitting sets of selection
parameters (Figure 2, A and B), the act2-1 allele should man and Asmussen 1997), our time-dependent solu-tions under selection can be used to obtain formal be effectively lost (frequency,0.1%) within 20
genera-tions (data not shown). For Arabidopsis in the field, maximum likelihood estimates for the set of fertility, viability, and gametic selection parameters that best ac-that would be only 20 years. Clearly, the ACT2 gene is
not redundant and is almost certainly required for the count for observed dynamical data from populations
with insignificant rates of outcrossing. These maximum survival of Arabidopsis. Third, the mathematical
frame-work developed here greatly facilitates the analysis and likelihood estimates are the parameter values that to-gether maximize the composite likelihood of the ob-detection of potentially deleterious genotypes and
al-leles in multigenerational studies. It should be equally served multigenerational frequencies of the three ge-notypes. With data from sufficient generations, it is useful in dissecting selection parameters in other
experi-mental and natural populations with low rates of out- possible to both estimate the fitness parameters and test the fit of the underlying model to the data. A computer crossing, where selection is weaker and the change in
Hartl, D. L.,andA. G. Clark,1997 Principles of Population Genetics.
and their 95% confidence intervals based on
multigen-Sinauer Associates, Sunderland, MA.
erational adult data is currently under development and Karlin, S.,1969 Equilibrium Behavior of Population Genetic Models with
Non-Random Mating. Gordon and Breach, New York. will be presented elsewhere, along with an application
Kimura, M.,andT. Ohta,1971 Theoretical Aspects of Population
Genet-to the complete Arabidopsis act2-1 study, which is now
ics. Princeton University Press, Princeton, NJ.
in progress. Mandel, S. P. H., 1959 The stability of a multiple allelic system.
Heredity 13: 289–302.
Wyatt W. AndersonandJames L. Hamrickprovided productive
Mauricio, R.,1998 Costs of resistance to natural enemies in field
suggestions and encouragement to this research, andMichael A. D.
populations of the annual plant, Arabidopsis thaliana. Am.
Natural-Goodisman, Gay Gragson,andMarı´a S. Sa´ nchezprovided helpful ist 151: 20–28.
comments on an earlier draft. This work was funded by grant support McDowell, J. M., S. Huang, E. C. McKinney, Y.-Q. AnandR. B. Meagher,1996 Arabidopsis thaliana contains ten actin genes
from the National Institutes of Health (NIH) to R.B.M. and from
encoding six ancient protein subclasses. Genetics 142: 587–602. NIH and the National Science Foundation to M.A.A.
Meyerowitz, E. M.,1989 Arabidopsis, a useful weed. Cell 56: 263–
269.
Meyerowitz, E. M.,1994 Structure and organization of the
Arabi-dopsis thaliana nuclear genome, pp. 21–37 in ArabiArabi-dopsis, edited
byE. M.Meyerowitzand C. R. Summerville. Cold Spring
LITERATURE CITED
Harbor Laboratory Press, Cold Spring Harbor, NY.
Overath, R. D.,andM. A. Asmussen,1998 Genetic diversity at a Abbott, R. J.,andM. F. Games,1989 Population genetic structure
single locus under viability selection and facultative apomixis: and outcrossing rate of Arabidopsis thaliana (L.). Heynh. Heredity
equilibrium structure and deviations from Hardy-Weinberg
fre-62:411–418.
quencies. Genetics 148: 2029–2039.
Asmussen, M. A., J. ArnoldandJ. C. Avise,1989 The effects of
Snape, J. W.,andM. J. Lawrence,1971 The breeding system of
assortative mating and migration on cytonuclear associations in
Arabidopsis thaliana. Heredity 27: 299–302.
hybrid zones. Genetics 122: 923–934.
Weir, B. S.,1970 Equilibrium under inbreeding and selection. Ge-Gilliland, L. U.,M. A. Asmussen, E. C. McKinney and R. B.
netics 65: 371–378.
Meagher, 1998 Detection of deleterious genotypes in
multi-Weir, B. S.,1996 Genetic Data Analysis II: Methods for Discrete Population
generational studies: I. Disruptions in individual Arabidopsis ac- Genetic Data. Sinauer Associates, Sunderland, MA.
tin genes. Genetics 149: 717–725. Workman, P. L.,andS. K. Jain,1966 Zygotic selection under mixed
Goodisman, M. A. D.,andM. A. Asmussen,1997 Cytonuclear theory
random mating and self-fertilization: theory and problems of for haplodiploid species and X-linked genes: I. Hardy-Weinberg estimation. Genetics 54: 159–171.
dynamics and continent-island, hybrid zone models. Genetics