F
u n c t i o n a lO
r g a n i z a t i o nO
fT
h eM
o n k e yV
i s u a lC
o r t e xF
o rS
t e r e o s c o p i cD
e p t h.
Daniel Lewis Adams.
PhD. Thesis.
University College London.
All rights reserved
INFORMATION TO ALL USERS
The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest 10017388
Published by ProQuest LLC(2016). Copyright of the Dissertation is held by the Author.
All rights reserved.
This work is protected against unauthorized copying under Title 17, United States Code. Microform Edition © ProQuest LLC.
ProQuest LLC
789 East Eisenhower Parkway P.O. Box 1346
Contents
A b stract...5
A cknow ledgem ents...7
Introduction Sectionl The perception of depth Historical review of stereoscopic vision...9
The m eeting of projections from the e y es...15
The random dot stereo g ram ...17
Local and global stereo p sis...18
Psychoanatom y...19
Ocular dom inance...20
Ocular interaction...22
Disparity tu n in g ...23
The functions of different types of disparity tuned cell...30
O ther types of d isp arity ... 31
Psychophysics...34
The correspondence p ro b lem ... 37
Colour and stereo p sis... 37
Isoluminance studies... 38
Spatial frequency...40
O rientation and stereo p sis... 41
Stereoscopic depth constancy... 42
Com putational m o d e ls... 44
Stereoblindness... 47
Prim ate lesion stu d ie s... 50
Effects of abnormal visual experience on binocular v isio n ... 51
Section 2 The physiology of the V3 complex Orientation tuning in the V3 com plex... 55
Colour cells in the V3 com plex... 56
Real-motion cells... 58
Gaze dependent c e lls... 59
Section 3 A natom y of stereopsis Anatomical description of the prestriate a re a s ... 62
Cytochrome oxidase divisions w ithin the visual cortex... 64
"What" and "where" p a th w a y s... 67
Materials and m ethods Pharm acology... 73
S u rgery... 75
P erfusion... 76
O ptics... 77
Physiology... 78
Categorization of cell properties Receptive field data statistics... 84
Receptive field characteristics... 85
Response characteristics... 86
Anatomical tracing and histological p ro ced u res...90 Histology...92 Cortical reconstructions...95 Results Section 1 Electrophysiological data Allocation of cells to visual a re as...103
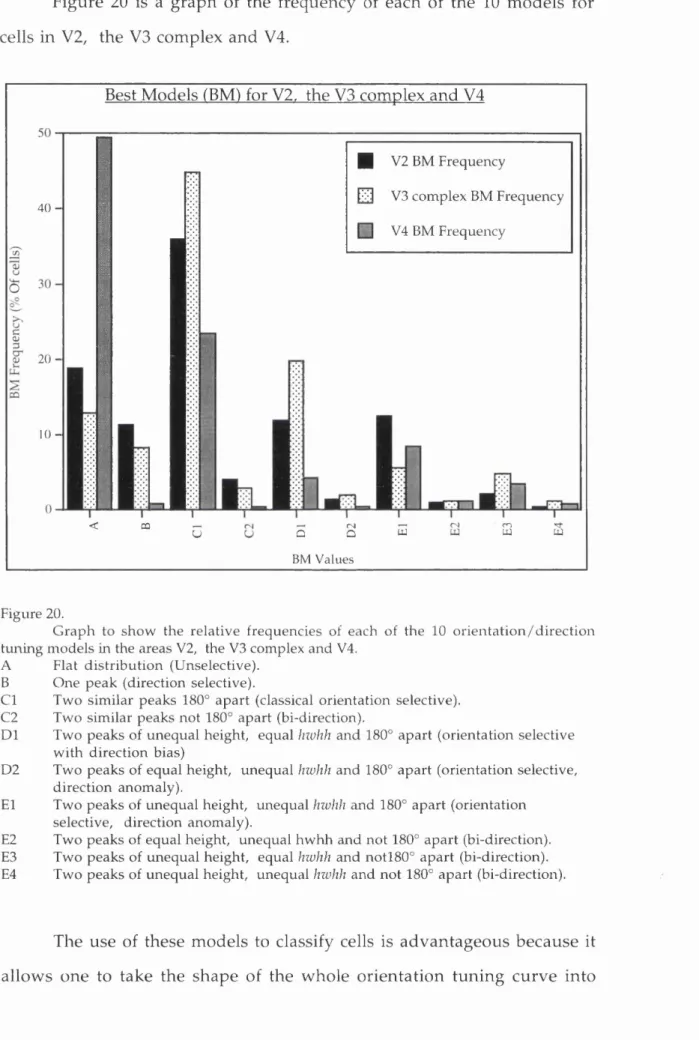
The properties of cells in the V3 com plex...105
a) Orientation selectivity tu n in g ...105
b) Direction selectivity...I l l c) W avelength selectivity...120
d) Clustering of cells w ith similar p ro p erties...123
e) Binocular properties...126
f) Disparity tu n in g ...130
Proportions of disparity cells...139
Properties of disparity tuned c ells...140
Orientation and direction tu n in g ...142
Length tu n in g ...144
Receptive field eccentricity...144
Vertical d isp a rity ...146
Functional organization of disparity tuned cells...149
Com parison w ith V 2 ...169
Section 2 Anatomical Data Introduction... 174
Description of the visual a re a s ...175
Injection sites... 176
Cortical projections of V3... 177
The projections of V 3 A ... 185
Com parison of V3 and V3A... 197
The projections of DP... 198
Discussion Section 1 Physiology Introduction... 208
Orientation and direction... 209
Non-classical orientation/direction cells...209
Com parison of V2 and the V3 com plex...210
O rientation and d isp arity ... 213
Topography of the V3 com plex... 214
Disparity selective cells in the prestriate areas... 217
Proportions of disparity tuned cell ty p es... 219
Vertical disparity selectivity... 220
U northodox disparity cells... 222
Quantitative depth selectivities... 224
Motor and sensory system s... 225
Form vision... 226 Section 2
Introduction...228
Pathw ays in the visual co rtex ...228
Connections of the V3 com plex...231
"What" and "Where" path w ay s...233
Oculomotor sy ste m s...234
Parietal corm ections...235
Patchiness of connections...237
General conclusions...238
A ppendix A General model of cell response to different stim ulus directions Description of the m o d e ls...240
Model optim isation...241
Statistical analysis of m o d e ls...245
Abstract
O ur own, and other previous studies of V3 and V3A (two visual areas
constituting the V3 complex) have show n it to contain m any orientation
tuned cells w ith strong binocular interactions (Zeki 1978b; Burkhalter and
Van Essen 1986; Felleman and Van Essen 1987). This, and the area's M
dom inated cortical input, led us to choose the V3 com plex as a likely
candidate for an area specialized in the processing of stereoscopic depth.
