PROPRIOCEPTION IN INSECTS
II. THE ACTION OF THE CAMPANIFORM SENSILLA ON THE LEGS
BY J. W. S. PRINGLE, B.A.
From the Department of Zoology, Cambridge
(Received 14 March 1937)
(With Thirteen Text-figures)
INTRODUCTION
THE previous paper described the action of the campaniform sensilla on the palps of Periplaneta as stress receptors responding to strains in the cuticular skeleton. These sense organs are widely distributed in insects, and we shall here consider their mode of action in the legs, where, as has been shown by Mclndoo (1914-20), they occur throughout the Class. As in the previous paper, the American cockroach,
Periplaneta americana, has been used for most of the experiments, and the methods
employed are identical to those already described; sensory impulses in the nerves from the sensilla are recorded by means of an amplifier and Matthews (1928) oscillograph.
A considerable amount of descriptive morphology is included in the following account, as it is necessary for a clear understanding of the working of the sensilla. In the latter part of the paper previous theories of their function are discussed, and finally an attempt is made to interpret their mode of action in terms of their known histological structure.
RESULTS
General morphology of the cockroach leg
The legs of the Periplaneta are all alike except for minor details of the size of the segments. The Ilnd leg is illustrated in Fig. 1, from which the general structure can be seen.
The coxa is articulated at the base to the pleuron by an extrinsic joint (pl.c.) with sclerotized condyles, and to the trochanter, a flexible projection of the pleuron, by a complicated hinge joint (h.). It can therefore pivot about the line joining these two, and can also rotate about the pleural attachment as far as the flexibility of the trochanter will allow. Its muscles are complicated, and need not be described.
Proprioception in Insects 115
[image:2.451.121.326.147.243.2]standing position (Fig. 2) is between these two extremes, so that the coxo-tro-chanteral hinge is not quite vertical. The muscles moving the trochanter are a flexor and an extensor (Fig. 3). These are termed levator and depressor respectively by Snodgrass (1935), but it seems simpler to describe them by their action on the joint and not by their supposed effect in moving the animal as a whole. The flexor
Fig. I. The Ilnd right leg of Periplaneta americana. Ventral view showing the types of articulation of the segments b ball and socket joint between tibia and tarsus; ex. coxa; fm. femur; h. hinge joint between coxa and trochantin ; pl.c. condylic joint between coxa and pleuron; pts. pretarsus;
tb tibia; tn. trochantin, tr. trochanter; tt. tarsus
is attached by a single chitinous apodeme to the base of the trochanter round the inner side of the joint; the extensor has one main apodeme attaching to the outer side of the trochanter and two accessory apodemes (Fig. 4, ext' Jr.). These last are well developed only in the second and third pairs of legs, the extensor trochanteris of which is used in the running movements of the insect. The trochanter contains
Fig 2 Fig 3
Fig 2. Pertplaneta americana, seen from below in the normal standing position.
Fig. 3. Trochanter of the Illrd right leg of Pertplaneta, dorsal view. c. coxo-trochanteral condyle;
ex. coxa; ext.tr. apodeme of the extensor trochanteris muscle; fl tr. apodeme of flexor trochantens; fm. femur, h hairs on the trochanter; tr. trochanter; i, 5, groups of campaniform sensilla.
but a single muscle, the reductor femoris, inserted along the dorsal side of the base of the femur.
The femur is firmly attached to the trochanter by an intrinsic hinge (Fig. 4, h.) on the ventral side. The cuticle forming this hinge is very tough and the pull of the reductor muscle is balanced by the elastic restoring force of the hinge. The hinge
[image:2.451.51.406.371.513.2]n 6
J. W. S. PRINGLEline, also, is not straight, so that any movement results in a distortion of the cuticle of the whole segment, which further resists movement. In the normal standing position the whole weight of the body is taken on the elasticity of this hinge, con-traction of the reductor femoris serving only to lower the insect further on to the ground and, if strong enough, to break the hinge and amputate the limb.
The tibia articulates with the femur by another pair of extrinsic condyles, giving a single plane of movement. The femur is slightly twisted, but the plane of the femoror-tibial hinge is also nearly vertical in the standing position. The two muscles, flexor and extensor (depressor and levator respectively of Snodgrass) act antagonistically, the flexor being the larger of the two.
