THE INFLUENCE OF LIGHT AND
TEMPERATURE ON THE ESTIMATION
OF AUXINS BY A STRAIGHT
GROWTH TECHNIQUE
BY C. L. MER, S. H. CHOUDHURY,* P. DATTARAY AND U. HAFEEZ:j: Agricultural Research Council, Unit of Plant Physiology, Imperial Co/hge of Science
and 1'echno/ogy, London, S. W. 7
PREAMBLE
THE detailed investigation into the technique of assaying auxin, an outline of which is presented in this contribution, was initiated by Prof. Gregory in connexion with his researches on Vernalisation, carried out over many years in collaboration with Dr. 0. N. Purvis. They accumulated extensive data about the variations in the sequence of developmental changes shown by the Rye plant following different vernalisation and photoperiodic treatments and, to account for their observations, they supposed that a chain of reactions occurred, involv-ing transformations between a number of substances some of which they considered to be responsible for particular stages of development.
This series of reactions constitutes the well-known schema:
A
Low temp.
"
"
High temp.
Low temp. A'---"'"
E
-t
Short Long B===-~"
C--)-D"
Days Days and it embodies the fundamental idea of Sachs, about organ-forming substances, and of Klebs, about the stage of 'ripeness to flower'. Although derived from a study of the variations resulting from diverse treatments, it was inferred that this sequence of reactions occurred in normal plants.For winter rye the vernalisation reactions are represented by the sequence from A to B. A is regarded as a precursor substance which is transformed to B; slowly at ordinary temperatures, more rapidly at low temperature. Before the reaction is completed it can be wholly
"' Present address: Botany Department, Masterpara, Rajshahi, East Pakistan.
; Present address: Botany Department, Government College, Quetta, West,,
Pakistan. ·
98
C. L. MER AND OTHERSor in part reversed by high temperature (35° C) depending on the duration of low temperature treatment already undergone. Germi-nating grain devernalised in this way can be subsequently fully reverna-lised by further exposure to low temperature. As fully vernareverna-lised plants cannot be made to revert to the untreated condition by high temperature-thought to indicate that A has been completely con-verted to B,-B must be regarded as heat-stable. Hence the need for a reversible reaction between A and A', whilst an irreversible reaction must be formulated between A' and B.
Spring rye, which is insensitive to low temperature, i.e., supposedly
devoid of A, is considered to form B under gene control. The re-actions from B to D are regulated by day-length: D is considered to be the flowering hormone. Substance E has been called a "leaf hormone" to account for the leafiness of plants which have been pre-vented from shooting by inappropriate day-length regimes (conti-nuous short days).
With the passage of time and the absence of any results conflict-ing seriously with these postulates, Prof. Gregory decided that some effort could be expended in a search for a substance, or substances, whose behaviour might agree with the supposed succession of trans-formations. 'B' attracted immediate attention for spring and winter rye differ specifically in their content of this substance. But because of the prevalent view that auxin might be related to flowering, the variation in auxin content in relation to temperature was ultimately formulated as a definitive research project.
INFLUENCE OF LIGHT, ETC.; ON ESTIMATION OF AUXIN .
99
assess the relevance of Auxin to Vernalisation the root-inhibition assay was used. This occurred when T. M. Das arrived from Calcutta to study with Dr. Purvis and he brought with him some experience of this pa-rticular technique. His work revealed little or no change in the auxin content of the apex in relation to low temperature treat-ment. With the arrival of Das's successor S. H. Choudhury, Prof. Gregory transferred to me the responsibility for the auxin-assay work. We started by using Das's technique but soon discarded it. The estimates were obtained from the mean length of the longest root of 10-15 replicate plants, each of which was grown on 7 ml. of agar-medium. To ensure the presence of an adequate amount of growth substance in this large volume of medium, a considerable weight of tissue was required for each extract. Also, many roots, instead of descending directly downwards into the medium, grew out into the air above the surface of the agar slope for variable distances before they entered it to become affected by the hormone contained therein. After the removal of the seedlings from the test-tubes in which they had been growing, it was quite impossible to tell which root had been growing in the medium and which had not. These difficulties were in part obviated by using the method of growing pj.ants singly in tubes which had been devised for the investigation of the. relationship. bet-ween riboflavin and growth· (Mer; 1957). With narrow bore tubing, the volume needed for each plant was reduced to 2 ml. (i.e., 20 mi.
for the 10 replicates comprising a single estimate), and the restricted space ensured that the roots were in contact with the culture medium at all times during their growth. Choudhury at once found a ten-fold loss in sensitivity as comp~red with Das's data. Also, the rela-tionship between length and IAA concentration was not linear and furthermore, the accuracy was not as good as had been hoped. In a series of tests, measurements were made of the content of IAA in a range of solutions; the estimates varied between 60% and 95% of the known content. Because of its lack of specificity this root-inhibi-tion method has not been widely adopted; virtually all active sub-stances will inhibit root growth so that promoters and inhibitors cannot be recognized. Professor Gregory had an intimate knowledge of the Avena "Curvature" method, through his long association with Hatcher's work and also of Hancock and Barlow's straight-growth method, which he recommended us to use. The situation presented here was then encountered.