Thus, the aim of this study was to record from single cells in V3 and V3A
and determ ine their selectivity for stereoscopic depth. The responses of cells
in these tw o areas w ere exam ined and the distribution of disparity tuned
cells was com pared w ith that in area V2, which has all m odalities of vision
represented in it. The results show that the V3 complex contains a high
percentage of disparity tuned cells and th at these are also orientation or
direction tuned. Thus, these cells are detecting the horizontal disparity of
visual features. Previously defined classes of disparity tuned cell (Poggio
and Fischer 1977; Poggio et ah 1988) w ere found in both the V3 complex and parts of V2. In agreem ent w ith previous studies, (Burkhalter and Van Essen
1986; H ubei and Livingstone 1987) disparity tuned cells w ere found in the
thick stripes of V2.
The second p a rt of this stu d y was to examine brains injected w ith
anatom ical tracing agents to discover the cortical connections of the V3
complex and an area on the dorsal prelunate gyrus (DP) w hich receives an
input from V3A. Results show ed that the areas injected V3, V3A and DP
have extensive connections w ith each other and w ith areas dealing w ith
other attributes of vision. The results dem onstrate th at the V3 com plex
areas w hich use this inform ation for the analysis of m ore complex visual
Acknowledgements
This piece of work w ould not have been possible w ithout the help of a
num ber of people, primarily, my supervisor Professor Zeki, whose w isdom
encouragem ent and advice was invaluable. I w ould also like to express my
gratitude to Dr. Stuart Shipp for sharing his enthusiasm and expertise during
experim ental w ork as well as his critical reading of the m anuscript. G rant
W ray and John Romaya for technical and com puter assistance and M ark
Rayan and Anne Fitzpatrick for histological tutoring and help. I w ould also
like to thank m y colleagues, Konstantinos M outoussis, A ndreas Bork and
Ludovica Marini for being good company and sharing the work load during
experiments that usually lasted 24 hours a day for an entire week.
I am also grateful to the BBSRC, W ellcome Trust and latterly the
D epartm ent of Social Security for providing the funds necessary for the
In trod u ction
Sectionl
The perception of depth
The ability to judge depth is an im portant faculty of visual animals;
it provides them w ith know ledge of distance w ith respect to the organism
and distances of objects w ith respect to one another and hence prepares
them for m aking a p p ro p riate m otor responses. It is of ev o lu tio n ary
im portance because it greatly aids the hunting and capture of prey as well
as the safe navigation of anim als in their environm ent. Psychophysical
studies indicate th at the perceptual transform ation from tw o to three
dim ensions relies on tw o types of cues: cues for m onocular depth, and
stereoscopic cues for binocular disparity. A t distances greater than about
50m the retinal images from each eye are virtually identical, so d ep th
m u st be judged using m onocular cues. There are at least five types of
m onocular d ep th cue:
1. Previous familiarity. If w e know from experience som ething about the size of an object we can judge the object's distance.
2. Interposition. If one object is partially h id d en from view by another object we assum e the hidden object is further away.
3. Linear and size perspectives. Parallel lines ap p ear to converge w ith distance. The greater the convergence of lines, the greater the im pression
of distance. The visual system interprets the convergence as d ep th by
assum ing th at the lines rem ain parallel.
4. Distribution o f shadows and illumination. Patterns of light and d ark can give the im pression of d ep th . The sh ad in g on objects im plies
different inclinations to the light source and on the w hole, b rig h ter
5. Motion for monocular movement) parallax. As w e m ove ou r h e ad s from side to side, the image projected by an object in the visual field
moves across the retina. N earby objects seem to m ove quickly and in the
o p p o site direction to our m ovem ent, w hereas d ista n t objects m ove
slowly.
All of the above cues (except the fifth) are used by artists to endow
their paintings w ith a sense of depth. This im pression of d ep th can be so
co nvincing to the b ra in th a t if som e p ain tin g s (p articu larly th o se
c o n ta in in g a g rea t deal of lin e ar p e rsp e c tiv e cues) are v ie w e d
m onocularly, the visual system will m ake convergent and divergent eye
m ovem ents that m atch those th at w ould be m ade w hen view ing the real
three dim ensional scene (Enright 1987).
Objects view ed at distances sm aller th an ab o u t 50m project a
slightly different image onto each retina because the eyes are separated
horizontally. This difference (called disparity) can be used by the visual
system as a pow erful d ep th cue. The analysis of d isp arity to p roduce
d epth vision is called stereopsis.
Historical review of stereoscopic vision
The history of ideas about vision is docum ented back to the tim e of
the Greeks, w ho, follow ing an idea by Em pedocles (5th century BC),
believed th at light leaves the eye in the form of a cone m ade u p of
straight rays and these rays gain know ledge of the w o rld b y "feeling"
objects rath er like an invisible hand. The inform ation g ain ed is then
tran sp o rte d back to the eye and b rain to generate visu al sensations.
Recently it has been com m ented th at this idea of active v ision is m ore
The first breakthrough in visual optics was the w ork of an Arabic
scholar by the nam e of A lhazen (965-1040). H e firm ly rejected the
em anation theory of vision proposed by the Greeks and in his book "The
Book of Optics" (Kitab al-manazir) (Alhazen 1989). He proposed that the
visual scene is projected as a two dim ensional inverted im age onto the
retina. Realising th at the im age w ould be inverted and reversed, he
suggested th at it was corrected by a further refraction from the back of the
eye to form a corrected im age on the optic disc. M any years later,
Johannes K epler (1571-1630) concluded th at there w as no basis for the
second inversion proposed by Alhazen and suggested th at the image w as
rectified by a m ental process. The realization that the visual image w as
projected onto the retina as a tw o dim ensional rep resen tatio n caused
people to w onder how the flat image on the retina was transform ed into a
three dim ensional p recept of the visual w orld. This problem w as of
p articu lar concern to artists, w hose m ain preoccupation w as how to
rep resen t three dim ensions on a flat canvas. Since stereoscopic cues
cannot be used in paintings, the foci of these enquiries w ere based on
m onocular depth cues.
A lthough the rudim ents of binocular vision had been observed by
Euclid, w ho noticed that the eyes each have a slightly different view of
the w orld, and Aristotle, w ho noted that w hen one eye is pressed w ith a
finger, a double image is produced, binocular vision was not equated
w ith d e p th perception. Thus the question arose "why do w e have tw o
eyes?" For m any years (since the tim e of Alhazen) it w as believed th at
two eyes w ere useful because if one becomes dam aged, the other rem ains
intact. This is not to say th at the geom etry of binocular optics w as not
studied; A lhazen him self described the w ay in w hich objects placed in
that lie at, or close to, the fixation plane, i.e. on corresponding points,
are fused to form a single image. W ith this observation, A lhazen
described w hat w e now know as the horopter, i.e. the plane in space at
w hich objects can be seen as single fused images. A lthough A lhazen
realized that this plane w as not fronto-parallel, its precise geom etry w as
n o t stu d ied u n til 1818 by Vieth and M üller (Vieth 1818; M üller 1826).
Sim ilarly, A lhazen described a sm all area in space either side of the
fixation plane, at w hich objects can still be seen as single and fused, this
area is now know n as Panum 's fusional area, and its study had to w ait
until 1858 (Fanum 1858).