The tarsus provides an example of extrinsic monocondylic articulation. The tibio-tarsal joint is of ball and socket type (Fig. i, b.), the two muscles being both depressors and attached on either side; thus besides the ordinary movement of
[image:3.451.114.343.255.391.2]fi ir
Fig. 4. Trochanter of the 111 rd right leg of Penplaneta, ventral view. c. coxo-trochanteral condyles;
ext.tr. extensor trochantens; ext.tr accessory apodemes of extensor trochanteris j fl.tr. flexor
trochantens; h. trochantero-femoral hinge joint; 2, 3, 4, groups of campaniform sensilla.
depression, a sideways movement could be produced by relatively stronger con-traction of one muscle or the other.
The tarsal segments and the pretarsus articulate by intrinsic hinges with one side toughened as in the palps. But from the end joint inwards there is a gradual increase of sclerotization and infolding of the cutical, so that the joint between the first and second tarsal segments is practically also a ball and socket joint. There is a small depressor pretarsi, with a long thread-like apodeme passing back to the femur, where lies most of its muscle.
Sensory arrangement
Campaniform sensilla occur on the legs in the following positions: four groups on the trochanter, one on the base of the femur on the dorsal side, one on the outside of the base of the tibia, and a group on each of the segments of the tarsus (Figs. 3, 4, 5 and 6).
Proprioception in Insects
1173rd Is
2nd Is
Fig. 6
Fig. 5. Femero-tibial joint of the Illrd left leg of Penplaneta, dorsal view. c. condyle; ext.tb. extensor tibiae; fl.tb. flexor tibae; 6, group of campamform sensilla.
[image:4.451.68.395.66.274.2]Fig. 6. Tarsus of the Illrd leg of Penplaneta. Dorsal view to show the position of the campaniform sensilla. ind U , 3rd tt. second and third tarsal segments; 8, 9, groups of campaniform sensilla
Table I. Number of campaniform sensilla on the legs of Periplaneta americana.
The right 1st leg had only four tarsal joints
Sensilla group
Leg I R.
L. II R. L. Ill R. L. 1 2 0 18 19 20 20 18 2 19 2 0 19 19 2 0 19 3 IS 18 18 18 19 19 4 13 13 14 13 15 15 5 10 9 7 7 8 7 6 8 8 7 7 11 11 7 2 1 1 1 1 1 8 2 2 2 2 2 1 9 2 2 2 2 2 IO 2 I I I I I II I I I I I I
Trochanter and femoral groups
If the trochanter and femur are treated with boiling potash to remove all but the cuticle, and are then cleared in clove oil, the sensilla are immediately obvious under a magnification of 50 x and have the appearance of pores, their thin covering membrane being almost transparent. The detail is shown in Fig. 7. The sensilla vary in shape and size, but are rarely quite circular and sometimes long and narrow. Their length varies from 6 to 24p. They are arranged more or less regularly in lines, with the long diameters of the sensilla diagonal to the direction of the lines. Sometimes there is indication (Fig. 7, 3) of a second alignment, at an angle to the first. When this is apparent, the direction of the long diameters of the sensilla bisects the angle between the two lines.
[image:4.451.41.410.329.435.2]the groups of sensilla on the ventral side of the trochanter of the Ilird leg show that there are here 52 fibres, varying in diameter from 6 to 15/x. Although two different specimens were concerned, this compares well with the 53 sensilla in groups 2, 3 and 4 of the Ilird leg, as shown in Table I. The sections were cut just before this nerve joins the main trunk of the leg, and evidently at this point there
»* 1
[image:5.451.98.357.156.481.2]# #
Fig. 7. Details of the orientation of the groups of campaniform sensilla on the Ilird legs of
Pert-planeta. The numbering corresponds to that in Table I, but the drawings were made from
different specimens from those used for Figs. 3, 4 and 5
has been no joining up of fibres (compare the sensilla on the palp; Pringle, 1937). Higher up the nerve the number of fibres is so large that it is impossible without serial sectioning to tell which comes from which sense organ.
Tibial and tarsal groups
Proprioception in Insects 119
The tarsal groups (7-11) occur on the distal ends of tarsal segments 1-5. There are no campaniform sensilla associated with the tibio-tarsal joint. Fig. 6 shows the sensilla on the tarsal segments 2 and 3. They lie on the overlapped part of the segment proximal to the joint, and their long diameters are always longitudinal to the limb. The long apodeme of the depressor pretarsi runs along the opposite, i.e. the under, side of the tarsal joints.
