FORMATION AND UTILIZATION OF
ASCORBIC ACID IN THE SHOOT
APEX OF WHEAT AS FACTORS OF
GROWTH AND DEVELOPMENT
BY J. J. CHINOY
Botany Department, University School of Sciences, Gujarat Universily, A!.medab(!d
INTRODUCTION
EVIDENCE has accumulated in recent years to qualify ascorbic acid (AA) as an important growth regulator in plants. The Milan School of workers has emphasized the growth inhibitory effect of the exogenous application of AA to shoot organs of oats, peas, beans, lupines and castor beans. The increase of AA in a tissue inhibited growth essen-tially through a concomitant larger increase of DHA (Tonzig and Marre, 1961).
On the other hand the work undertaken by the associates of the present writer and himself has shown that ascorbic acid, whether exogenously applied or endogenously produced, accelerates growth and development (for references see Chinoy et al., 1961). It however, appears that the AA concentrations with which the Milan School has worked are much higher than those used in the writer's laboratory. It is now known that at different concentrations AA .produces either retardation or acceleration in growth of Avena coleoptile sections (Grover, Chinoy and Nanda, 1958).
The object of the present paper is to analyze some of the experimental evidence on the role of AA in growth and development which has been accumulating in the writer's laboratory since last more than ten years especially that pertaining to the concentration and utilization of AA and auxin level in the shoot apices of wheat in relation to the laying down of organizer centres, cell elongatiOJi and flowering.
EXPERIMENTAL PROCEDURE
ASCORBIC ACID IN GROWTH AND DEVELOPMENT
173
Varieties
The three varieties were:
1. Triticum aestivum var. New Pusa 710 was obtained through the kind courtesy of the Director, Indian Agricultural Research Insti-tute, New Delhi;
2. Triticum aestivum, var. Rival, late-spring wheat; and, 3. Triticum aestivum, var. Hansa, winter wheat, both produced at Svalof Experiment Research Station, Sweden, were obtained through the kind courtesy of Prof. Arne Muntzing, Director, Institute of Genetics, University of Lund, Sweden.
Vernalization of germinating seeds, transplanting and the sub-sequent photoperiodic treatment as well as manuring and watering were carried out as already reported elsewhere (Chinoy and Nanda, 1951; Chinoy, 1956; Chinoy eta!., 1959). In this experiment only two photoperiodic treatments, viz., (I) Normal-day (ND) of 11 hours, and (2) Long-day (LD) of 24 hours illumination were given.
Growth Data.-Gregory's method of collecting and analyzing perio-dic data for growth and flowering was used in these experiments (Gregory and his collaborators, 1917, 1921, 1926, 1928, 1928 a, 1928 b, 1932). Weekly determinations of height, tiller and leaf numbers were carried out. Morpho-physiological change in the shoot apex as well as initiation and weekly growth measurements of different organs of the spike, such as glumes, lemma, stamen, carpel and ovule were recorded.
Initiation of the organs of the spike as well as growth measurements of the same were recorded in iwo ways. Weekly samples of shoot apices were collected and fixed in FAA from the first or secon i week after transplanta·ion upto almost the full maturity of the grain. Firstly these were dissec ed and observed directly under a compound micro-scope and later under a stereoscopic micromicro-scope and growth measure-ments recorded by means of a stage micrometer. Secondly some of the fixed material was passed through the grades stained with standard safranin and fast green schedule and permanent preparations made out of them. This second method was adopted as long as the shoot apex and later the spike remained within the range of the compound or the stereoscopic microscope. In most of the cases organ initiation and growth data were obtained for all the four florets of the two central spikelets. However, in a number of representative samples for each treatment combination these observations were recorded for all the florets of all the spikelets.
Ascorbic Acid, Its Utilization and Auxin Content
174 J. J. CHINOY
the vegetative state from the second week onwards. After the develop-ment of the spike the whole differentiating primordial spike was taken as the 'shoot apex' for analysis. After the differentiation of the spikelets only the two central spikelets were taken for analysis. Similarly with the gradual differentiation of stamens and pistils the 'sampling was restricted to those parts only and finally sampling was restricted to the developing kernel from the two central spikelets of the spike on the main shoot of the plant. In a number of cases the analysis was carried out on embryo dissected from the seed. The nature of sample taken for analysis is indicated on graphs (Figs. 1-6) by an appropriate symbol. Thus: (V) signifies the last sample of vegetative apex, (R) the reproductive apex in the initial stages of differentiation (sampled in its entirety), (S) spikelets, (A) Androecium, (G) Gynoecium, (F) anthesis, (K) kernel and (E) embryo.