P erhaps the earliest description of stereopsis w as p ro d u ce d by
Leonado da Vinci (1452-1519); in his w ork Trattato della pittura (see Keele 1955; Strong 1979) he observed; "That a p a in tin g , th o u g h
conducted w ith the greatest art and finished to the last perfection, both
w ith regard to its contours, its lights, its shadow s and its colours, can
never show a relievo equal to that of the natural object, unless these be
view ed at a distance and w ith a single eye." taken from W heatstone
(1838). H ere Leonado im plied that tw o dim ensional pictures can only
truly p o rtray the real scene if that scene is devoid of stereoscopic d ep th
cues. H ow ever, Leonado did not show a full u n derstanding of the finer
points of stereopsis other than the idea that closing one eye transform ed
the visual w orld into an image m ore resem bling that on a canvas.
In 1613 Franciscus A guilon published his w orks on optics entitled
Opticorum libri sex. His 6 books followed the w ork of Euclid, A lhazen and others w ith the ad d en d u m that d ep th perception is im proved w ith
binocular vision. This point w as illustrated in A guilon's book w ith an
estimating the distance to an object held in front of him (reproduced in
figure 1).
Figure 1.
Rubens' engraving depicting a one-eyed man underestim ating the distance to an object held in front of him to portray the point that binocular vision is im portant for depth discrim ination. Produced in 1613, it w as com m issioned by Franciscus A guilon to illustrate his work O pticorum libri sex (Taken from Judson and van de V elde 1978).
Thus, the question "Why do we have two eyes?" was answered
nearly two centuries after it was asked. Aguilon studied the geometry of
binocular vision using the theorem s of Euclid and coined the term
"horopter" to describe the plane in space at which both fused and diplopic
images appear to lie. The word horopter comes from the Greek "horos", meaning space and "opter”, observer. The word horopter is still used today, but to describe a slightly different plane. Aguilon used the term to
describe a fronto-parallel plane at the same distance from the observer as
the fixation point. Today the horopter describes a plane in space, points
fronto-parallel plane, this produces a horizontal circle (the Vieth-Müller circle)
which passes through the centres of both eyes (see figure 2).
Eyes'
V ieth -M ü ller circle (H orop ter)
Figure 2.
I llu s t r a t io n o f th e V ieth-M üller circle or horopter. T h e f ig u r e r e p r e s e n t s a horizontal plane in clu d in g both ey e s and a fixation p oin t (F). Points A and B lie on the Vieth-M üller circle. From E uclidean geom etry, angles a = a' and b = b'. Thus the points A and B both project to corresponding retinal p o in ts and are see n as sin g le fu sed im ages, as do all points that lie on the circle.
The Vieth-Müller circle was first described in a paper by Vieth
(1818), although it was simultaneously studied by Müller (1826). They
showed that all points projecting to corresponding retinal points form a
circle that passes through the optic centres of both eyes. Vieth mistakenly
generalized his theory of corresponding points in three dimensions to a
sphere. His mistake was corrected by Prévost (1843) who showed that the
theoretical horopter is in fact a toroid formed by sweeping the Vieth-
Müller circle through the interocular axis.
The notion that binocular vision gives rise to depth perception was
not further pursued until the 19th century w hen Charles W heatstone
(the inventor of the W heatstone bridge) published a paper entitled:
remarkable, and hitherto unobserved. Phenomena of Binocular Vision"
(Wheatstone 1838). He showed an understanding of stereoscopic depth
perception and illustrated the phenom enon w ith the invention of the
stereoscope. With this instrument, shown in Figure 3, two photographs
of a scene taken 60-65mm apart, one from the position of each eye, are
m ounted onto a binocular like device such that the right eye sees only the
picture taken from the right position and the left eye sees only the left
picture. Fusion of the two images produces a three dimensional image.
Figure 3.
A diagram of W heatstone's stereoscopic apparatus. The tw o m onocular im ages (E' and E) are reflected by mirrors (A' and A) placed close to the eyes of the view er, so that each eye only sees one im age. The tw o im ages are therefore superim posed and the view er fuses them to produce a "solid" stereo image. (Taken from W heatstone 1838).
Thus the stereoscope vividly demonstrates stereopsis by enabling
convincing stereoscopic depth to be perceived from tw o-dim ensional
images. In W heatstone's own w ords, "The preceding experim ents
render it evident that there is an essential difference in the appearance of
objects w hen seen with two eyes, and when only one eye is employed,
and that the m ost vivid belief of the solidity of an object of three
dimensions arises from two different perspective projections of it being
d isp arity in W heatstone's stereoscopic pictures are exam ined it can be
seen th at corresponding elem ents in each picture are shifted tow ards or
aw ay from each other; the form er causes a shift in d ep th in front of, and
the latter behind, the view er's fixation plane.
In 1858, Panum described a range of disparities w ithin w hich tw o
sim ilar images, one in each eye, are perceived as a single image (Panum
1858). This plane (Panum 's fusional area) is an area in d ep th that extends
in front of and behind the fixation plane that corresponds to the region of
b in o cu lar single vision. The im ages from the tw o eyes of all p o in ts
w ithin this area are said to be 'fused'. The front to back size of the area in
d ep th is d ependent upon a num ber of factors: it increases in a geometric
w ay as the fixation plane becom e fu rth er aw ay from the eyes b u t the
visual angle subtended from the front of the area to the back rem ains
constant, and it tends to increase in size tow ards the p eriphery of the
visual field. It is also d ependent on the n atu re of the stim ulus and its
background (Panum 1858; Ogle 1964).
The m eeting of projections from the eyes
In order th at the tw o view s of the w orld be unified to p roduce a
single percept, inform ation from both eyes m ust arrive at single cells in
the brain. A lthough this concept seems obvious today, for a long tim e it
w as th o u g h t th at the m onocular images rem ained separate th ro u g h o u t
the visual system and w ere combined "by a m ental act" (Helm holtz 1893).