Physiology
[image:6.451.42.410.209.416.2]The fibres from the leg campaniform sensilla all run in the larger of the two nerves to the leg, and impulses from them can be recorded in it. It has not been possible to study the organs singly, and all records therefore show the combined
Fig 8. Discharge in the large nerve from the Illrd leg of Penplaneta following light (A) and strong (B) pressure on the side of the trochanter. Time in I/IO sees.; recording from the base of the coxa.
effect of impulses in many fibres. The types of response shown in Fig. 8 are obtained by pressure with a glass needle on the side of the trochanter. This distorts the cuticle and will set up large forces in its surface. Evidently the sensilla here are reacting like those on the palp to stresses in the cuticle (Pringle, 1937). Sometimes, with less strong pressure, a regular rhythm is observable (Fig. 8 A), though the large variation in impulse size shows that more than one fibre is active. A regular adaption can be observed in this record and is difficult to explain except as the action of a single fibre with other impulses superimposed; in which case the total number of active fibres in the nerve at this point (the base of the coxa) cannot be very large.
supply to the leg is removed and the muscles relax. The central end shows a large spontaneous discharge, which is likely to include the normal mechanism for main-taining tone. With the muscles relaxed, the trochanter and the leg distal to it may be moved on the coxa as far as the dicondylic hinge will allow, and very little activity is observed in the nerve during this movement; there are certainly none of the large impulses from the campaniform sensilla.
Any movement of the femur on the trochanter, however, gives rise to a discharge similar to that obtained by pressure on the trochanter. This is easily understood from the fact that the trochantero-femoral hinge is not straight but curved, and cannot therefore be bent without distorting the whole of the cuticle in the neighbourhood. This will produce the necessary strains for stimulation of the campaniform sensilla. It has already been explained that in normal standing the weight of the body is taken on the elasticity of this hinge; the intensity of excitation of these sensilla will thus be dependent on the vertical pressure on the legs.
Passive bending of the dicondylic femoro-tibial joint also produces a slight discharge of impulses in the nerve, but it is difficult to be sure that some of the long spines are not being stimulated by this movement, and these are known to give an incompletely adapting discharge (Pumphrey, 1936). The campaniform sensilla can, however, be strongly excited by any sideways bending of the joint out of the normal plane of movement.
The response of the tarsal groups has not been studied. Any touching of the tarsus produces such a large discharge of impulses from the tactile hairs that further activity in the nerve would not be noticeable. Neither has it been possible to investigate the effect of active contraction of the muscles, though stresses would certainly be set up in the cuticle if such contractions were resisted by an outside force. But before discussing in general the function of these leg sensilla, it will be necessary to consider more closely the nature of the stimuli to which they respond. This involves a consideration of the structure of the sensillum, and a review of previous theories of its function.
The function of the campaniform sensilla
Terminology. The term "sensilla campaniformia" was proposed by Berlese
(1909) to include the whole class of insect sensilla that resembles in general characters those first described by Hicks (1857). Minor structural variations had produced in the literature a wealth of different names, some of which, particularly "Gruben ohne Kegel", "Sinneskuppeln", and "Papillen" of German writers, still persist. But Berlese's term is now in general use (Snodgrass, 1935).
Structure. The morphology has been extensively examined and is now relatively
clear (Fig. 9). It may be summarized as follows:
Proprioception in Insects
1 2 1direction; with the cap membrane thickened along the long diameter of the oval (sensilla on the halteres of Diptera, (Pflugstaedt, 1912); and on the appendages of
Periplanetd).
(3) The cap membrane usually domed above the surface of the cuticle, but sometimes recessed so that its top is level with the surface ("scapalen Papillen" of the halteres, Pflugstaedt, 1912; Fig. 9 D).
(4) Sensilla usually occurring in groups with a definite arrangement and orien-tation (Mclndoo, 1914).
[image:8.451.136.320.236.342.2](5) The single sense cell large, with a distal process inserted on the under side of the middle of the domed cap by a highly refracting body, which in oval sensilla may also be elongated on the long diameter. It is this body that makes the dome
Fig. 9. Types of campaniform sensilla A. Diagrammatic. B, C. Sections through the long and short diameters of a sensdlum on the haltere of Calliphora. D. Sensillum on haltere of Syrpkus. E. From the cercus of Penplancta. F. From the labium of Dytiscus G. From the mandible of
Dytiscus. a, b, outer and inner lamellae of cap membrane; c, cuticular connexion of d, the distal
process of the sense cell. (From various authors, after Snodgrass, 1935.)
appear to be a pore when viewed with certain arrangements of transmitted light, as wrongly described by Mclndoo (1914) and Newton (1931).
(6) Other modified hypodermis cells usually absent, but apparently chitogenous cells occur in some cases (Sihler, 1924).