INTRODUCTION
While using the Hanco~~ and Barlow auxin-assay techriiq~.'(i9.53) Choudl),]fY-. n,ot~d, in 22 calibr!!tion~ out of 29 performed, t!J..~.t a low :conceti~~tion ,;e>f indole _acetic aci~ ...
D
o-
2 mg/1) inhibited the grpwth100 C. L. MER AND OTHERS
60
40
--0,-- ---·
20
-2 -J. 0 J
LOG I A A CONC£NrR~Ti OIV
J:iiG. l. Calibration of IAA concentration using Rye coleoptile segments, to show the significant depression occasioned by to-~ mg./1 lA A The upper and lower fiducial limits are indicated by the vertical lines through the points.
by a broken line. The growth depression due to treatment with I0-2 mg/1 IAA there shown was statistically sir:nificant, and so the
lowest concentration of IAA which would be estimated with certainty was approximately 5 x I0-1 mg/1. Inevitably, a solution known to
- contain IAA at a concentration of I0-1 mg/1 was estimated to be
devoid of growth substance.
The value of the technique for estimating auxin was thus consi-derably reduced and experiments have been carried out with the object of avoiding this difficulty and so improving the usefulness of this method of assaying plant growth hormones.
MATERIALS AND METHODS
Wheat grain (var. Heine VII or Jufy), sown thickly on a
f'
deep bed of moist vermiculite in shallow polystyrene dishes and covered to a depth of!"
with further moist vermiculite, was allowed to ger-minate in the dark in a closed growth chamber at 22° C. Whilst growing for 31-4 days in darkness the plants were continuously swept by a stream of air purified by passage through an activated-charcoal filter and subsequently moistened with water. From the coleoptiles of these seedlings 10 mm cylinders were cut in dim orange light using a conventional cutter with which the 3 mm tip segment could be readily discarded.with either 0·5 ml. of buffer solution (0·01 M KH2P04 , pH 4·5, with
2% sucrose) or the same volume of solution plus an appropriate amount of IAA to form the necessary range of concentrations for calibration purposes (usually I0-2 ; 5 x I0-2 ; I0-1 ; 5 x I0-1; 10° mg/1). Ethanol
was used to dissolve the IAA; the necessary volume was included in the control solution to ensure uniformity of treatment. For each IAA concentration 5 (or occasionally 10) replicate specimen tubes were prepared and these were placed horizontally in small cylinders (aluminium cigar cases) fixed to the wheel of a klinostat which rotated slowly during a 24-hour growth period. The increments in length of the cylinders were then determined; the total increase of 5 cylinders was regarded as a single observation.
Treatments
(i) Darkness.-10 mm coleoptile cylinders for use in assays were
prepared one at a time in darkness using a special cutter made from a short length of perspex rod (Fig. 2). Into one end of the rod a funnel-shaped hole was drilled; the 'stem' (2 mmidiameter) was
A
I
- - - . - _
--;-~:_.-::----C. L.' MEl~ AND· OTIDlRS ·
traversed by two slots respectively 31nri1 and Hmm .. fi'om the dosed (internal) end. The cone-shaped mouth facilitated :the insertion Of
a
coleoptile into the stem in darkness, and when it could be pushed in no further, a hinged carrier with two blades attached to it was brought down to engage with the two slots. As shown in Fig. 2 B, the slots were made by removing about 75% of the tube wall so that two annuli remained connected together as a unit by bridges of pers-pex (shaded area), which also served as a stop for the razor-blades. Before using the apparatus in darkness the perspex rod was rotated so that the blades were halted when the coleoptile was partially cut through (Fig. 2 B) and then fixed in this position; after the blade-carrier had been lifted the cut, but whole, coleoptile was withdrawn. The 3 mm tip segment was then broken off and discarded; the 10 mm cylinder was carefully removed and placed in a specimen tube waiting to receive it, already provided with 0 · 5 ml of either buffer or IAA so1ution. Again, 5 replicate tubes were prepared for each IAA con-centration and these were inserted into predetermined places on theklinostat. ·
(ii) Illumination.-1 hr., 1 hr. or
3
hr. exposure to dim orange light of intensity about 5 f.c. The seedlings were illuminated during the preparation of the cylinders either before or after heating and chilling treatments.(iii) Heating.-About 100 coleoptiles detached from the endo-sperm were wrapped in moist filter-paper, placed vertically in closed jars containing a little water and heated for 1 hr. at 40° C in darkness. (iv) Chilling.-Coleoptiles prepared as in (iii) above were sub-jected to low temperature 0
-4°
c
for 2 days in darkness.Extraction and Chromatography of Auxins
Growth substances were extracted by immersing replicate samples of 5 coleoptiles (with leaves) for 24 hours in darkness at
oo
C in 5 ml of ether, freshly distilled over ferrous sulphate. In dim orange light the ether was poured off and the plant material briefly rinsed three times, using 5 ml of ether on each occasion; washings and extract were combined. The plant material was dried in vacuo at 50° C and weighed.The ether extract was thoroughly shaken with 9 ml of 1% sodium bicarbonate solution for 2-3 minutes; the aqueous phase was removed in a separating funnel and the procedure was repeated (Larsen, 1955).