Ram on y Cajal stood alone in his belief that inputs from corresponding
parts of the tw o retinae converge on w hat he called "isodynamic cells" and
it is this th at forms the basis of unified binocular vision (Ramon y Cajal
1911). The location of the first binocular cells in the visual system has
binocular vision m ay be elucidated. N ot unreasonably, Galen (175) and
after him , A lh azen (1989), th o u g h t th a t the locus of b in o cu lar
com bination w as the optic chiasm . D uring the R enaissance, René
D escartes p roposed th at the optic fibres m ight converge on the pineal
gland for unification (Traité de l'Homme). In fact the first possible site for unification of inform ation from the tw o eyes is the lateral geniculate
nucleus (LGN), w here fibres from both eyes term inate having already
been com bined en-passage th ro u g h the optic chiasm . The LGN is a
n u c le u s c o m p risin g six lay ers. P h y sio lo g ica l a n d a n a to m ic a l
exam inations of the layers (Silva 1956) reveals them to contain cells that
receive m onocular inputs; layers 1, 4 and 6 receive projections from the
c o n tralateral eye and the oth ers are exclusively in n erv a te d by the
ipsilateral eye. Cells in these layers are also solely excited by their
respective eye. Extensive connections exist betw een the layers of the LGN
b u t these do n o t endow the cells w ith the p ro p e rty of b in o cu larity
(G uillery 1971). Thus, fibres leaving the LGN still carry m onocular
inform ation. The next stage in the visual p athw ay is the prim ary visual
cortex (striate cortex, area 17 or VI), more specifically layer 4c, the in p u t
layer. In 1959, H ubei and Wiesel proved Ramon y Cajal to be correct by
d e m o n stra tin g the existence of binocular cells in the cortex (Cajal's
"isodynamic cells"). These cells first occur in VI except in layer 4c. V I is
therefore the first locus w here fusion of the tw o m onocular im ages can
occur (H ubei and Wiesel 1959). Quite w hy cells retain their exclusive
m onocularity until this stage, w hen they have h ad am ple o p p o rtu n ity
for generation of binocularity at previous parts of the visual system , is
The random dot stereogram
H ow does the brain know w hich p a rts of the m onocular im age
correspond to the sam e object view ed by the other eye? This question
(the co rresp o n d en ce problem , w h ich w e w ill re tu rn to) rem ain s
unresolved today b u t a great advance w as m ade in 1960. Bela Julesz, a
ra d a r engineer, w as w orking w ith w artim e aerial p h o to g rap h s taken
b eh in d enem y lines. A technique used to spot cam ouflaged structures
w as to take tw o photographs from slightly different locations and view
them w ith a stereoscope this w ay objects taller th an their surro u n d in g s
stand o u t in binocular depth. At the time, the prevalent view about the
correspondence problem held by psychologists w as th at the b rain first
recognized an object in the m onocular images, and then paired the tw o
objects to produce single vision and stereoscopic depth. W orking w ith
stereoscopic images com prising few recognizable forms, Julesz knew this
n o t to be the case. To dem onstrate his observation he m ade images w ith
a com puter, sim ilar to his stereoscopic aerial photographs b u t containing
no recognizable forms. These "random dot stereogram s" (RDS) com prise
tw o random , yet correlated patterns of dots w ith a central portion of dots
being shifted by an integer m ultiple of the dot size in opposite directions
for each im age (Julesz 1960). Fusion of the tw o im ages p ro d u ces a
pow erful sensation of depth, the central portion being displaced in front
of, or behind the surrounding texture, depending on the direction of the
Figure 4.
An example of Julesz's original random dot stereogram. Diverging the eyes in such a way as to superimpose and fuse each monocular image produces the sensation of depth. In this figure, divergent viewing produces a smaller square displaced behind the plane of the paper.
These figures v iv id ly d em o n strate th a t, c o n tra ry to th e
p red o m in an t theory of the day, binocular d ep th perception does not
require m onocular form recognition and is an operation carried out quite
"early" in the visual system. This revelation shifted the em phasis of
n eu ro p h y sio lo g ical research from p ro b lem s in m o n o cu lar form
recognition to the search for binocular cells in the visual brain. It also
introduced the notion of local and global stereopsis.
Local and global stereopsis
One of the m ost im portant consequences of the invention of the
random dot stereogram was the revelation that m onocular form or d ep th
cues are not necessary for binocular fusion. Thus, the visual system
m u st be using a very m uch m ore sim ple m ethod of fusing binocular
images than previously supposed. Since the m any similar elem ents in a
random dot stereogram could easily give rise to false matches, the system
m ust be using m ore than just the form of the individual elem ents to
produce binocular fusion. Thus, global stereopsis is the m ethod the
visual system uses to examine large portions of the m onocular images
a n d u n d e rta k e s a cross correlation analysis, n o t only m atch in g
n eig h b o u rh o o d to disam biguate false m atches (Julesz 1971; Julesz and
O sw ald 1978). Local stereopsis refers to the m ore classical kind of
binocular fusion w here, w ithin Panum 's fusional area, unique features
in each m onocular im age are paired to resolve am biguity (Julesz 1971;
Julesz and O swald 1978).
Psychoanatom y
Since the d ep th features of random d o t stereogram s can only be
seen w hen m onocular inform ation from each eye has converged onto a
single cell, it can be said th at all the n eu ral m achinery u p to V I is
incapable of responding to stereoscopic depth. Thus, any perceptual
effects produced by view ing stimuli presented as random dot stereogram s
m u st be p ro d u ced in the cortex rath er th an the retina or LGN. The
lo c a liz a tio n of p e rc e p tu a l effect u sin g p sy c h o p h y sic s w as called
psychoanatom y (Julesz 1971). M any types of ran d o m dot stereogram s
(both static and dynam ic) have since been p ro d u ced , each designed to
exhibit a particular optical illusion or perceptual aftereffect. These studies
are n u m ero u s and m ostly draw the sam e conclusions, i.e. p ercep tu al
effects can be reproduced w ithout the need for m onocular features and
m u st th erefo re be p ro d u c e d w ith in th e v isu a l cortex. The first
phychophysicist to em ploy disparity instead of lum inance gradients for
the presentation of optical illusions w as Papert (1961). By presentation of
com m on optical illusions, such as the M üller-Lyer lines^, Papert show ed
th at the illusions p ersist w hen no m onocular cues are p resen t, th u s
dem onstrating that the locus of the illusion is cortical rather than retinal
or thalam ic.
O cular dom inance
The advent of single unit recording bro u g h t about a revolution in
the study of the visual system. First applied in the prim ary visual cortex,
cells w ere fo u n d th at resp o n d ed to stim uli p resen ted to the anim al
(H ubei and Wiesel 1959). Cats w ere used at first, and later, because of
their superior vision (especially colour vision) and the sim ilarity of their
visual system to hum ans, m onkeys w ere recorded from. The relative
influence of the two eyes on the response of a cell (ocular dom inance)
varies from cell to cell in VI and a scheme of classification w as devised
(H ubei and Wiesel 1962). The stim uli used to categorize cells into these
groups w ere presented to each eye in turn. Thus, it was not possible to
find out how the responses of the cell m ight change w hen both eyes are
stim u la te d sim ultaneously (binocular interaction). Cells in V I w ere
found to fit into one of seven groups: group 1 contains cells th at are
exclusively contralaterally driven, w hile group 7 cells are exclusively
ipsilaterally driven, w ith all gradations in between.
