SOME OBSERVATIONS ON POTENTIAL
DIFFER-ENCES ACROSS THE ECTOPLASM MEMBRANE
OF PARAMECIUM
BY TAKEO KAMADA.
(Tokyo.)
(From the Laboratory of Experimental Zoology, Cambridge.)
(Received i6thjuly, 1933.)
As might be expected, a measurable potential difference is found between a pair of micro-electrodes, one of which is inside a Paramecium and the other in the external medium. The magnitude of this potential depends on the nature of the external solution. A brief account of the influence of the environment on the nature of the potential observed is given in the following pages.
The non-polarisable micro-electrodes were fundamentally the same as those described by Ettisch and Peterfi (1925). They differed from those used by these workers in that the capillary portion was made of quartz and rilled, not with agar, but with the external solution to be investigated (unless otherwise stated). The tip of the capillary, which was inserted into the cell, was generally 2-3/x in external diameter. The electrode in the external medium had a considerably wider tip (20-30/x). The tip of the electrode contained the external solution while the rest of the capillary was filled with a gel of 1 per cent, agar made up in a 10 per cent, dilution of frogs' Ringer. This prevented the hydrostatic flow which might otherwise occur at the tip. Non-polarisable electrodes of the Cu-CuSO4 type were found to be satisfactory.
Membrane Potential of Paramecium 95
This current may give rise in passing through the high resistance of the micro-electrode system (mainly that of the tip) to an E.M.F. which will bev • R.V . .
R
-
t =RTs <
a>-Substituting for V the value of 25 volts (in practice 30-32 volts), the ratio of R to S can be calculated if R . i is of the order of \ millivolt (t.e. the smallest E.M.F. detectible by the null method). Then (2) becomes
, 25 x R « o n 25 x io~5 = -p , e , or icr* S= R.
Thus, if R exceeds a hundred-thousandth of S, there will appear in the micro-electrode system a detectible E.M.F., the polarity of which is, unlike that of the
E.M.F. due to asymmetry of the electrode system, independent of the above-men-tioned reversing key. The specific resistance of N/100 KC1 being 8 x 10* ohms (Kaye and Laby's Tables, p. 90), the resistance of the tip of the capillary (2/n in internal diameter and 1 mm. in length) filled with N/100 KC1 is 2-5 x io* ohms. Therefore, with this electrode, if the leakage resistance S is as low as 2-5 x io14 ohms, there should be in the micro-electrode a detectible E.M J . which is not reversed by the reversing key.
Let y represent the value of this E.MJ\ and x the true value of the potential to be measured. The potentiometer readings with the reversing key in the two positions should be (y + x) and (y— x), so that x is given by \ [(v + *) — (y— *)]. In this paper * has been determined by this method. As a matter of fact, y was observable generally when using capillaries with a very fine tip {e.g. 2/x or less in external dia-meter) filled with dilute solution {e.g. N/100 NaCl or less).
The tip of the external electrode was always less than 30/x from the organism, and its position was found to have no influence on the value of the potential.
Since Packard (1925) has shown that light affects the permeability of
Para-mecium,the illumination of the microscope was kept constant. The light was filtered
through a flask of distilled water.
Measurements were made with successful preparations only. These were characterised by absence of deformation, active ciliary movement, normal appear-ance of the contractile vacuole and of cyclosis, and in particular by active behaviour when the electrode was withdrawn. Preparations having these characteristics generally gave a consistent value for the potential difference. However, if in such a preparation the organism rotated about the inserted electrode, the potential was less, probably owing to short-circuiting of internal and external media by leakage around the electrode. A specimen in which the trichocysts had been discharged gave a very low value for the potential.
Since the carbonate equilibrium of a thin hanging drop varies rapidly with external changes in CO2 tension, the/>H of such a drop is difficult to control unless the liquid is strongly buffered. Such buffering is impossible when the effect of a single electrolyte is to be investigated. It was therefore desirable to find a/>H value, in the region of which a change in/>H would not considerably affect the potential. A series of phosphate mixtures was prepared as shown in Table I, in which thepH was
Table I.