The sensilla have been described from a variety of positions in many insects. Snodgrass (1935) lists: "head, thorax, abdomen, the antennae, mouthparts, legs, wing bases, cerci, and ovipositor of adult insects,... also... on the larvae of some species". In addition, the halteres of Diptera and Strepsiptera are richly provided at their base (Pflugstaedt, 1912; Ulrich, 1930). Mclndoo, in a series of papers from 1914 to 1920, gives tables of the number of sensilla in various positions in different insects, but usually only as totals of a number of groups. His paper of 1914 gives a more detailed description of the main groups on the legs, wings, and sting of
Apis. Unfortunately he is not always accurate in his descriptions and seems to
more likely sockets where hairs have been shed; these can resemble campaniform sensilla very closely in untreated material, but after clearing with clove oil the characteristic structure of the sensillum is at once obvious. Mclndoo's results are thus likely to be unreliable as to the number of sensilla, but his papers do show that these occur very generally thoughout insects.
Function. Mclndoo alone among investigators of the campaniform sensilla
regards them as olfactory in function. His work must be examined in some detail. He described their histology and published figures (Mclndoo, 1914) showing the protoplasm of the distal sensory process freely exposed to the exterior. Newton (1931) reinvestigated the question, and established that there is a definite cuticular covering over the cell process, and that the pore of Mclndoo was an artefact due to treatment of the preparation. The existence of this covering does not, however, exclude the possibility of the sensillum having a chemoreceptive function, as a number of workers have shown that chitin, when unimpregnated with other sub-stances, is highly permeable (Wigglesworth, 1933). Mclndoo's other evidence must therefore be considered.
In many of his papers he describes experiments purporting to show that the "olfactory pores" really have this function. All the experiments are similar in form. The insect is placed in a gauze cage, and left until it becomes stationary. The odour under test is then brought up under the cage and any reaction on the part of the insect noted. Any movement is taken as a positive result, and the reaction time is measured with a stop-watch. By measuring the reaction time of individuals with various numbers of their "pores" removed or covered up with some substance he finds a rough relation between average reaction time and number of pores left. The odours tested were usually strong essential oils such as pepper-mint, wintergreen, bergamot, etc.
The method is crude and liable to a number of objections. No attempt is made to exclude other stimuli, such as light intensity and air vibration which might stimulate the insect to movement. The effective concentration of the odorous chemical at the surface of the sense organs will be affected by draughts and will thus not be constant throughout the series of experiments. And the reaction time depends on other things besides the intensity of excitation of one type of sense organ; it is affected by the whole state of the central nervous system, which will be seriously upset by amputation of any part of the body. Even the amount of handling to which Mclndoo's insects were subjected would be enough to slow down their reactions.
Proprioception in Insects 123
response obtained to pressure on the trochanter, and the experiment suggests strongly that the campaniform sensilla have nothing whatever to do with olfaction. The alternative view, that they are mechanical receptors of some type, has been put forward by several authors; and Weber (1933) quotes Demoll as suggesting more definitely that they might serve a proprioceptive function by responding to bending of the cuticle. No possible mechanism, however, has been described.
The mechanism of the campaniform sensilla
The experiments already described suggest that the campaniform sensilla are mechanical receptors, responding to stresses in the cuticle. This behaviour has been verified experimentally for sensilla on the palps and legs of Periplaneta. It remains to show that such a function is possible for all such organs wherever they occur, and to define more accurately the adequate stimulus.
With the exception of some of Mclndoo's account, which have been shown to be unreliable, there is no case known of a campaniform sensillum occurring in a part of the cuticle which would not be liable to strains.
The main groups are located at the limb joints, where stresses are set up by bending and muscular contractions, and at the wing and haltere roots where are concentrated the forces that move the organ. Other well-established positions are at the base of the barbs of aculeate stings (Mclndoo, 1914), on the mandibles of various biting insects (Dytiscus, Hochreuter, 1912; bees, Janet, 1911), and on the oscillatory gills of ephemerid nymphs (Eastham, 1936). In all these cases stresses will occur in the cuticle under normal conditions. And though the sensilla described by Sihler (1924) from the cerci of Orthoptera would seem at first to be an exception as the cercus as a whole should not be liable to strains, it is clear from his descriptions that the sensilla occur only in close connexion with structures at the base of the hairs, and on portions of the cuticle which are likely to be strained when the hair is bent.
There is nothing then in previous publications that contradicts the view.