C ells in th ese g ro u p s w ere fo u n d to fit into a c o lu m n a r
o rg a n iz a tio n in the p rim a ry v isu al cortex, the ocular d o m in an ce
colum ns. These robustly organized colum ns are only p resen t in the
p rim ary visual cortex and com prise a patchw ork of m odules norm al to
the surface of th at area, each eye supplying irregular stripes of cortex
about half a m illim etre wide. Ocular dom inance colum ns rem ain one of
the m o st solid fin d in g s in the v isu al system and have since been
d em o n strated anatom ically and physiologically. The first anatom ical
dem o n stratio n w as achieved by m aking lesions in single layers of the
LGN an d looking at the p attern of degenerated axon term inals in the
striate cortex (H ubei and W iesel 1969). Since the layers of the LGN
m onocular ocular dom inance colum ns. Technical advances led to an
im p ro v e d anatom ical m eth o d of v isu alizatio n of ocular dom in an ce
colum ns involving the injection of a radio-labelled am ino-acid (tritiated
proline) into one eye. It w as first achieved in the m ouse (Grafstein and
Laureno 1973) and later em ulated in the m onkey (Wiesel et ah 1974). The am ino-acid is transported from the retinal ganglion cells to the LGN and
th en transneuronally to striate cortex. A utoradiography of sections taken
from striate cortex exhibit patches of dense transport of the amino acid in
layer 4, corresponding to the dom ains that contain m onocular cells. The
2 D eoxy-glucose (2DG) technique (Sokoloff 1977), w hen em ployed by
T ootell et al. (1988a), also dem onstrated, am ong other things, the p attern of ocular dom inance columns in the striate cortex of the m acaque.
The 2DG technique is a m ethod of show ing a p ictu re of functional
activity; radioactive glucose (^4(]_2_deoxy-d-glucose) is injected into the
anim al's circulation w hile it is presented w ith a visual stim uli. A fter
view ing the stim uli for a few m inutes in the paralysed and anaesthetized
state (to p rev e n t eye m ovem ents) the anim al is killed and its b rain
sectioned. Areas that were very active directly before the anim al's death
co n tain m ore of the radioactive glucose an d can be v isu alized by
autoradiography. Thus, full field m onocular stim ulation of the anim al
w ill p roduce pictures of the ocular dom inance colum ns in flat m ounted
sections, cut parallel to V i's surface. These techniques have established
th at the ocular dom inance colum ns form a series of parallel bands, each
0.25 to 0.5 m m wide. They spread through all layers of the cortex and in
each hem isphere a retinotopic m ap of the visual hem ifield is represented
twice, one w ithin the left eye colum ns and one w ith in the right. The
b ands tend to ru n 90° to the V 1/V2 border in iso-eccentric stripes', thus
m apping tw o circular visual fields onto the elliptical surface of the striate
H ubei and W iesel suggested th at the differing degrees of ocular
dom inance m ight, in some way, provide the basis for a depth sensitivity
m echanism , the dom inance characteristics being segregated so th at each
m onocular com ponent of an im age could be com pared to th at of the
other eye. This has so far proved not to be the case, and the functional
significance (if any) of ocular dom inance and its colum nar organization
in V I in som e (but not all^) anim als w ith stereoscopic vision rem ains
unclear.
O cular interaction
A side from H ubei and W iesel's ocular dom inance characteristics,
another type of binocular pro p erty exists for cells in the visual cortex.
Since H ubei and Wiesel stim ulated their cells m onocularly, they did not
find inform ation relating to the binocular interactions of cells. W hen
cells are d riv en binocularly their responses m ay change and are n o t
predictable from the m onocular responses. Zeki (1979) exam ined this
p ro p e rty an d d ev ised a m eth o d of classification of b in o cu lar cells
d e p en d in g u p o n their response d u rin g binocular stim ulation. Thus,
cells w ere divided into six groups according to w hether they w ere driven
by one eye only (two categories), dom inated by one eye (two categories),
e q u a lly w ell d riv e n by e ith e r eye (one categ o ry ) or d riv e n by
sim u ltan eo u s binocular stim ulation only (one category). U sing these
categories, so called "ocular interaction histogram s" could be plotted for
p o p u latio n s of cells. W hen these histogram s are p lo tted for p articu lar
visual areas one can see th at nearly all cells in prestriate cortex, i.e. all
cells beyond V I, are (i) binocular and (ii) produce differing degrees of
bin o cu lar interaction. Thus all these p restriate areas are, in theory.
candidates for analysing stereoscopic depth. However, since disparity was
n o t systematically varied in Zeki's study, cells that only respond at critical
disparities m ay have been missed. It is this property that enables cells to
be d ep th selective, and an area w ith a high proportion of these disparity
selective cells w ould be a far m ore likely candidate for an area of d ep th
specialization.
D isparity tuning
The receptive fields of binocular cells in VI w ere found to occupy
corresponding areas on the tw o retinæ and their response properties w ere
qualitatively identical for both eyes (Hubei and Wiesel 1962). Stereoscopic
vision requires m ore than the presence of binocular cells alone. In order
to get inform ation reg ard in g the relative d ep th s of objects from the
fixation plane the system needs to be capable of fusing the images from
b o th eyes an d an aly sin g the in fo rm atio n d eriv e d from th e sm all
differences betw een each image. To achieve this one w ould expect some
b in o c u la r cells to h av e rec ep tiv e field s th a t are slig h tly n o n
corresponding, this receptive field disparity being in the order of a few
m inutes of arc, thus allowing the cells to respond maxim ally w hen slight
disparities exist betw een the two m onocular images. This is w hat m akes
cells disparity tuned.
T he e x p e rim e n ta l s tim u la tio n of b o th re c e p tiv e fie ld s
sim u ltan eo u sly w ill yield inform ation reg ard in g binocular interaction
(facilitation or attenuation), b u t in order to stim ulate cells at specific
disp arities a m ore elaborate regim e is required. One solution in the
paralysed anim al is to superim pose both receptive fields on a screen using
q u a n tita tiv e ly v aried to stim u late the cell over a ran g e of retin al
disparities. A nother m ethod is to leave the receptive fields separated and
em ploy tw o stimuli, one for each receptive field (Henry et ah 1969). If the tw o stim uli are correctly positioned, the anim al should fuse them and its
visual system be presented w ith a single binocular stim ulus. By varying
the distance betw een the tw o stim uli one can change the disparity. The
advantage of both of these techniques is that one can stim ulate cells at
specific disparities and if a cell's responses are sensitive to small changes
in disparity, it shows that the cell is tuned to stim uli placed at a specific
distance relative to the horopter.
The m ethod of superim posing the receptive fields of a cell using
prism s w as em ployed by Pettigrew et al . (1968) in the striate cortex of the cat. Their m ajor finding w as th at cells had greatly facilitated responses
w h en stim u lated by a single m oving bar w ith th eir receptive fields
superim posed; furtherm ore, some cells were exquisitely sensitive to the
exact position of the receptive fields. Differences as small as 3 m inutes of
arc w ould greatly m odify the cell's responses, and if disparity (i.e. the
prism setting) w as set to non optim al values, some cells' responses could
be inhibited. Thus cells in cat striate cortex can be selective for disparity.
If the d isp arities these cells are tu n ed to deviate from zero one can
assum e they are doing m ore than fusing the tw o retinal im ages to form
the "cyclopean im age"^. A p o p u latio n of cells w hose responses are
m axim ized over differing disparities w ould in effect be tuned to different
positions in d ep th relative to the horopter.