Number of solu-tions (for use in subsequent tables)
No. 1 No. 2
No. 3 No. 4 No. s No. 6
pH
6 0 6-4
6-5
72
7-6
8 0
Component
M/10 KH,PO4
c.c.
50-0 500 50-0 50-0 500 50-0
M/10 NaOH c.c.
5-6
126 236 349 427 469
Ml 10 NaCl
c.c.
44-4 374 264 I5-I 7-3
3 1
A modified Clark and Lubbs' phosphate mixture.
graded from 8-o to 6-o, while the concentration of ([Na]+ [/£]) and the ratio [Na]/[X] were kept constant. The observed potentials in these solutions of varying dilution are shown in Table II. Each potential value in this table (and in all tables throughout this paper) is an average of 10 measurements (sometimes 15), and is given in the tables together with its probable error and the room temperature during the measurement.
Membrane Potential of Paramecium
97
Table II.
Solution used (see Table I)
No. i No. 2 No. 3 No. 4 No. 5 No. 6
A74°
M.V.
+ 3-5 ±°-S (i9 + 3'3 ±o-6 (20 + 3-9 ±o-6 (18
+ 6 - 4 ± I - Q ( I 8
+3-8 ±0-9 (19
+4-2±i-o (19
U C
\S°
•5"
• su
C.) .) C.) C.) C.)
(J.)
Dilution
iV/200
M.V.
- i 5 ' 9 ± : - i (20 - i 5-9 ± i ' 3 U9 —16'4 ± i ' 6 (18 -15-2 ± 2 6 (19 -14-9 ± 1 2 (20 - i S - i ± 0 9 ( 1 9
3C.)
S°C.) S°C.) 5°C.)
3
C.)
5
c.) 1
iV/iooo
M.V. — 28-7 ±0-9 (20° - 2 6 - 3 ±1-3 (20° —25-6 ± 0 9 (10° - 2 4 - 0 ± i - 8 (i8° - 2 4 6 ± 1 0 f 19-5
- 2 3 ' 6 ± I - I (19-5
c.)
C.) C.) C.)"(J.) °C.)
Note. (1) The sign is that of the cell interior with reference to the external surface.
(2) " N " represents the total cation (K+Na) concentration in normality.
(3) In these measurements the capillary portion of the micro-electrodes was made of hard glass and filled with a 10 per cent, dilution of frogs' Ringer.
cation, the carbonate equilibrium of which had been adjusted beforehand to give a
pH of 7-2. Hence the external medium was at/>H 7-2 at the moment of preparation
of the hanging drop, and even if thepH changed in time, its effect on the measured potential could be neglected.
It is conceivable that the intracellular pH of the organism may adjust itself to changes in the pH of the external environment, but within the range of solutions here (Table II) employed no such change could be detected by intra vitam staining with neutral red.
The measurements given in Table II represent the difference in potential of the systems, 1 and 2, in the following diagram:
(1) Non-polarisable
electrode 10 % Ringerin capillary ProtoplasmofParame- Externalsolution 10 % Ringerin capillary Non-polarisableelectrode
(2) Non-polarisable electrode
io % Ringer
in capillary External solution 10 % Ringerin capillary Non-polarisableelectrode
If, for the sake of convenience, the electrical homogeneity of protoplasm be assumed, the measured potential is the difference between the potentials (a + b) and c. All other junction potentials are cancelled by subtraction of (1) from (2). According to Sumwalt (1929, p. 196) the junction potential at b or c may have a considerable value, depending on the pore diameter of the capillary tip as well as the dissimilarity of the solutions inside and outside the tip. Hence the value of (b— c) may not be zero unless the ionic composition inside and outside the Paramecium is the same. Table III shows that the nature of the solution in the capillary tip is an important factor in determining the magnitude of the potential. In order to keep the data free from variation due to c, the electrode tip was filled with the external
Table III
External solution No. 4 of Table II
(N/2OO)
N/ioo NaCl
Solution in capillary tip 10 % dilution of frogs' Ringer •No. 4 of Table II (JV/aoo)
JV/40 KCl JV/200 NaCl 10 % dilution of frogs' Ringer
Measured voltage (M.V.)
-15-2 ±2-6 (19-5° C.) -8-2±O-6(2O°C.) - i 8 -3± i - 5 ( i 8 ° C . )
- 7 - i ± 0 7 (20° C.)