Primitive type
A simple type of sensillum is found on the segments of the thoracic legs of the caterpillar of Smerinthus ocellatus (Lepidoptera, Sphingidae). It is circular in appearance, and in cleared whole preparations the only structure that can be distinguished is a slightly heavier pigmentation of the sides of the canal. The whole of the cuticle is very elastic and flexible except for the claws at the end of the leg. A slowly adapting discharge of impulses can be detected from a few fibres in the nerves to the legs when the cuticle is distorted, and the behaviour thus seems to be similar to that of other sensilla. It can be supposed that the normal adequate stimulus is resistance to the contracting of the leg muscles by contact of the claw with an outside object.
of the sensillum could produce enough local movement to excite the end of the nerve. The difference in appearance of the lining of the canal may give a clue to this.
Adult type
In adult insects the cuticle is often greatly toughened by deposition in the chitin of other substances (Wigglesworth, 1934); actual bending of the skeleton is reduced to a minimum, and it is necessary to consider how the sensilla can be excited by mechanical stimuli, situated as they often are in the thickest parts of the cuticle. To do this we must first discuss some of the general mechanical properties of surfaces and hollow tubes.
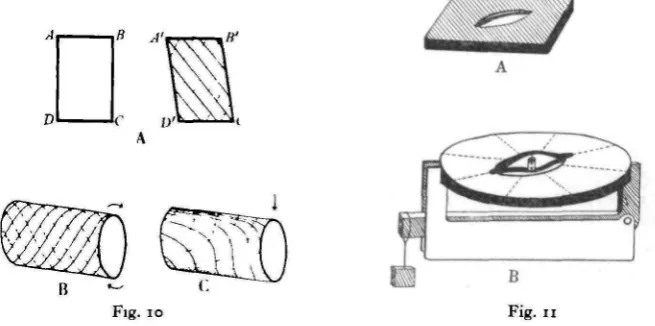
[image:11.451.58.386.226.389.2]A thin, fiat surface can be subjected to two types of strain; to bend by forces at right angles to its plane, and to shear by forces in its plane. Illustrative models
Fig. 10. Diagram to illustrate the resolution of shear forces into compression and extension com-ponents. A, a flat surface; B, C, a hollow cylinder subjected to twist and bend. Continuous lines, extension. Dashed lines, compression. For further explanation see text.
Fig. 11. Diagram of models illustrating the action of the campaniform sensillum. A. To represent the actual sensillum. B Model constructed in a circular framework for measurement of the effect of compression in different directions. See text.
can be made out of sheet rubber, which is sufficiently elastic to allow of visible movements. Fig. 10 A shows a diagram of such a model, made by stretching a piece of 1/16 in. rubber over a rectangular metal frame ABCD. A shearing force can be set up in the rubber by distortion of the rectangle into a parallelogram
(A'B'C'D'). The distance AC is lengthened and BD shortened. The surface material
is in tension in the direction AC, and in compression in the direction BD. Unless there was an initial stretch in the rubber, creases will appear parallel to AC. A shearing force thus consists of tension in one direction and compression in another. It should be noted that the lines of compression and extension need not intersect at right angles.
Proprioception in Insects 125
spiral manner round the tube in opposite directions; with a bending moment, the outer side has extension longitudinal and compression transverse, the inner side extension transverse and compression longitudinal, and the sides a normal diagonal shear. Again it must be noted that the lines do not necessarily intersect at right angles, and with a bending force they are not even parallel. All this applies, of course, only to a homogeneous surface; any variation in thickness or composition alters the position and direction of the compression and extension lines. It assumes also that the forces are applied evenly round the ends of the tube.
Insect limb segments are never articulated evenly all round their base; if they were the articulation would be rigid. Nor are they perfect homogeneous cylinders. The force lines will therefore not be regular. But the important point is that in a hollow structure such as the insect skeleton, all stresses can be expressed as shearing stresses, and resolved into compression and extension components.
The probable mode of action of the campaniform sensilla in registering these stresses can best be explained by reference to a model which shows many of the properties of the sensillum. Fig. 11 A shows one variation of this model. A rubber membrane stretched over a framework is pierced by a circular or oval hole, and across the long diameter of the oval is glued a strip of paper, slightly domed out of the plane of the diaphragm.
The hole in the rubber represents the canal through the cuticle, and the strip of paper the longitudinal thickening of the cap membrane or of the attachment of the nerve process. The rest of the cap is assumed to be relatively more elastic than this thickening and is omitted. This differential elasticity of two parts of the skeletal structure is an essential assumption. It need not be a sudden change of property along a definite line; it can in fact be a gradual grading; but it must exist for the proper functioning of the sensillum according to the present theory. If opacity can be taken as a measure of the thickness of the cuticle or of its degree of impregnation with other substances, and this as the cause or constant accompaniment of increased toughness, then there is good evidence for its existence. For in all adult sensilla examined the surface view in transmitted light shows a clear line down the long diameter of the cap (Fig. 12).