After acidifying the 18 ml of alkaline solution to pH 2 · 8-3 · 0 with 0 · 5 N HCl, it was evaporated almost to dryness in darkness with a stream of warm air. The active substances were then exiracted in a"measured volume of acetonitrile (Nitsch, 1955), an aliquot pf which ·"w.a& applied to a 1 "-wide strip of 'Yhatmari filter~paper and chromlj!t~;-,:
graphed using water as an ascendmg eluant (N1tsch, 1955). AfterJ~
each of which was put into a specimen tube containing 0 · 5 ml of buffer solution and 5 coleoptile cylinders. Simultaneously with each group of assays performed at one and the same time, a set of standard IAA solutions was included for calibration purposes.
Statement of Results
The data obtained are expressed as p. gm auxin per 100 mg dry-weight of material. As an illustration of the method of calculation one result is set out in detail.
The equation for the line of best fit to the calibration points shown in Fig. 3
was-Y
= 45·92
+
10·22 (x+
0·92)where 'y' is the increment of growth in mm, and 'x' is the logarithm of the auxin concentration. The control segments elongated by · 27 · 2
±
0 · 77 mm, so that increments in excess of the upper fiducial limit of 29 · 3 mm indicated growth-promoting activity in the solution.~
60~
~
so
0~ ~
~
40 0
~
~ ,
~ /
/
0 30
b//
~
-2 -.1 0
LOG 1 A A CONaNTRA T!Oh
FIG. 3. Calibration of lAA concentrations using cylinders cut from detached coleoptiles of Wheat var. Jufy, which had been illuminated and subjected to high and low temperatures. The upper and lower fiducial limits (for control and J0-2 mg/1 IAA respectively) are indicated by the vertical lines.
The coleoptile cylinders included with the successive pieces of filter-paper elongated by
28·0: 28·0: 28·0: 29·0: 28·0: 30·0: 37·0: 32·5: 28·5 and 28 ·0 mm. respectively. Evidently active substance was present only in the three tubes, for which the growth increments are underlined.
Substituting in the equation gives
for y = 30·0: x = -2·477 = 3·523 = ·00333 p. gm/ml
, y = 37·0: X = - 1·792 = ~·208 = ·01614 , , y
= 32·5:
X = - 2·232= 3·768
=
·00586 ,'104 C. L. MER AND OTHERS
For this chromatogram 1/lOth of the original volume of acetonitrile was applied to a strip of a filter-paper; hence the growth-promoting substance would have been present at a concentration of 0 · 2533 ,.,_ gm/ ml. As this was presented to the coleoptile segments in 0 · 5 ml of buffer, tl-.\! actual weight of auxin present was 0 · 127 ,.,_ gm. The dry-weight of the coleoptiles extracted was 10 · 6 mgm; the auxin content was therefore 1·19 ,.,_ gm /100 mg dry-weight.
Alternatively, the regression can be computed in terms of weight of IAA, as has been done for a particular purpose in the discussion
(p. 111).
ExPERIMENTAL RESULTS
(1) The Effect of l!lumination.-The effect of illuminating test coleoptiles of Heine VII on the response to IAA is displayed in Fig. 4. Elongation of the control segments was not affected by exposure to light, but a growth depression resulted when a small ( · 005 fi-g) but not a larger amount ( · 05 ,.,g) of IAA was present in the medium. Evidently, the inhibition represents an interaction between the illu-minated tissues and the small amount of IAA infiltrating into them.
70
~
~.... ~0
~ ~
~
~
50
~
iec,-,--~
0
~
40
-J -2 -1
LOG lA A CONCENTI?A TION
FIG. ~- The effect of illumination on the response of coleoptile cylinders to IAA solid points-in darkness; open points-illuminattd.
!0
~
A,
~
...