In order to m easure differences in disparity tuning from cell to cell,
the very sm all eye m ovem ents that occur, even after complete paralysis
of the eye m usculature, m ust be stopped, or m easured and subtracted
fro m th e d ata. The m eth o d s of eye m o v em en t e lim in a tio n in
B ishop/P ettigrew 's group were to plot the optic disks of each eye onto a
screen and m onitor their m ovem ent. This m ethod is only possible in the
cat because it, unlike the monkey, has reflective retinae; shining a bright
light into the cat's eyes produces an image of its retinae on the tangent
screen. A dditionally, the eyes w ere im m obilized by gluing the scleral
m argins to brass rings. W ith these m easures in place the authors found a
spread of optim al disparities of ±1° (Nikara et al. 1968), m ost cells being tuned to points at or very close to zero disparity, i.e. the horopter.
U sing the prism m ethod of aligning receptive fields, H ubei and
Wiesel w ere also looking for binocular cells (Hubei and Wiesel 1970), b u t
in the m onkey visual cortex. In order to keep track of eye m ovem ents
they sim ultaneously recorded from a binocular V I "reference" cell in the
opposite hem isphere and repeatedly plotted its receptive fields, taking
note of any m ovem ents. Since the receptive fields of VI cells are sm all
and well defined, any m ovem ent of the positions of the reference cell's
receptive fields will correspond to eye m ovem ents, and can be rem oved
from the data obtained from the investigating electrode. Unable to find
binocular d ep th cells in area 17 (VI) they m oved onto the territory of the
prestriate cortex w hich at the time w as thought to comprise tw o areas (18
an d 19). H ere they found a large p ro p o rtio n of cells (43%) w hose
responses w ere critically d e p en d en t u p o n the prism setting and w ere
therefore disparity selective or "binocular depth cells". H ubei and Wiesel
n o ted th at although there w as variation in horizontal d isp arity am ong
v ary in g orientation selectivities. These groups of binocular d ep th cells
w ere segregated from cells not concerned w ith d e p th and tentatively
called d ep th columns. Hubei and Wiesel also noted that the disparity of a
cell w as related to its orientation selectivity, "The displacem ent of the
field in one eye, relative to the field in the other, is usually at right
angles to the receptive field orientation." (Hubei and Wiesel 1970). Thus
o n ly cells w ith n e ar vertical o rien tatio n tu n in g w ill be tu n e d to
h orizontal disparity. Retrospective exam ination of this paper show s th at
the au th o rs w ere recording from tw o areas, V2 and V3A; they found
b in o cu lar d e p th cells in both, alth o u g h they w ere u n aw are of this
subdivision w ithin the lunate sulcus at the time.
The sp read of optim al d isparities found by B ish o p /P ettig rew 's
g roup (Barlow et ah 1967; Nikara et ah 1968) in cat striate cortex prom pted H u b ei and W iesel to re-exam ine the stereoscopic m echanism s in th at
anim al (Hubei and W iesel 1973). H ubei and W iesel's recordings from
m o nkey striate cortex h ad show n there to be very little v ariatio n in
receptive field disparities in VI; the great m ajority of cells had their fields
in precise registration (only 5% show ing detectable disparities) and they
h ad found no convincing exam ples of binocular d ep th cells (H ubei and
W iesel 1970). In order to find out if the difference w as a genuine species
difference they recorded from the cat's striate cortex, controlling for eye
m ovem ents by keeping a striate reference cell in the opposite hem isphere
w ith w hich to com pare disparities. C ontrary to B ishop/P ettigrew 's data
they fo u n d very little variation in disparities and concluded th at the
m ain m echanism s subserving stereoscopic depth perception in the cat (as
in the m onkey) lie outside the striate cortex. The discrepancy betw een
these tw o groups is probably dow n to their stim ulation and field plotting
b e tw ee n th e receptive fields of a cell (taking eye m ovem ents into
account), b u t B ishop/P ettigrew 's group defined it as the prism setting
required to produce a maximal response of a cell. It is possible that a cell
m ay still n o t give a maximal response w hen its receptive fields have been
perfectly superim posed. Rodieck recognized this problem in term inology
(Rodieck 1971) and suggested th at the term "incongruity" be used for
receptive field d isp arity (i.e. non-corresponding locations of receptive
fields), keeping the term "disparity" for response-defined disparity only.
Since these early studies m any disparity tuned cells have been found in
cat and m onkey striate cortex (Nelson et al. 1977; Foggio and Fischer 1977; Von der H eydt et al. 1978).
Once isolated, the best way to study a binocular depth cell is to plot
a grap h of horizontal stim ulus disparity against the cell's response to that
stim ulus. For non-disparity tuned binocular cells such a graph will give
rise to a bro ad curve, show ing sim ply th at the cell responds better to
b in o cu lar stim u latio n th an m onocular. For a sm aller p ro p o rtio n of
binocular cells the curve will be a far more specific shape, show ing that
the cell is disparity tuned. D isparity tuned cells m odulate their responses
over a very narrow range of disparities and the type of m odulation they
show fits them into a schem e of classification devised by Foggio and
Fischer (1977). This classification scheme divides cells into tw o groups,
eith er n e a r /f a r (asym m etric) or tuned. Figure 5 show s the response
properties (in the form of disparity tuning curves) for their four types of
F ixation p lan e
E yes
TE
Far
N ear
D isp arity
Figure 5.
S h a p e s o f th e d is p a r it y -response curves for the four types of disparity tuned cell described by Foggio and Fischer in their 1977 paper. TE, Tuned excitatory, Tl, Tuned inhibitory. The N ear and Far cells fall into the asym m etric category, and the TE and TI cells into the tuned category. The positions of the curves on the x axis are relative to the position of the fixation p lane.
Poggio and Fischer's recording set-up had a num ber of advantages
over previous ones that enabled them to find and classify these cells:
firstly they recorded from the awake behaving monkey, trained to fixate a
particular point while its receptive fields were plotted and stim uli
presented. This carries the advantage that one always knows the position
of zero disparity and eye movement problems were eradicated. Also the
stimuli were presented in real depth relative to the depth of the fixation
spot (i.e. the stim ulus screen was moved tow ards or away from the
animal) rather than using disparate stimuli or superim posing receptive
fields, as had been used in the previous studies. An important feature of
fixation plane. Poggio and Fischer found th at m ost of their cells w ere
tuned to points w ithin ±0.5° of the fixation plane. This corresponds to a
real distance in depth of roughly ±3cm at their particular fixation distance,
a value roughly equal to Panum 's fusional area. They concluded that the
range of optim al disparities they found (For TE and TI cells) w as the
neural correlate of Panum 's fusional area (Poggio and Fischer 1977).
Since their 1977 study, Poggio et al. have found disparity tu n ed cells in both V2 and V3 (Poggio et al. 1985; Poggio et al. 1988). In their 1988 paper they enlarged their categories of disparity tuned cells to include the
tuned near (TN) and tuned far (TF) types: these neurones have response
profiles sim ilar to the tuned excitatory type, b u t their responses occur at
disparities betw een ±0.01° to 1.0° thus, they differ from TE cells in their
location, b u t not the w id th of their optim al response. These neurones
are in a m inority in VI (14/31), increase in V2 (21/30) and are com m on
in V3/V3A (23/26) (Poggio et al. 1988). The eccentricities of different types of d isp arity tu n ed cells w ere also studied by the authors b u t since the
m ajority of the cells they investigated in VI and V2 had their receptive
fields w ithin 4°, and all of their V3 cells were at eccentricities greater than
4°, th ey w ere n o t able to p ro v id e any strict co rrelatio n b etw een
eccentricity and stereotuning.