Slight distortion of model 1 from the rectangular shape produces in one direction a lengthening of the hole, and in the other a shortening. This causes the paper strip to be more or less humped, and its middle point moves on a line at right angles to the plane of the diaphragm. If the end of the nerve process is imagined as being inserted on the under side of the middle of the paper strip, it will be stretched by one distortion and compressed by the other. One or both of these movements could lead to excitation of the ending.
diaphragm. The model was fixed horizontal and a small electrolytic cell constructed on the lower of the two strips. This consisted of a short length of glass tubing fixed vertically with one electrode inserted through the paper from below. The other electrode was mounted on a straight piece of wire from the upper strip and was free to move inside the glass tube. Both electrodes had circular disks of platinum foil as their main surfaces. With the tube filled with an electrolyte (dilute salt solution) a cell is thus made with the interelectrode distance variable; provided the upper electrode moves vertically, the resistance of such a cell is roughly proportional to the distance between the electrodes.
[image:13.451.96.340.213.387.2]Model 2 was made in a circular frame consisting of a pair of wooden embroidery rings. This made possible the determination of the effect of distortion in various directions. With the arrangement shown in Fig. 11 B any required distorting force
Fig 12. Photomicrograph of the sensilla of group 4 on the Illrd leg of Periplaneta, showing the appearance in transmitted light Note the doming and longitudinal thickening of the cap mem-brane. From a potashed preparation cleared in clove oil.
can be applied in any direction. The model has the disadvantage that in it the compression and extension lines always cross at right angles, and it is therefore not possible to reproduce the diagonal intersections that occur in the insect cuticle. Owing, also, to the rigidity of the oak rings of the framework, large forces are required to produce a distortion, and only a small proportion of the restoring force will be derived from the elasticity of the rubber diaphragm.
Proprioception in Insects 127
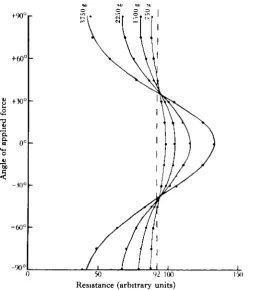
electrode which will introduce an error into the readings. This factor would not appear in the actual sensillum, in which forces with 45 ° orientation should produce no movement.
There is thus a critical angle below which there is an increase and above which there is a decrease of doming. If it be assumed for the moment that excitation only occurs with increased doming, then the sensillum will be excited when the com-pression component of the shear force makes an angle of less than a certain angle with the direction of thickening of its cap membrane; for force lines intersecting at right angles the critical angle will be 45 °.
•a
| "a
a
+90°
+60°
+30°
0°
-K)°
-60°
-90°
Si*
\
-/
. / 1
225(
]
\
/
I
I
I
i i
1 \ 1 ' 1 1
NX
J /
/ / /
_L1
\\
i i
50 92 100 Resistance (arbitrary units)
[image:14.451.100.358.199.490.2]15*1
Fig. 13 Graph obtained from model 2, showing the relation between the angle of t^ie applied compression force and the resistance of the cell, for various values of the compression force.
The sensitivity of the sensillum will be dependent among other things on the absolute length of the long diameter of the cap. For a sensillum of given area, a greater sensitivity will thus result from an oval shape. In the various groups of sensilla on the legs of Periplaneta there is often a considerable variation in length. This, if other things are equal, should give a variation in sensitivity among the individuals of a group.
Since over any given small area of the cuticle the compression lines will always be running approximately parallel, the important rule may thus be laid down, that
group of such sensilla will act as a unit, and if innervated from the same fibre will be extending the range of that fibre quantitatively but not qualitatively. This condition we have already seen in the sensilla of the palps of Periplaneta (Pringle, 1937), and even without proof of a sensory anastomosis in the legs, it simplifies very con-siderably the interpretation of the behaviour of these sensilla also.
Unidirectional sensitivity
The assumption made above that each sensillum reacts only to. the compression component of the shear force must now be justified. Owing to the irregularities of the cuticle of the legs it has not been possible to prove this objectively, and it rests only on a number of circumstantial considerations, none of which is clusive. The point is therefore not finally settled, but the following may be con-sidered as evidence:
(1) In each of the leg groups in Periplaneta except group 6, the sensilla are all orientated in the same direction. If the sensillum were equally sensitive to the compression and extension components of shear, an equally efficient system would result if some were at an angle of 900 to the others (for force lines intersecting at right angles). The fact that this arrangement is not found suggests that only one component is effective.