~
•
~ so (5
~
0· •••
~ lt
~
•
A 6
JO
-t -1
_,
-2
LOa IAA COIYctNTRATIOit
FIG. 5. The interaction between temperature and light on th; calibration of IAA using Wheat var. H.:ine VII. A-no temperature treatment; (B) High and low emperature treatment.
e., in darkness; 0, I hour dim orange ligh,; CD, 1 hour dim orange light.
to small amounts of IAA became somewhat more pronounced. Also, the growth-promoting effect of the higher concentration of IAA diminished with the longer exposure to light. When heated and chilled coleoptiles were used (Fig. 5 B) a concentration of IQ-2 mg/1
promoted growth in darkness. Although a I hr exposure to light markedly reduced the promotive effect of the IAA solution containing I0-2 mg/1 a depression was not recorded because the elongation of
the controls was correspondingly reduced. Also these particular cylinders of tissue elongated in a solution of l0-1 mg/1 IAA almost
as much as did those grown in darkness; that is their response to the externally provided IAA was not impaired by the temperature treat-ments, and a steeper calibration curve resulted. With a 3-hr exposure to light the inhibition due to I0-2 mg/1 IAA reappeared; the increment
of the cylinders without IAA, although much reduced by illumination, exceeded that of the cylinders provided with · 005 mg IAA.
These results agree with those previously presented in showing that a growth inhibition occurred when illuminated tissues were provided with a small amount of IAA. From the viewpoint of a cali-bration and the measurement of a concentration of 10-2 mg/1 they
106 . C, :1:·: MER AND OTHERS
exposure to light. They show that IAA at a concentration of lQ-2
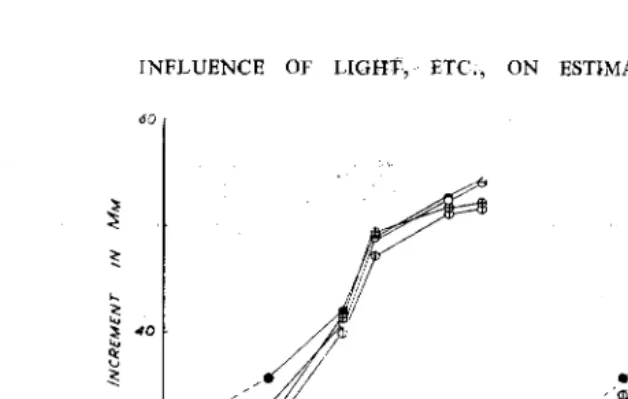
mg/1 can be successfully estimated, for a slight growth depression occurred only twice (Nos. 11 and 15). Some individual calibrations showed a departure from linearity with the highest concentration used (1 mg/1). The use of these regions of the response can be avoided, however, by using extracts with a low auxin content, so utilising that range of concentrations providing the steepest response curve as shown in Fig. 6, which depicts the mean curve derived from these data.
80 p--0
~ 70
~
...
~ 60
~
~
~
50
~ 31:
0
~ /
/
40 /
/ 0/
-2 - j 0
LOG I A A CONCENTRATION.
FIG. 6. Mean Calibration curve of lAA concentrations derived from the data of Table I.
During the course of the work the stock of Heine. VII became exhausted and it could not be replenished. A fresh start was therefore made with an alternative variety, Jufy. The response of coleoptile cylinders of this variety .to the range of IAA concentrations is sh?wn in Fig. 7 (A and B agam refer to untreated, and heated and chtlled respectively). These coleoptiles. were much less sensitive t.o light than was Heine VII, and the depressive effect of low concentratiOns of TAA was not very marked. Also, their sensitivity to IAA was much less; the largest increment recorded was 54 mm as compared with 89 for Heine VII (Table I). The effect of heating and chilling is clearly apparent; growth of the controls was reduced while the response to externally applied IAA was not impaired. A concentration of IQ-2
mg/1 was easily detectable as all th~ points at this concentration were significantly in excess of the elongatiOn of the controls. .. A 3-hr illu-mination was undesirable in view of the loss of sensitivity to the highest concentrations. The calibration shown. in Fig. 3 was also obtained with Jufy wheat.
(3) The Auxin Content of Wheat Coleoptiles.-Coleoptiles of Jufy
60
Jm
~···
· / = •
.---e- ft\
,_';
A 8
?0
-1 0 -? -J 0
l.OG IAA CONaNTRATION
FIG. 7. The interaction between temperature and light on the calibrauon of IAA using Wheat var. Jufy. (A) No temperature treatment; (B) High and low temperature treatment.
e, darkness; 0, 1 hour dim orange light; (!), 1 hour dim orange light; EB 3 hours dim orange light.
obtained by combining Heating (I hr. at 40° C), Chilling (2 days at 2-4° C) and Light (15-30 mins) in a 23 factorially designed
experi-ment. Auxins were extracted separately from 5 replicate samples each of 5 coleoptiles (with leaves) and assayed. In experiment 1, the five samples of each treatment were taken at the same time, so that differences between occasions were included with treatment effects. In experiment 2, groups of different treatments were dealt with on 7 occasions to make up the necessary total of 40 observations. For each assay a separate calibration was performed at the same time and the regression equation obtained was used for the computation of the auxin contents which are recorded in Table IT; analyses of variance are also quoted.