Methods of extraction and determination of ascorbic acid and auxin have been given elsewhere (Chinoy, Grover and Sirohi, 1957; Chinoy, Singh and Sirohi, 1958). As regards the determination of the AA-oxidation a sample was macerated and extracted in a special micromacerator (Chinoy, 1945) as shown earlier using double glass-distilled water at 1-2° C., making it up to volume, adding a known quantity of standard 2-6-dichlorophenol-indophenol to an aliquot and titrating against standard ascorbic acid solution stabilized with HP03 • Ascorbic acid concentration was calculated as mg. of AA per g. fresh weight of plant material. To the remaining portion of the extract an equal volume of a freshly prepared standard AA solu-tion (5 mg. per 50 mi. prepared without the addisolu-tion of HP03 and kept at 1-2° C.) was added and duplicate aliquots titrated immediately against standard stabilized AA solution after the addition of sufficient quantity of the dye solution (which should be slightly in excess after reacting with all the AA of the extract as well as with the amount added). The remaining portion of the extract containing the added AA was allowed to stand at 30-35° C. and at different intervals of time (say after 1, 2, 3 hours) aliquots in duplicate were removed, sufficient quantity of the dye solution was added to react with the AA left over in the extract and excess of the dye titrated against standard stabilized AA solution. The AA utilized during a given period was calculated as per cent. of the original quantity of AA found at the beginning of 1 he experiment.
Anthes is
Dates of first anthesis in the central spikelets were also noted for 20 plants for each of the treatment combinations and growth periods were worked out by determining the number of days it took from the day of transplantation to the day of first anthesis in each case. Derived Data
weekly production (numbers) of organizer centres were calculated throughout the growth period.
2. Development.-Was determined by calculating any given period of growth as per cent of the period elapsing between transplantation and anthesis.
3. Cell elongation.-Was determined by calculating the height as well as the spike length on any given sampling day as per cent. of
the maximum.
4. Period of vegetative differentiation (v) was determined m days from transplantation to the stage of double ridges.
5. Period of reproductive differentiation (r) was determined in days from the stage of double ridges to the stage of first anthesis .
. EXPERIMENTAL FINDINGS
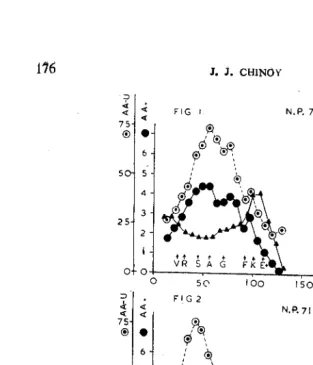
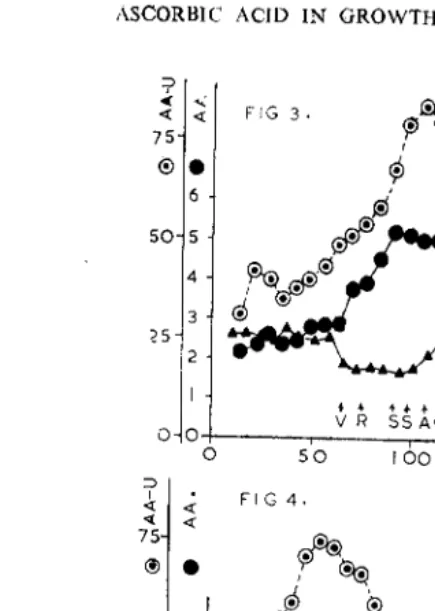
Ascorbic acid (AA) concentration in mg./g. fresh weight of the shoot apex, amount of AA oxidised in one hour in an extract of a growing apex as per cent of the total quantity and auxin content of the growing apex measured as relative growth rate of elongation by the Avena coleoptile straight-growth method are presented graphically in Figs. 1 to 6 for normal unvernalized (NDC) and long day-vernalized (LDV) plants of three varieties of wheat N.P. 710 (Figs. 1, 2), Rival (Figs. 3, 4) and Hansa (Figs. 5, 6). The change in the growing apex from the vegetative to the reproductive state as well as the differentiation stages of the spike are shown on each diagram by suitable symbols. Mean values of AA content and its utilization and mean auxin content of the shoot apex during the periods of vegetative (v) and reproductive (r) differentiation are presented in Table I.
Main results are as follows:
Concentration of Ascorbic Acid in the Shoot Ape.\·
(1) The concentration of AA in the shoot apex o · all the three varieties under both the treatments is low in the initial stages of growth. The duration for which the AA content remains relatively low in different varieties increases with the length of the vegetative period of a variety. Thus in NDC plants of N.P. 710 there is a significant rise after 30-37 days, in NDC plants of Rival such a rise comes after 70-75 days and in NDC plants of Hansa a distinct enhancement in concentration of AA occurs after 120-130 days.