The p ro p o rtio n s of the various types of d isparity tu n ed cells in
different areas have also been the subject of investigation and it has been
show n to vary in different visual areas. The p ro p o rtio n of tu n ed cells
versus asym m etric cells changes as the level of hierarchical processing is
increased. In VI, V2 and V3 tuned cells and asymmetric cells are in equal
proportion of asymmetric cells increases to about 90% (Roy et al. 1992) and in the lateral intraparietal area (LIP) the figure is 100% (Gnadt and Mays
1991).
The functions of different types of disparity tuned cell
Since "tuned" cells (TE, TI, TN and TF cells) are often present at
the early stages of visual processing it has been suggested that they m ay be
p re d o m in a n tly involved in the stereo-m atching process to m ain tain
accurate stereo fusion w ithin Panum 's fusional area (Trotter 1995). The
w idth of tuning of TE/TI cells w ould therefore correspond to the lim it of
stereo acuity (Poggio and Fischer 1977). The asym m etric cells' function
m ay be m ore to do w ith the control of convergent and divergent eye
m ovem ents and to tell the visual system w hether an object ap p earin g
outside Panum 's fusional area is in front of or behind that plane (Poggio
and Fischer 1977). Indeed the near and far cells in LIP have been show n
to be m odulated by vergence (Gnadt and Mays 1991) and this area projects
to the su p erio r colliculus, a region assigned to oculom otor control
(Sparks 1986). Asymmetric cells, especially the near cells, m ay also have
a function in the control of h an d m ovem ents because they resp o n d to
stim uli nearer to the animal than the fixated object, and could be useful
for the direction of hand m ovem ents tow ard th at object. The changing
proportions of tuned and asym metric cells m entioned above suggests that
the fu rth er up the parietal sequence of areas one progresses, the greater
Other types of disparity
Disparity is a general term encompassing all types of difference
between the two monocular images of an object. These differences may
be further subdivided: position disparity is the shift in horizontal
position that arises from an object being displaced from the fixation plane;
orientation disparity is the perceived difference in orientation between
each m onocular image of a line which tilts in depth tow ards or away
from the observer (illustrated in figure 6).
1
Figure 6.
Illustration of the orientation difference betw een each im age on the retinae w h en a line tilted tow ards or aw ay from the observer is view ed binocularly. L and R represent the (exaggerated) im ages on the left and right retinae of a single line tilted in depth, the cube is added to aid perspective view ing. By analysing the angle 6 the inclination of the line can be calculated and depth perceived.
D isparity curvature is the difference in perceived cu rv atu re
between the two monocular images of a surface of a three dimensional
object. Shapes rotated about their vertical axis cause disparity of
horizontal width. Together these differences are analysed to produce a
position disparity allows one to know the distance of an object from the
fixation plane, b u t alone it gives no cues to the three dim ensional form
of an object. One of the constituents of three dim ensional form is tilt in
depth. Since we are capable of discrim inating w hether lines are rotated
about a horizontal axis, one m ight expect to find cells selective for tilt in
dep th in the visual cortex. In effect, a cell selective for tilt in depth could
com prise a cell whose two m onocular receptive fields are each tuned to a
slightly different orientation, hence the term "orientation disparity".
Since our eyes are separated in a horizontal direction it is only possible to
perceive tilt in d ep th w hen view ing vertically or near vertically oriented
lines (neglecting all other d ep th cues). Cells of this type w ere indeed
found in the cat's striate cortex (Blakemore 1970) and since in m onkey VI
(H anny et ah 1980).
So called "tilt in depth" cells give an optim al response w hen each
eye is p resen te d w ith a line slightly differing in o rientation. Eye
m ovem ents will cause problem s w hen looking at this type of cell because
cyclotorsion will m ake all cells appear to be selective for tilt in depth.
Blakemore (1970) elim inated this possibility by recording from m any cells
in a single anim al w hose eyes w ere held still (by suturing them to firm ly
h e ld m etal rings) and dem o n stratin g th at the difference in the cell's
preferred orientation betw een the two eyes varied significantly from cell
to cell (Blakemore 1970). Another feature of these cells was that they w ere
also selective for horizontal disp arity , and their responses could be
occluded if the optim al orientation disparity stim ulus w as not presented
at the correct horizontal disparity. The existence of cells selective for tilt
in d ep th in the m acaque was disputed by H ubei (Hubei and Wiesel 1977)
w h o m ain tain ed th a t the p ro p erties of each receptive field rem ain
found th at have opposite direction selectivities in each eye (Zeki 1982) it
is not unreasonable to suggest that VI cells m ay have slightly different
orientation preferences for each eye.
Some cells have been found to m ed iate an o th er a ttrib u te of
stereoscopic vision, th at is m otion in depth (Pettigrew 1973; Regan and
Beverley 1973; Zeki 1974a). Stim uli com prising a fixed d isp arity are
perceived at a fixed depth but if disparity is changed w hile the stim ulus is
b ein g v iew ed , m otion in d e p th is perceived. The d irectio n of 3
dim ensional m otion is dependent u pon both the direction of the images
in bo th eyes and the direction of change of disparity. By changing these
param eters, trajectories of any direction can be p roduced. Pettigrew
fo u n d cells in area 18 of the cat th at signalled changing d isp a rity
(Pettigrew 1973), and later C ynader et al (1978) found cells in area 18 of cat's visual cortex th at w ere selective for specific trajectories; p erhaps
unsurprisingly, the m ajority of cells found w ere selective for trajectories
th at w ould result in the stim uli coming close to, or actually hitting the
anim al (Cynader and Regan 1978). Zeki (1974a) found cells in V5 of the
m acaque th at had opposite direction selectivities or each eye and w ould
therefore respond m axim ally to m ovem ent tow ards, or aw ay from, the
,
---^ b '/
XV
Figure 7.
Diagram to sh ow that w hen a point at A ,
having its retinal images at a
and a', is displaced to B,
having its images at b and b',
the d isp la c e m en t is in opposite directions in the two e y es (a ssu m in g no eye movements take place). Cells w ith o p p o site d irectio n selectivities in each eye have been found in V5 (Zeki 1974a).
H ow ever, the existence of such m otion in d ep th cells in m onkey
area V5 w as d isputed in 1983 (M aunsell and Van Essen 1983a) because,
alth o u g h th ey ap p eared to respond w ell to m otion in d e p th w h en
stim ulated at trajectories far from the unit's optim al fixed disparity, their
best response w as still to m otion in the fronto-parallel plane, i.e. at a
fixed disparity. This m ay be true for some trajectory cells, b u t cannot
stand for cells w ith opposite direction selectivities in each eye as reported
by Zeki in V5 (Zeki 1974a). It has since been suggested th at m otion in
d ep th is not analysed by single units, b u t by the change in o u tp u t from
m any units, each tuned to a fixed disparity (Gum m ing 1995). W hether
the o u tp u ts from these units w ould have to converge onto a single cell
for the m otion in depth to be perceived is unknow n.