Except in two instances, H and HL, the values agree as between experiments, in spite of the inclusion of occasion effects with treat-ment in experitreat-ment 1. The error mean square in the two experiments was also virtually identical. The data show that only H and C signifi-cantly reduced the auxin content in the coleoptiles, and in experiment I ' the interaction H XC reached significance, and was promotive. The effect of light on auxin content was negligible, and in view of the apparent interaction between chilling, heating and light on elongation, the absence of a significant interaction in auxin content, is noteworthy.
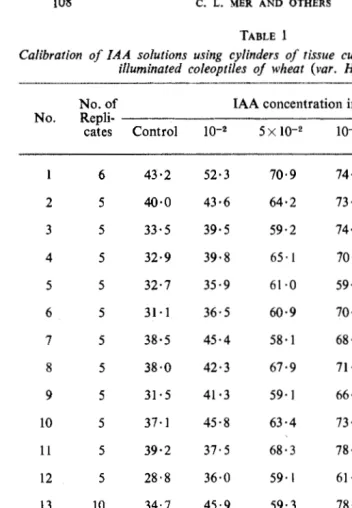
108 C. L. MER AND OTHERS TABLE 1
Calibration of IAA solutions using cylinders of tissue cut from heated, chi/led and illuminated coleoptiles of wheat (var. Heine VII)
No. of IAA concentration in mg/1
No.
Repli-cates Control IQ-2 5 X IQ-2 IQ-1 5x I0-1 lQO
6 43·2 52·3 70·9 74·8 82·4 89·2
2 5 40·0 43·6 64·2 73·3 86·5 84·1
3 5 33·5 39·5 59·2 74·8 82·5 81·2
4 5 32·9 39·8 65·1 70·5 79·2 77·4
5 5 32·7 35·9 61·0 59·5 69·1 72·8
6 5 31·1 36·5 60·9 70·0 68·9 74·4
7 5 38·5 45·4 58·1 68·1 78·3 80·0
8 5 38·0 42·3 67·9 71·7 80·3 82·0
9 5 31· 5 41·3 59·1 66·3 76·8 74·8
10 5 37·1 45·8 63·4 73·8 77·3 87·0
11 5 39·2 37·5 68·3 78·4 77·8 75·2
12 5 28·8 36·0 59·1 61·7 71·2 74·2
13 10 34·7 45·9 59·3 78·3 83·0 86·8
14 10 43·6 46·8 63·7 77·1 83·9 84·9
15 10 44·6 44·0 58·6 67·6 78·8 72·1
16 10 45·7 48·2 64·3 77·5 85 ·1 88·4
17 10 40·9 45·6 61·7 65·9 84·1 82·1
18 10 38·8 44·2 61·2 73·4 84·9 86·9
19 10 42·9 47·4 66·4 77·6 81.5 84·0
20 10 39·6 44·3 64·9 66·8 83·3 86·2
·--~--···----~-ot
109
TABLE II
Auxin contents of wheat coleoptiles (and leaves)
Each entry is the mean value p.gy'JOOmg dry weight of 5 replicate determi-nations each of 5 coleoptiles (with leaves) of wheat, var. Jufy. The treatments were Heating (H), Chilling (C) and Light (L) combined in a 28 factorial
arrange-ment, (I)= Controls.
~---~---·
Expt. l Expt. 2
-(l) 0·73 0·61
H 0·23 0·19
L
0·90 0·48c
0·33 0·35HL
0·33 0·19HC
0·19 0·14LC
0·31 0·36HLC
0·11 0·13
-Analyses of variance
Mean squares
Variant D.F.