-7-4 ± I - I (20° C.) The sign is that of the cell interior with reference to the external surface.
• In this determination quartz capillary was used, the other four measurements were made with hard glass capillary.
tion throughout the measurements in the present paper (unless otherwise stated), and the difference between the following two systems was determined:
(3) Non-polarisable
electrode Externalsolution in capillary
Protoplasm
of Para- Extemalsolution Externalsolution in capillary
Non-polarisable electrode
(4) Non-polarisable
electrode Externalsolution in capillary
External solution External solution in capillary
Non-polarisable electrode
Since the solution inside and outside the capillary tip is the same in system (4), there should be no junction potential at the tip, no matter how small be the pore of the tip. The difference between the potential values of these two systems gives the sum of the potentials (a + d). Although it is impossible to estimate the sign and magnitude of a and d separately, the total potential so obtained is directly comparable to the "injury potential" of muscle or nerve, in the sense that in these tissues the injured and non-injured parts are in contact with the same fluid.
The effect of pure electrolytes is shown in Table IV. The r&le of the cation could be compared by replacing it by another cation of equivalent normality without any other modification in the solution. As has been mentioned, each solution was buffered with a 2 x 10-4 N carbonate solution of its own cation, so that, strictly
Table IV.
External solution
NaCl KCl CaCl, MgCl,
JV/40
M.V. + I I - I ± I - 4 ( I 8 - 5 ° C . )
+ 13-3 ± 0 5 (21° C.) + i9-2±o-6 (20° C.) + i8-5±2-3(2o°C.)
iV/200
M.V.
- 7-1 ±09(19° C.) - 5'5±o-4(i9'5° C.) + 136 ± 1 6 (21-5° C.) + 9-2 ± 0 9 (20° C.)
JV/1000
M.V.
Membrane Potential of Paramecium 99
speaking, the solution was not pure for one anion. It is clear from the table that the observed potential is modified characteristically according to the nature of the cation, and that the two divalent cations give larger positive values as compared with the monovalent cations. The effect of dilution was, however, the same for the four cations tested—the more dilute the external medium, the more negative the charge on the inside of the membrane.
Table V shows that the replacement of Cl' by equivalent SO/' without any other modification in the solution increases the negativity of the cell interior, but the
Table V. External solution NaCl Na,SO4 MgSO4 MgCl,
{ 1 1 + 8 + 11 + 18 N/40 M.V. •I ±1-4(18 •2 ±0-3 (22
•4 ±0-7 (20 •5 ±2-3 (20
s°c.)
5 C.)s°c.)
5 C.) N/zoo M.V.- 7-1 ±0-9(19 - 1 4 7 ±2-2 (19 + 3-1 ±0-8(20 + 9-2 ±0-9 (20
5
c.)
'C.) 5°C.) 'C.) JV/1000 M.V. -19-8 ±O-8(2O° -22-3 ±1-9 (20°- 3-4 ± 0 7 (20° + 5-1 ±0-8(19°
C.) C.)
C.) C.)
The sign is that of the cell interior with reference to the external surface.
influence is less marked than in the case of the exchange of a divalent for a mono-valent cation.
In order to compare the potential increment at each fivefold dilution, the data are collected in Table VI. It is evident that the increment is larger with the mono-valent cations, and that in weaker solutions the effect of dilution becomes smaller.
Table VI. External solution NaCl KCl CaCL MgCl, Na,SO4 MgSO4
Difference between the potential values at
A740 and at N/200 M.V. -18-2 - 1 8 8
- 5-6 - 9 3
-229
- 8-3
Difference between the potential values at
N/200 and at N/1000
M.V. -12-7 - 1 0 5
- 4'S
- 4 1
- 7-6
- 6-s
Calculated from Tables IV and V.
The sign is that of the inside charge of the cell membrane in contact with weaker solution, with reference to the corresponding charge in stronger solution.
Any attempt to interpret the above results must necessarily be tentative, for any explanation at present involves two assumptions.
In the second place the fact that the volume of Paramecium does not increase when the animal is placed in a hypotonic medium (although water is lost in a hyper-tonic medium) seems to indicate that the organism can maintain its internal osmolar concentration constant even when the external medium is diluted.