176 J. J. CHlNOY
'?
<
<
FIG I N.P. 71 O-NDC< <
75
c,!i <II
®
•
<i
~~ z6 $ • ',
x
' ::J
5 5 <(
4 •4
3 i3
25
2 -2
I I I
'
+VR 5 A G
0 0
0 5(' IOO ISO
::J F' I.G 2
<!: < N.P. 710- L DV
< <
75
®
•
<IIz
6
x
::J
5 5 <
4 •4
3 ·3
25
2 ·2
J~
t + •IVR
0 so 100 ISO
GROWTH PERIOD- DAYS
FIGs. 1-2. Figs. 1 and 2. Graph of (i) Ascorbic Acid (AA) Concentraticn - • ; (ii) Ascorbic Acid-utilization (AA-U)-0 ; and (iii) A Lxin rcr tu t -A;
of the shoot apex and of different parts of the developing spike of Norrral d~y unvernalized (NDC) and Long da)-verraliz'd (lLV) p!Hts cf N.P. 710 (Tritium aestivum)-(V) Vegetative apex, (R) reproductive arcx, (S) spikekts, (A) andrce-eium, (G) gynoeandrce-eium, (F) anthesis, (K) kerneL (E) embryo.
(3) The ascorbic acid concentration continues to maintain a high level almost right up to the stage of anthesis and thereafter begins to decline reaching an extremely low level in mature embryo. A similar trend is observable in all the three varieties under NDC treatment with the only difference that the period of high AA-concentration appears to get shortened in the case of very late flowering winter variety Hansa compared to the early flowering N.P. 710 presumably on account the higher range of temperature obtaining during the differentiation of the spike in the former.
?
•
...
FIG 3. 'lJ~ Rl VA L-NDC <( <(75
I ® ®
®
•
¢
(!:5 '•
6 ' ~ z
®
~
x
~
::>so 5 <(
4
~®
'43
i ..
?5
2 ·2
•
•
+ -4-+ +•
+ •IV R SS AG F K 0 0
0 so 100 I 50 200
::>
I Fl G 4.
<( <( RIVAL-LDV
<( <(
75
~
li
®
•
I~®
~f5
·~~~
::>50 5
~
<(4 @• I '4
@
3 •3
25
2 •2
••
+ + + + t•
•
•I VR SAG F K K E0 0
0 50 100 ISO
GROWTH PERIOD- DAYS
Fros. 3-4. Figs. 3 and 4. Graphs of: (i) Ascorbic Acid (AA) Concentra-tion-e; (ii) Ascorbic Acid-utilization (AA- U)-0; and (iii) Auxin content-A; off e shoot apex and different parts of the developing spike of normal day-unvernalized (NDC) and long day-vernalized (LDV) plants of Rival (Triticum aestivum). (V) Vegetative apex, (R) reproductive apex, (S) spikelets, (A) andrctcicm, (G) gynoecium, (F) anthesis, (I<) kernel, (E) embryo,
5 mg. in the case of the winter variety Hansa. Similarly the maximum level of AA-concentration in N DC plants also progressively rises with the lateness in time of flowering from 4·4 mg. in N.P. 710 to 6·6 mg. in the very late winter variety Hansa, with the late spring variety Rival occupying an intermediate position at 5 · 2 mg. of AA per g. fresh weight of the shoot apex. Mean ascorbic acid content of the shoot apex (Table I) also shows a rising trend with progressive lateness in flowering of a variety. It appears that both for the vegetative (v) as well as the reproductive (r) differentiation in the very late winter variety Han sa a higher threshold value of AA is necessary.
(5) Under vernalization and long photoperiod plants of all the three varieties reach the stage of 'square ridges' and then 'double
178 J. J. CHINOY
?
<t <t
<( <( 75
(!)
•
6
50 5 4
3.
25 2
0 0
50 100 150 200
=>'
.:.: <( HAN SA- LDV ~
<( <(
75 ~~·I 0 I
..
(!)
•
Fl G 6 • ' IJ ~ z
6
~
~
x
#
::::>50 5
~
(!) <(
.