Expt. 1 Expt. 2 - - - ·
H 1·241(-) ·836 (-)
L
·019 ·012c
·974 (-) ·143(-)HxL
·009 ·008HxC
·342 ( +) ·045LXC
·081 (-) ·010HxLxC
·002 ·015Error 32 ·016 ·015
110
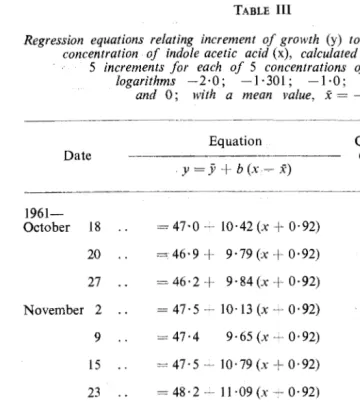
-· '•. ·C. t: MER AND OTHERSTABLE Ill
Regression equations relating increment of growth (y) to the logarithm of the concentration of indole acetic acid (x), calculated from the mean of
5 increments for each of 5 concentrations of fAA (having logarithms -2·0; -1·301; -1·0; -0·301
and 0; with a mean value,
x
= -0·92)---···----~~---~---~
Equation Correlation Standard
Date coefficient error of' b'
. y
=
ji+
b (x - x)11
DISCUSSION
In connexion with the estimation of auxins the data .presented demonstrate that an unequivocal calibration can be conducted with IAA solutions from I0-2 to 10° mg/1 as standards, using 0 · 5 ml of
solution for each of five concentrations. The growth increments- of the coleoptile cylinders in the lowest concentration used represents a definitive response to · 005 f-Lg IAA. This has been achieved by pretreating the coleoptiles used in the assay with high and low tem-peratures and exposing them to dim orange light during the prepara-tion of the tissue cylinders. In this way the growth increment of the control cylinders is so reduced that the effect on elongation of · 005 f-Lg
IAA can be detected. The reason for the reduced growth would appear to be some destruction of natural auxin by the temperature treatments, as is shown by the data in Table II. Although light seems to have exerted a marked effect on the growth of cylinders of wheat, var. Heine VII (see Fig. 5) its influence on the elongation of the second
variety, Jufy, was not nearly so pronounced (with the exception of 3 hr. exposure in conjunction with the higher concentrations of IAA, Fig. 7 B). In agreement with these observations the effect of light on the auxin content of the coleoptiles of variety Jufy was also negli-gible; the analyses of variance (Table U) show that only one inter-action (experiment I; L X C) reached significance. It is possible that the two varieties in fact differ in their response to light, but the data necessary to support such a suggestion are not at present avail-able and must await the acquisition of a further supply of wheat, var. Heine VII. Since the exposure to light employed had so little effect on the content of native auxin and did, in some way, decrease the growth of the control cylinders, it is obviously advantageous from a purely practical point of view to illuminate the plants during pre-paration rathe'r than to work in darkness.
The presentation of the data as f-Lg auxin/100 mg dry weight of tissue involves the assumption that the 5 coleoptile cylinders absorb all the auxin or IAA originally presented in the 0 · 5 ml of solution. Consequently the growth increment can be regarded as a measure of the quantity of active substance, as well as of its. concentration. To
justify this assumption we start from the original ohservation.:that a volume of solution containing · 005 f-Lg IAA when applied to coleop-tile cylinders for 24 hr. inhibited growth whereas one containing more IAA promoted elongation (Figs. 1 and 4). Choudhury (1957) then found that a corresponding growth depression could be induced with a strong solution in IAA by dipping coleoptile cylinders in it for short periods of time (10 sec to 60 min), rinsing.them and transferring them to sucrose-buffer for 24 hr. The_ same IAA solution if, applied to cylinders in the usual way for 24 hr . .invariably gave rise to a growth promotion. It must be concluded that during the brief exposure to a growth-promoting concentration of IAA only
·a:
limited amount of active substance was absorbed, and this causeda
growth depression _itist,-aii"did ·005/-Lg when applied for 24hr .. ,,Vetylikely, this smalf112
C. L. Ml3R AND OTHERSwould then remain in sucrose-buffer devoid of IAA so, effectively, the circumstances obtaining in the 'short-period' immersion experi-ments were duplicated. In further experiexperi-ments, by applying IAA for longer periods-up to 4 hr.-Choudhury found that the growth depres-sion which occurred as a result of a brief immerdepres-sion was overcome and the growth-promoting effect obtained when the cylinders remained in the solution for 24 hr. was reproduced. Also, a direct test has been performed to find whether any growth-promoting activity remained in the solutions used in a calibration at the end of a 24-hr. growing period. After removal of the first set of cylinders, 5 newly prepared ones were introduced into the tubes. To make up, in part, for the depletion of sucrose which may have occurred, 0 · 05 c.c. of sucrose-buffer was added to each tube, but it is unlikely that the original situa-tion respecting pH and sugar content was precisely realised.
The measurements obtained, mean values in mm of 5 replicates, were,
Original IAA content Control · 005 1-Lg
Growth in 1st 24 hr. 29·4 36·7 Growth in 2nd 24 hr. 27·6 26·4
50·4 27·1
54·8 27·3
Evidently the solutions were exhausted of IAA by the growth of the original cylinders; the assumption made can therefore be experi-mentally supported.
No simple explanation can be advanced to account for the inhibi-tion of growth which made this investigainhibi-tion necessary. In some way the absorption of a small amount of IAA by coleoptile cylinders, cut from seedlings which have been exposed to orange light, reduces their capacity for elongation as compared with similar cylinders pro-vided only with sucrose-buffer. This inhibition can be overcome by additional IAA and because cylinders cut from illuminated and un-illuminated plants then show either similar growth increments (Fig. 4; concentration - 1) or slightly smaller increments (Figs. 5 and 7) it seems unlikely that the sensitivity of the tissues to IAA has been changed by exposure to light. Brief exposure to dim orange light does not indeed affect the extensibility of the cylinders (control obser-vations in Figs. 4 and 7 A); the tissues must have lost neither their content of native auxin nor their sensitivity to it. Hence, as light alone does not give rise to an inhibition, its appearance here must be due to the small additional quantity of IAA.