(2) Group 6 on the tibia of Periplaneta is exceptional in that two directions of orientation are apparent. In some insects this group is represented by two groups
{Apis, Mclndoo, 1914). If each sensillum reacts only to one component, then the
functional duplicity is preserved in Periplaneta, though all the sensilla are grouped together.
(3) It is very noticeable with many hair sensilla that only one direction of movement is effective. This was noted by Pumphrey (1936) for the tibial spines of
Periplaneta, and is also true of certain hairs on the metathoracic spiracle (author's
observation of the sensory impulses). Histological studies have shown that the nerve fibre is inserted on one side at the base of insect hairs in very much the same way as under the dome of the campaniform sensilla. The same behaviour should therefore be shown by both types of sensillum.
(4) The sensilla on the tarsus of Periplaneta are situated on the upper surface and have their long diameters parallel to the length of the leg. When the tarsus is pressed against the ground by contraction of the depressor metatarsi, the compression lines in the cuticle of the upper surface of the segments will also be longitudinal. If only one component is effective, then it must be the compression component, for under no normal circumstances can the reverse condition (tension lines longitudinal) be produced in the living insect.
Proprioception in Insects 129
Discussion of the function of the campamform sensilla on the legs
of Periplaneta
With the above theoretical background we can now return to the question of the function of the sensilla on the legs of Periplaneta. Each group must be considered in turn.
The tarsal sensilla present no difficulty; as stated above, they are orientated parallel to the only direction in which a compression component of shear can normally be set up, and they should react to the pressure of the tarsal segments on the ground. But the others are more complicated.
Group 6 of the Ilnd and Illrd legs is situated in a region where the cuticle is irregular, owing to the proximity of the thickened edge round the base of the first tibial spine. Its two subgroups are orientated almost at right angles to each other, and in general direction the larger group is at an angle of about 700 to the length of the tibia. This makes its function difficult to decide. In the 1st leg the cuticle is more regular and the orientation is almost exactly longitudinal and transverse, which would make it respond to the bending forces produced by resistance to the contractions of the extensor and flexor tibiae. This may also be the function of group 6 in the Ilnd and Illrd legs; but there is also the possibility here, owing to the fact that in normal standing the femoro-tibial hinge of these legs is nearly vertical, that it may respond rather to the sideways bending that must occur under these conditions. That this movement is certainly effective was shown experimentally for these legs, but it was not excluded that resistance to straight extension or flexion might not also excite. In the 1st legs, the coxa is normally rotated until this hinge is nearly horizontal, so that the distinction does not arise, but in the hind two pairs it becomes important to distinguish between the sideways forces present in the standing position and those produced by the movements of running. Possibly both these excite group 6.
In the case of the trochanteral and femoral groups deductions from anatomy become even more difficult owing to the great divergence of this region from the cylindrical shape. It can only be pointed out again thar in the normal standing position there will be a force tending to bend the trochantero-femoral hinge, and that it has been shown experimentally that this excites some of the groups of sen-silla.
When the insect is standing on the ground, or when it is hanging inverted, there will, then, be excitation of at least some of the groups of campaniform sensilla (certainly 6-11, and some or all of 1-4). This behaviour provides a factual basis for a sense that has many times been postulated from experiments on the general behaviour of insects. Some of these may be considered.
In the cockroach itself, Hoffmann (1933) has described a righting reflex which is initiated by the absence of contact of the legs with the ground. Hoffmann was unable to decide between tactile and pressure stimuli as the normal means of inhibiting this, but the following experiment shows that the latter are responsible. If all but the first tarsal segments of the legs of a cockroach are amputated, glass
rods can be inserted in the cut ends of the legs in such a way that there is no stimulation of any tactile hairs. An insect so treated appears to be perfectly normal in its standing and locomotion. If placed on its back, the movements of the righting reflex at once appear, and can be inhibited again, as shown by Hoffmann, by lowering a glass plate on to the insect. This inhibition appears when there is no contact with the plate except by the tips of the glass rods in the legs, the pawing movements ceasing in any leg of which the rod can be pressed against the plate. It seems evident that it is excitation of the campaniform sensilla that is normally producing the inhibition of the reflex.