If the above two assumptions (that the observed potential is mainly the mem-brane potential and that the osmolar concentration of the protoplasm is constant) are made, the membrane potential phenomena in Paramecium may be explained on the theory that the membrane is more permeable to cations than to anions as is the case in the dry collodion membranes of L. Michaelis. It follows from this that the more dilute the external solution is, the more negative is the charge on the internal surface of the membrane (cf. Tables IV, V and VI). The results also indicate that the mobilities of Na and K ions are greater than those of Ca and Mg ions, and that the mobility of S04 ion is less than that of the Cl ion.
Evidence from conductivity measurements (Gelfan, 1927) and from the effect of electrolyte concentration on the flattening of Paramecium and on the discharge period of the contractile vacuole (Stempell, 1914) indicates that the internal concen-tration of electrolyte is roughly equivalent to NJ2O NaCl. Yet when the external solution is iV/40 NaCl the interior is positive to the exterior which is contrary to expectation if the above explanation is correct. This anomaly may be explained by the presence of non-diffusible cations (e.g. protein cations) in the protoplasm of
Paramecium. These would cause a potential difference (E) opposite in sign to that
due to the modified diffusion potential (D) existing between the external and internal salt concentrations. When the organism reaches equilibrium with a medium of divalent cations (Ca, Mg) the interior is always positive to the exterior, indicating that E is numerically greater than D in this case. This agrees with the known order of diffusion potentials of the salts of univalent and divalent cations.
It is of interest to consider the effect of mixing two electrolytes on the membrane potential of Paramecium. It was found that the potential was intermediate between the potentials obtained with the separate solutions provided the total ionic concen-tration was kept the same.
The potential obtained with composite solutions (Em) can be expressed by the
following empirical equation:
£ „ = £, + ( i - a ) ( £ „ - £ , ) (3), where Et and En are the potentials obtained with the two component solutions, I
and II, of equivalent normality and a is the ratio e—;r~o~ for the mixture, where Sl
and Sn are the volumes of the solutions, I and II, respectively. The values
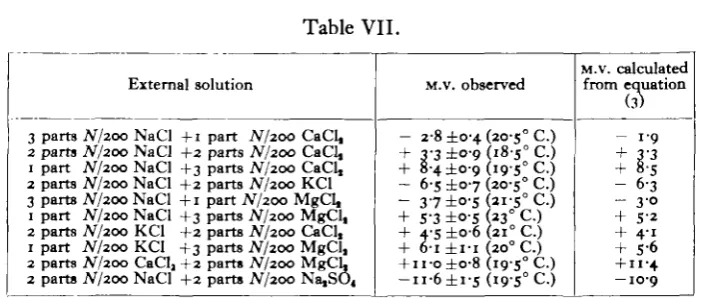
esti-mated by means of this equation agree well with those determined experimentally, as shown in Table VII. In the case of frog's skin (Hashida, 1922, p. 293) and onion scale (Pumphrey, 1932, p. 440) the relation between the potential and the composi-tion of the medium is more complex.
Membrane Potential of Paramecium
Table VII.
External solution
3 parts iV/200 NaCl +1 part N/200 CaCl, 2 parts N/200 NaCl +2 parts iV/200 CaCl, 1 part N/200 NaCl +3 parts N/200 CaCl, 2 parts iV/200 NaCl +2 parts N/200 KCl 3 parts JV/200 NaCl +1 part A7200 MgCl, 1 part JV/200 NaCl +3 parts JV/200 MgCl, 2 parts N/200 KCl +2 parts N/200 CaCl, 1 part JV/200 KCl +3 parts N/200 MgCl, 2 parts N/200 CaClj +2 parts N/200 MgCL 2 parts JV/200 NaCl +2 parts N/200 Na,SO4
M.V. observed
- 2-8 ±0-4 (20-5° C.) + 3-3 ±°-9 185° C.) + 8-4 ±0-9 (i9-5° C.) - 6-s ± 0 7 (20-5° C.) - 3 7 ±o-s 2i-s° C.) + S-3 ±0-5 (23° C.) + 4-5 ±o-6 (210 C.)