INFLUENCE OF LIGHT. ETC., ON ESTIMATION OF AUXIN
11:3
the regeneration of a capacity to synthesise auxin (Bonner, I 933). When a coleoptile is decapitated regeneration of auxin production takes place after about 90 minutes. Thimann and Bonner (1933) have shown, however, that the onset of regeneration can be delayed when the cells, exposed at the top of a coleoptile stump after cutting off the tip, are covered by a gelatine block containing a supply of growth hormone. Upon removal of the block of agar, regenerative processes are set in train and auxin production begins again after about 90 minutes.
It is thus possible that the small amount of IAA which inhibits growth does so by delaying the onset of regeneration, and is consumed in the process; consequently it is not available for growth. But in dark-ness a growth inhibition was not observed (Figs. 4 and 7 A); by implication regeneration did not occur and the IAA provided was fully utilised for growth. It seems unlikely that regeneration would only occur in illuminated wheat coleoptiles for it is known that de-seeded oat seedlings can resume auxin production in darkness (Mer, 1951).
Hafeez (1960) attempted to explain the inhibition by supposing that a supraoptimal content of auxin was present in the cylinders at the time they were removed from the coleoptiles. In this event addi-tional IAA could give rise to an inhibition; but it then becomes impossible to account for the growth promotion which occurred when even further IAA was provided. A more intense inhibition should have been observed.
No satisfactory and consistent explanation of this inhibitory effect is therefore available.
The relationship between auxin content and extension is also somewhat complex. For convenience, growth of cylinders in darkness will be discussed first. In the absence of added IAA treatment with high and low temperatures reduced elongation (compare 7 A and 7 B, black circles); the measurements were 31· I mm and 27 ·4 mm respectively. Assuming that these increments resulted from the presence of a discrete amount of growth substance, a value can be assigned to each. For this purpose the calibration data shown in Fig. 3 have been recalculated using the logarithms of the weights of IAA instead of concentrations. Calculation of weight of IAA from the concen-tration is thus avoided, so it is unnecessary to know the volume of the cylinders.
The observations then read:
Increment 34·5 40·1 48·4 52·2 54·4 mm
Log. wt. IAA 0 · 6990 I · 3979 1 · 6990 2 · 3979 2 · 6990 (X I0-9 gm)
The equation becomes y ,~ 45 · 92 : 10 · 22 (x- l· 78) and from it we calculate that the two increments, 31 · I mm and 27 · 4 mm, could occur in the presence of 2 · 3 7 g and I · 06 g ( >~ 1 o-u) of hormone. These are approximately in the ratio of 2 : 1. The temperature treatments. however, reduced auxin content. as shown in Table II (0 · 73 and 0 · 19;
l14 C. L. MER AND OTHERS
0· 61 and 0 · 14 fLg for controls and C -t- H in experiments 1 and 2 respectively) in the ratios of 3 · 8 : 1 and 4 · 3 : 1. The cylinders subjected to high and low temperatures thus appear to grow more than would be expected on the basis of these measurements of auxin content. Had the observed ratio of about 4 : 1 obtained then the increments would have been 31 · 1 mm and 25 · 0 mm. Such a discrepancy could occur if regeneration took place differentially and the auxin content of the temperature-treated cylinders was raised to the level of that present in those not so treated. Observations of auxin content in treated and untreated coleoptiles have been made at intervals follow-ing the termination of temperature treatment; the data, in fLg
I
100 mg dry-weight, are as follows :Hours
0 3 6 24
-Controls 1·02 2·18 1·41 1·20
C and H 0·21 0·32 0·31 0·12
---·---~ ---- · - - - · . -
-··---~---The rise and fall in auxin content with time occurred in both sets of coleoptiles but after heating and chilling treatment auxin content remained low. Some other factor at present unknown must therefore be sought to account for the discrepancy. Again, the rising growth increments of the cylinders provided with increasing amounts of IAA in darkness (Fig. 7-A and B; black circles) were also similar for treated and untreated cylinders although, because of the destruction of native auxin, the total amount of hormone available to each set must have been very different. Similarly the measurements for wheat, var. Heine VII (Fig. 5) show first that the control cylinders with dif-ferent contents of natural hormone (A and B; black circles) elongated to the same extent and secondly, that when · 005 (Lg IAA was provided additionally the treated cylinders (B; concentration,-2) grew more than did untreated cylinders (A; concentration,-2) even though in sum. less hormone was available to them. A brief examination of the data recorded for illuminated cylinders (Table II and Figs. 5 and 7) will suffice to show that similar considerations apply to them also. The conclusion must therefore be reached that it is not possible to account precisely for the elongation of these isolated cylinders under the variety of treatments employed, solely in terms of the amount of growth substance available.