Fraenckel (1932) and Hoist (1935) describe further effects due to contact of the legs with the ground. Fraenckel relates the onset of flight to the removal of this stimulation, while Hoist assigns to pressure stimuli on the legs an important role in the co-ordination of walking movements, both in insects of all sorts, and in myriapods, and Crustacea. Campaniform sensilla do not, indeed, occur in some of the animals he mentions, but it is possible, nevertheless, that some similar cuticular strain sense is at work throughout. It may, perhaps, be found to be a general rule for arthropods that here, in direct contrast to the vertebrate plan, the force exerted on the limbs by the weight of the body and the contraction of muscles is measured, not by the tension in the tendons of the effectors, but as compression in the skeletal supports. If this be so, it must modify to a large extent our conception of the role of proprioception in this phylum. The conclusions of Crozier & Stier (1928-9), who find in a differential force sense in the legs a mechanism for explaining geotropic orientation, must be reconsidered if this sense does not, as they thought, provide a separate measure of the state of each muscle. The campaniform sensilla may not be the sole proprioceptive sense organs in the leg (Slifer (1936) has recently shown that chordotonal organs are much more widely distributed than had been thought), but the large size of the fibres supplying the campaniform sensilla (cp. Barnes, 1930), and the type of stimulus to which they respond, suggest that they are an important part of the reflex system. They may provide the only force sense in the legs.
Proprioception in Insects 131
SUMMARY
1. The campaniform sensilla on the legs of Periplaneta are similar in action to those on the palps, and respond to strains in the cuticle.
2. They are arranged in groups at the joints, with parallel orientation of the sensilla of a group.
3. Tests with various chemical substances show a complete absence of sensitivity to olfactory stimuli.
4. A theory is given of the mode of action of the sensilla in terms of a mechanical model based on their observed structure. Each group of parallel sensilla should act as a unit, responding to those forces which have a compression component of shear in the direction of their long diameters.
5. This theory makes it possible to predict the behaviour of the sensilla from their anatomical arrangement. Most if not all the groups on the legs are so arranged as to be sensitive to the forces present when the insect is standing on the ground. 6. The sensilla probably provide the basis for the sense of contact pressure postulated by Hoist (1935), Hoffmann (1933), Crozier & Stier (1928-9), Fraenkel (1932) and others.
7. Comparison of this propnoceptive mechanism with that of the vertebrate limb reveals an absence of qualitative sensitivity that may have an important bearing on the question of the evolution of behaviour.
REFERENCES
B\RNES, T C. (1930) C R Soc. Bwl., Paris, 105, 385. BERLESE, A. (1909) Git Insctti. Milan.
CROZIER, W. J. & STIER, T . J. B. (1928-9). J. gen. Pkynol 12, 675.
EASTHAM, L . E. S. (1936). Trans. R. ent. Soc. Land. 85, 401 FRAEJIKEL, G. (1932) Z. vergl. Pkynol. 16, 371.
HICKS, J. B. (1857) J Linn. Soc. (Zool.), 1, 367. HOCHREUTER, R. (19:2). Z tvui. Zool. 103, 1. HOFFMANN, R. W. (1933). Z. vergl. Phystol. 1 8 , 740.
HOLST, E. V. (1935). Bwl. Rev. 10, 234.
JANET, C H (191 I ) . Apiculteur, 55e Annee, No 3, Mars, p. 107 M C I N D O O , N . E. (1914). J exp. Zool. 16, 265.
(1915) Bwl. Bull. Wood's Hole, 28, 407. (1917). J. comp. Neurol. 29, 457. (1918) J.Morph 31, 113. (1920) J. comp. Neurol. 3 1 , 405. MATTHEWS, B. C. H . (1928). J. Pkynol. 6 5 , 225. NEWTON, H. C. F . (1931). Quart. J. rmcr. Set. 74, 647.
NICHOLAS, J. S. & BARRON, D H. (1935). J. comp. Neurol. 6 1 , 413. PFLUGSTAEDT, H. (1912). Z. wiss. Zool. 100, 1.
PRINGLE, J W. S. (1937). J. exp. Bwl. 15, 101.
PUMPHREY, R J. (1936). J. Phystol. 87, 6 P . SIHLER, H (1924). Zool. jfb., Abt. Anat. 4 8 , 519. SLIFER, E H (1936). Ent. News, 47, 174.
SNODGRASS, R. E (1935). Principles of Insect Morphology. New York. ULRICH, W. (1930) Z Morph. Okol. Tiere, 17, 552.
VOGEL, R. (1911). Z. ivtss. Zool. 9 8 , 68.
WEBER, H. (1933). Lekrbuch der Entomologte. Jena. WIGGLESWORTH, V. B. (1933). J. exp. Biol. 10, 1.
(i934)- Insect Phynology. London.