4'
•43 ®e ·3
25
2 ·2
+ "' + + •I
SAG F
0 0
0 50 100 ISO 200
GROWTH PERIOD· DAYS
Fros. 5-6. Figs. 5 and 6. Graphs of: (i) Ascorbic Acid (AA) Conrentraticn
-e; (ii) Ascorbic Acid-utilization (AA-U)-0; and (iii) Auxin content-A.; of the shoot apex and different parts of the developing spike of normal day-unverra-Jized (NDC) and long day-vernalized (LDV) plants of Hansa (Triticum aativum). (V) Vegetative apex, (R) reproductive apex, (S) spikelets, (A) androecium, (G) gynoecium, (F) anthesis, (K) kernel and (E) embryo.
ridges' earlier. The period of vegetative differentiation ( v) of LDY plants also increases progressively from the early flowering variety to the very late winter variety Hansa as in the case of NDC plants (Table II).
TABLE I
Mean ascorbic acid content and its utilization as well as mean auxin content during periods of vege_tative and reproductive
differentiation
Variety
-N.P. 710 Rival
Hansa NDC LDV NDC LDV NDC LDV
Ascorbic Acid v* 2·3 2·1 2·7 2·3 3·5 2·7
r* 3·7 3 ·I 4·6
3·6 5·8 5·0
AA-Utilization v 33 40 41
45 50 48
r 58 65 71
69 78 75
Auxin Content v 0·27 0·23 0·25
0·22 0·21 0·11
r 0·21 0·15 0·20
0·16 0·22 0·16
* v, the period of vegetative differentiation; r, the period of reproductive differentiation. NDC, Normal day-unvernalized; LDV, Long day-vernalized.
Rival from Svalof. Mean AA-concentration is also significantly higher in Hansa in both the periods of vegetative (v) and reproductive (r) differentiation compared to the other two varieties (Table 1).
(7) The period of low AA-concentration (before the change in the shoot apex) is reduced in all the three varieties under LDV treat-ment. The AA-concentration remains low for 15, 25 and 65 days respectively in LDV plants of N.P .. 710, Rival and Hansa. This indicates that although the AA-concentration registers an earlier rise in the shoot apices of plants of all the three varieties under vernaliza-tion and long photoperiod the varietal order remains the same as in the case of NDC plants.
(8) The maxima of AA-concentration attained by all varieties are also lower in LDV plants compared to the corresponding NDC ones of the same variety. Here again the varietal differences in maxima persist. Thus the maxima of AA-concentration in LDV plants of N.P. 710, Rival and Hansa are 3·75, 3·90 and 5·20mg./g. fresh weight respectively.
!!SO L J. CHINOY
TABLE II
Periods (in days) of vegetative (v) and reproductive (r) differentiation as well as periods (in days) elapsing from transplantation to
the initiation of different organs in the spike
Variety
N.P. 710 Rival Han sa
NDC LDV NDC LDV NDC LDV
----·~--- ·-·--·---- ~-
---·-~---Period of Vegetative
Differ-entiation (v) 30 23 75 35 130 80
Period of Reproductive
Differ-entiation (r) 63 36 52 38 35 39
Period (days) between Trans-plantation and Initiation of:
(i) Glume 37 26 88 41 137 88
(ii) Lemma 43 32 97 50 142 92
(iii) Stamen 45 34 101 54 145 94
(iv) Carpel 50 36 105 56 146 95
(v) Ovule 52 38 108 58 148 95
- -
-NDC, Normal day-unvernalized; LDV, Long day-Hrr;aliz(,d,
Effects of varietal, photoperiodic-vernalization treatments, periodic readings as well as their interactions are highly significant. Even the second-order interaction, viz., variety-treatment-periodic reading
is highly significant.
Utilization of Ascorbic Acid
The data for the utilization of ascorbic acid in the growing apex are also presented in Figs. 1-6 and mean values for periods of vegetative and reproductive differentiation in Table I.
( 1) The AA-utilization (AA- U) curve also shows a rising trend in NDC and LDV plants of all the three varieties. During the period of high AA-concentration AA-utilization also remains at a high level falling off only after anthesis in all cases.
TABLE IlJ
Analysis of l'ariance of ascorbic acid contents of the shoot apex
Factor
A. Replicate
B.
Treatment C. VarietiesD. Periodic Readings (8 X C) Treatment x Varieties (8 X D) Treatment x Periodic
Readings
(C X D) Varieties X Periodic Readings
(A X D) Replicate X Peri odic Readings
(B x C x D) Treatment x Variety X
Periodic Readings Error
*
Indicates values significant at 1Degrees of freedom
2 25
2
25
50
25
50 130 311
per cent. level.