Psychophysics
Stereopsis has been studied extensively by psychophysicists because
it is an accessible attribute of vision to study in this way. The advent of
the ran d o m d o t stereogram inspired a d elu g e of stu d ies because it
p e rc ep tu a l p ro p erties of stereopsis. The form s seen in ran d o m d o t
stereo g ram s are also invisible m onocularly and therefore, m u st be
constructed once the visual signals have reached the cortex.
V ario u s a u th o ritie s have su b d iv id e d h u m a n ste reo p sis into
categories; here I shall review each of these (som etim es overlapping)
divisions form ed m ainly on the basis of psychophysical studies.
Local/global
In order to prevent the false m atching of the small black and w hite
squares th at constitute random dot stereogram s, a type of stereopsis th at
exam ines m ore th an the local features of the visual scene m u st exist.
Julesz called this type of stereopsis "global" (Julesz 1971) because it has to
take inform ation from a large area of the visual field in order to prevent
m ism atching of the identical local features. Local stereopsis is therefore
the m ech an ism req u ire d to p erceiv e d e p th in the m ore classical
stereoscopic scenes w here there is no chance of false m atching because
each p a rt of the m onocular image is unique. Thus, the division of local
and global stereopsis m ay be looked upon as an absence and presence
respectively of interactions betw een different parts of the visual field at or
beyond the initial disparity processing level. A lthough this classification
of stereopsis w as p ro d u ced entirely on the basis of psychophysics it
corresponds well to a physiological p roperty of cells, i.e. their receptive
field size. Cells w ith large receptive fields w ould be able to u n d ertak e
global stereopsis because they can take inform ation from larger areas of
the visual field, w hereas, for exam ple, striate cells, w hich have very
sm all receptive fields, w ould be better suited to the analysis of local
Fine /coarse
The division of stereopsis into fine and coarse is one of m agnitude
of disparity. A "fine" system w ould be capable of transform ing sm all
disparities into fused percepts of three dim ensional objects. D isparities
th at exceed Panum 's fusional area cause double vision. H ow ever, even
w hen an image appears double due to its large disparity, it still carries a
dep th percept. Thus, the coarse system does not require fusion of the tw o
m o nocular im ages in order to gain inform ation about relative depth.
Physiologically, this concept could co rresp o n d to the responses of
different types of disparity tuned cell. Fine stereopsis could be signalled by
"tuned" cells w hich resp o n d specifically to d isp arities at or n ear to
Panum 's fusional area giving narrow response profiles. Large disparities
w ill n o t stim ulate "tuned" cells, b u t w ill cause "asym metric" cells to
respond because these cells are stim ulated over a large range of disparities
th at exceeds the limits of the horopter, either in front of (near cells), or
behind it (far cells).
C y c lo p e a n /n o n c yclo p e a n
Julesz coined the term cyclopean; it refers to visual stim uli th at
can only be perceived binocularly, for exam ple, the contours p resen t
b e tw ee n areas of d ifferen t d isp a rity in a ran d o m d o t stereo g ram .
Essentially, cyclopean stim uli do dot exist w hen view ed m onocularly.
The "cyclopean eye " is the concept of a system in the brain w here the
d ep th inform ation is extracted by processing disparity. The cyclopean eye
is therefore the p art of the brain able to 'see' depth contours defined by
disparity in random dot stereogram s. A non-cyclopean stim uli will still
appear on the cyclopean eye, b u t unlike the cyclopean image it will also
appear in m onocular images. This concept corresponds to the physiology
stim u la tio n , b u t w ho's resp o n ses are only tu n e d w h en th ey are
stim ulated binocularly at the correct disparity.
The correspondence problem
In order that two retinal images be fused, the brain has to decide
w h ich p a rts of each m onocular im age correspond to the sam e object
v iew ed by the other eye. This is the correspondence problem . From
Julesz's ran d o m -d o t-stereo g ram studies in the 1960s, w e know th at
binocular correspondence happens early in the visual system, i.e. before
object recognition, and th at the stim ulus requirem ents necessary for
c o rresp o n d en ce (as w ell as stereoscopic d ep th ) do n o t n eed to be
so p h is tic a te d . The exact stim u lu s re q u ire m e n ts n e ce ssa ry for
correspondence have been studied psychophysically for m any years.
Colour and stereopsis
Several lines of evidence have suggested that colour is not used by
the stereom atching system to achieve correspondence. An often used
m ethod of producing stereopsis is to view tw o im ages th ro u g h red and
green anaglyphs. One image, m ade up of red and w hite dots, is view ed
th ro u g h a green filter placed over one eye, producing a black and green
pattern, and the other image, m ade up of green and w hite dots is view ed
th ro u g h a red filter producing a black and red pattern. The tw o patterns
are superim posed, avoiding the need for divergence or crossing of the
eyes (as in Julesz's random dot stereogram s). W hen this is done, the
region of the p attern s containing d isp arate dots is seen stan d in g o u t
vividly in d ep th even though each eye sees it in a different colour. Thus
it w as suggested th at colour cannot be im p o rtan t for the stereoscopic
coloured filters to explore this phenom ena; polarized filtered random dot
stereogram s containing different coloured dots (red, green, blue and
yellow) w ere prepared and the coloured dots w ere arranged so th at no
spatially corresponding dots w ere of the same colour, the colours being
shifted in one direction. The result w as a stereogram that contained tw o
cues: colour and position; the view er could p a ir the dichoptically
presented dots on the basis of their colour or their position. In this case
co lo u r b ased m atch in g w as possible, b u t the m ore com m on a n d
preferable m atch w as based on the position of the dots, p ro d u cin g an
id e n tic a l sen sa tio n of d e p th as w h en p u re c o n tra st ran d o m d o t
stereogram s w ere used.
Isolum inance studies
Isolum inance is used by psychophysicists (and physiologists) as a
tool to p re se n t stim u li w hose contours are visible d u e p u re ly to
chrom atic (and not luminance) differences. Presenting various types of
stim u li at isolum inance is a m eth o d of disco v erin g w h a t role the
chrom atic contrast sensitive system in the brain plays in the perception of
the p a rtic u la r stim ulus. A stim ulus th at is u n d etectab le or w hose
detection is adversely affected w hen presented at isolum inance w o u ld
probably be analysed by a part of the brain w hich cannot draw know ledge
from chrom atic contrast alone. Various experim ents have been carried
o u t u sin g isolum inance and disparity, som e of w hich have p ro d u ced
conflicting results.
Lu and Fender (1972) found that stereoscopic d ep th perception is
difficult or absent in fine random dot stereogram s com posed of tw o
iso lu m in an t colours rather th an black and w hite. Their stim uli w ere