AcKNOWLEDGEMENT
We wish to express to Professor H. K. Porter, F.R.S., our thanks for her unfailing interest in this work throughout its tortuous progress.
SuMMARY
Method.-Wheat seedlings (varieties Heine VII or Jufy) are raised at 22° C. on moist vermiculite in a growth chamber, through which is continuously passed a stream of moist air, exhausted of London smog by means of an activated-carbon filter.
The coleoptiles. when 3!-4 em long, are detached from endo-sperm and roots while illuminated with dim orange light (about 5 ft. C); they are wrapped in bundles of about 100 in wet filter-paper, placed vertically in small glass jars containing a little water and, n darkness, are chilled for 2 days (2-4° C) and heated for 1 hr. (40° C). In dim orange light 10 mm cylinders are cut from the coleoptiles, discarding the 3 mm tip segment, and 5 such cylinders are placed in each of a series of tubes, as follows :
(1) Contro!s.-5 tubes, 0 · 5 ml sucrose-buffer solution in each. (2) Calibration.-5 tubes for each concentration of IAA in sucrose-buffer medium; concentrations I0-2 ; 5 x lQ-2 ; I o-1; 5 x 10-1; JOO
mg/litre ; 0 · 5 ml of each.
(3) Assay.--1" square pieces of filter-paper, after chromatographic separation with water as eluant, are put into a series of tubes with 0 · 5 ml of sucrose-buffer solution.
Groups of seedlings are dealt with sequentially so that the total exposure to light experienced during preparation does not exceed 30 minutes.
A regression equation relating the two variables is calculated using the mean growth-increments observed and either the concen-tration, or weight of IAA, as required. The upper fiducial limit (5% level) for the growth-increments of the control cylinders is cal-culated; assay increments in excess of this are assigned a concentra-tion, nor weight, of IAA by substituting in the equation. The quantity of auxin so evaluated is expressed as 0giJOO mg dry-weight of material extracted.
Results.-Elongation of coleoptile cylinders in a sucrose-buffer medium is inhibited by a small amount ( · 005 0g) of indole acetic acid
(IAA) when the cylinders. cut from either Rye or Wheat seedlings, have been exposed to dim orange ligbt, but not when preparation is carried through in darkness.
116 C. L. MER AND OTHERS
temperatures prior to cutting cylinders from them. The treatment reduces the extensibility of the cylinders not receiving IAA but does not diminish their responsiveness to added IAA. Exposure to orang~
light tends to reduce further the elongation of the cylinders without IAA, so that ·005 fhg and less, of IAA can be estimated with certainty. The influence of the temperature treatments and of light on the content of natural auxin in the coleoptiles used for assaying growth-promoting substances has been investigated; light had little or no effect, but both high and low temperatures reduced significantly the content of native auxin. The equations for the series of calibrations carried out in connexion with these determinations are quoted to display the uniformity achieved on different occasions.
The bearing of the data presented on the relationship between auxin content and elongation is discussed.
REFERENCES
BENTLEY, J. A. (1950). An examination of a met ted of arx'n ~sozy nir p tl:.e growth of isolated sections of Avena colcoptilcs in test soluticr.s. J. E:xp. Bot., 1, 201.
BoNNER, J. (1933). The action of the pl8nt growth hormone. J. Gen. Pl:ysiol.,
17, 63.
CHOUDHURY, S. H. (1957). Ph.D. Thesis, University of London. HAFEEZ, U. (1960). Ph.D. Thesis, UJiversity of London.
HANCOCK, C. R. AND BARLOW, H. W. B. (1953). The assay of growth substances by a mJdifiJd straig1:t growth mJthod. Ann. Rep. East Mailing Res. Stn. for 1952, 88.
LARSEN, P. (1955). Growth nbstances in higher plants. In Modern Methods of Plant Analysis. Editors: K. Paech atJd M. V. Tracey.
MER, C. L. (1951). A criticLl study of the auxin theory of growth regulation in the m~socotyl of Avena sativa. Ann. Bot. NS., 15, 179.
- - - . (1957). A re-examination of tLc supposed effect of riboflavin on gro\1 th. Plant Phys., 32, 17 5.
Nns.cH,. J: P. (1955). Methods for the investigc.tion cf ratur;;l ilUXirs and grc\\th mh1b1tors. In The Chemistry and Mode of Action of Plant Gnwth Substances. Buttt;rworth. Editors: R. L. W~b and F. Wightman.
THIMANN, K. V. AND BONNER, J. (1933). H.e mccr.<mism of the action of the growth substance in piants. Prcc. Roy. Soc., 113 B, 126.