Original Article Correlation between miR-126 expression and DNA hypomethylation of CD4+ T cells in rheumatoid arthritis patients
Full text
Figure

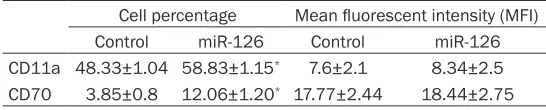

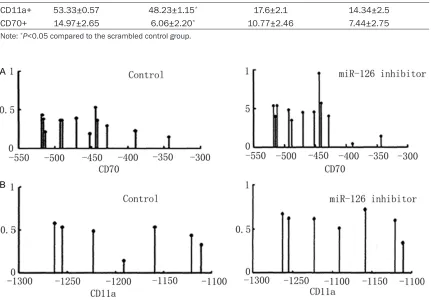
Related documents
They were cultured on Potato Dextrose Agar (PDA) medium. Spores were harvested in 0.1% Tween – 80 solution in double distilled water. Concentration of spores/ml in the
We reveal that exposure to lipids supplied by marrow adipocytes induces expression of lipid chaperone FABP4, pro-inflammatory interleukin IL-1β, and oxidative stress protein HMOX-1
In striving to achieve a higher humanity in the context of human rights and intellectual property, domestic economic policy should be implemented to attain the
ARDS: Acute respiratory distress syndrome; BPD: Bronchopulmonary dysplasia; CI: Confidence interval; CPAP: Continuous positive airway pressure; CRIB-II: Critical risk index
Using the sodium iodide symporter (NIS) as theranostic gene, non-viral gene delivery vehicles based on linear polyethylenimine (LPEI), polyethylene glycol (PEG)
Instead of unsupervised classification technique , we have used supervised classification technique by using feed forward back propagation neural network. Supervised approach
Purpose: To report outcomes of phakic intraocular lens (IOL) implantation after deep anterior lamellar keratoplasty (DALK) to correct high ametropia.. Setting: Centro
![1,1 Bis(4 fluorophenyl) 3,4 dihydro 1H 1,3 oxazino[3,4 a]indole](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)