+ 6-i ± I - I (200 C.)
+ n-o±o-8 (19-5° C.) - u - 6 ± i - s ( i 9 - S ° C . )
M.v. calculated from equation
(3)
- 1 9 + 3'3 + 8-5 - 6 3 - 30 + 5-2 + 4-i + S-6
+ I I - 4
— 10-9
The sign is that of the cell interior with reference to the external surface.
As is described in the text, each component solution was buffered with a weak carbonate of its own cation.
McCleland and Peters, 1919), only a dilute solution (e.g. 3 x io~s N CeCl3 made up in NaCl) could be used, since in higher concentration the organism did not survive exposure to the solution for 2 hours before the experiment. As is clear from Table VIII, these ions, even at very low concentrations, modify the membrane potential due to NaCl and shift the value of the charge on the cell interior to the negative side, while the effect of dilution of NaCl is fundamentally the same with or without these ions. Expressing the composition of the solutions in Table VIII in the form of those in Table VII, 3 x io~8 N CeCl3 made up in N/200 NaCl (for example)
Table VIII.
External solution
3 x io-» AT CeCl, made up in buf-fered NaCl solu-tion
The same buf-fered NaCl solu-tion without LaCl, or CeCl,
8 x io-« JV LaCl, made up in buf-fered NaCl solu-tion
Concentration of NaCl solution in which La or Ce salt was dissolved
N/40
M.V.
+ 8 7 ± I - 6 ( I 8 ° C . )
+ I I - I ± I - 4 ( I 8 - 5 ° C . )
+ 67 ±o-8 (210 C.)
N/200
M.V. -i4-S±o-8(i8°C.)
- 7-1 ±0-9 (19° C.)
-14-5 ±i-6 (360 C.)
JV/1000
M.V. - 2 4 7 ±i-6 (20° C.)
-i9-8±o-8(2o°C.)
-23-3 ± 0 8 (2i-5° C.)
The sign is that of the cell interior with reference to the external surface.
in the membrane is greater than that of Na' ion. Such a conclusion, however, seems paradoxical, since the hydrated Ce'" or La"" ion is generally believed to be much larger than the Na' ion. The modification of the charge produced by the Ce"" or La"" ion must be explained in some other way; e.g. these ions may be adsorbed strongly on the external surface of the membrane, reducing its negative zeta potential, and diminishing the difference between cationic and anionic mobilities on that side of the membrane.
SUMMARY.
1. The electrical potential which exists between the cells interior and the outer surface of Paramecium when these are connected by a solution, depends largely on the nature of this solution.
2. The effect of the external />H (6-o-8-o) seems to be negligible.
3. The nature of the cation largely determines the potential observed. The effect of the anion is less marked.
4. In all solutions tested, the more dilute the solution, the more negative is the charge on the cell interior. Hence it seems probable that the membrane of
Para-mecium is more permeable to cations than to anions.
5. On mixing two solutions of equivalent normality, the potential observed in the mixture is intermediate in value between those measured in the solutions before mixing and the deviation is proportional to the ratio of the volumes mixed. Certain electrolytes (LaCl3 or CeCls) are anomalous in this respect.
6. It is shown that in wiring a Lindemann electrometer it is important to insert a reversing key between the paired electrodes and the electrometer, when using high-resistance electrodes.
I wish heartily to thank Dr James Gray, F.R.S., for the many facilities he has afforded me in his laboratory and also for his valuable advice. I am grateful to Mr L. E. R. Picken for his kind help in preparing the manuscript.
REFERENCES.
ETTISCH, G. and PffrtRFl, T. (1925). Pflilgert Arch. 208, 454. GKUAN, S. (1927). Univ. Calif. Publ. Zool. 29, 453. HASHIDA, K. (1922). Journ. Biochemistry, 1, 289.
MCCLELAND, M. and PFTBRS, R. A. (1919). Journ. Phytiol. 53, 12 P. PACKARD, C. (1925). Journ. Gen. Pkytiol. 7, 363.
PUMPHREY, R. J. (1931). Proc. Roy. Soc. Lond. B, 108, 511. (1932). Proc. Roy. Soc. Lond. B, 109, 434.