Variance Value of F
0·38 5·0
53·49 706· 5* 80·93 1069·1
*
l [. 79 155·8*2·52 33 ·2*
I ·38 18·2*
4·01 53·0*
0·02 0·3
[. 76 23·2*
0·08
---(3) Under LDV treatment the high level of AA-utilization (AA-U) is reached earlier in all the three varieties together with high AA-con-centration (Figs. 2, 4, 6). Here the degree of shift does not depend so much upon the time of flowering of the variety as in the case of AA-concentration. Thus for instance all the three varieties begin to utilize 45-50 per cent. of total ascorbic acid within 20-30 days after planting. However, the ascorbic acid utilization is significantly higher during the period of reproductive differentiation (Table I).
182 J. J. CHINOY
(5) Analysis of variance of data for AA-utilization was carried out for 312 combinations of 26 periodic readings, 3 varieties, 2 photo-period-vernalization treatments and 2 replicates. The analysis is presented in Table lV.
TABLE IV
Analysis of variance on data of ascorbic acid-utilization in the shoot apex
Degrees Value
Factor of Variance of
freedom F
A. Replicate 5·6 0·4
B. Treatment 1 2990·8 224·9*
c.
Variety 2 15787·7 1187·0*D. Periodic Readings 25 2597·4 195·3*
( B x C) Treatment X Variety 2 1487·3 111·8*
(B x D) Treatment x Periodic
Readings 25 660·0 49·6*
(C x D) Variety x Periodic Readings 50 965·4 72·6*
(Ax D) Replicate X Periodic
Readings 25 17·9 1·3
(B x C x D) Treatment x Variety x
Periodic Readings 50 368·5 27·7':'
Error 130 13·3
311
* [ndicates values significant at 1 per cent. level.
Effect of photoperiod-vernalization treatment, varieties, periodic readings and their first-order interactions on AA-utilization are highly significant. Here also the second-order interaction : variety-treat-ment-periodic reading is highly significant.
Auxin Concentration of the Shoot Apex
apex are also graphically presented in Figs. 1-6 and mean values in Table I. Main results are as follows:
(1) Auxin concentration is generally at a higher level in the begin-ning of the growth period up to the stage of square-ridges. There is a gradual decline in the auxin content almost up to the stage of anthesis and thereafter it continues to increase in the developing grain. From the milk-ripe stage onwards, there is a steep decline in auxin activity right up to the full ripe stage. There is hardly any auxin activity in an air-dry embryo of wheat. This is very significant because it snychronizes with almost complete absence of ascorbic acid in air dry seed.
(2) The general trend of auxin activity is similar in all the three varieties. The relatively higher auxin activity synchronizes with a relatively lower AA-concentration and AA-utilization and a relatively lower auxin activity parallels high AA-concentration and high level of AA-utilization.
(3) Analysis of variance was carried out on the data of auxin concentration and all the single factor as well as interaction effects were found to be highly significant. The analysis is not presented here.
Development, Ascorbic Acid Concentration and Its Utilization
From the foregoing it appears that the degree of development attained by plants of three wheat varieties under different treatments depends upon the level of AA attained by cells of the shoot apex as well as upon the amount of AA utilized by them in a given period of time. With a view to confirming this relationship regression of deve-lopment on the product of AA and AA-utilization was determined. The analysis is presented in Fig. 7. The regression coefficient is highly significant (value of t
=
19·9 at p=
0·01). The position of the regression trend line was determined by using the regression equation shown in Fig. 7. The relationship appears to be significantly valid.Metabolic Index
Considering the close correspondence between development and AA. A A-U product it can be assumed that the latter plays a major role in the metabolic activity of cells in the shoot apex. It, therefore, follows that:
The metabolic index I111 ex: C. U., and
li-:4 J. J, CH!NOY
0 /
0
0 Ell
•
0.
XIP
"'
0.
Ell"
• Gl ED1- ~,pl
z . . ~ 0
w ®xa~
o
~0
~"'
:t
...J
w Ell
> w o.,
X=O·I62Yt IS•2
0+---.---r--0 250 500
(Y) A A. - A A-U. PRODUCT
FIG. 7. Linear regression of development on ascorbic acid (AA) (·) ascorl:ic acid-utilization (AA-U) product. Key to the graph same as in Fig. 8.
per cent of the growth period up to the stage of first an thesis. Tt remains to be seen whether the metabolic index has any relation to cellular activity occurring in the shoot apex. For this purpose two types of growth estimates were made, viz., (l) number of organizer centres (indicating meristematic activity) and (2) cell elongation; the former from periodic data of tiller and leaf production and formation of different organs in a spikelet of the spike; and the latter from measure-ments of height and spike length. Paucity of space precludes the presentation of full growth data here, which has been adequately dealt with in earlier communications from this laboratory (for references please see Chinoy eta!., 1959). Few mean values of growth data are presented in Table V as indicative of the trend.
Linear regression of the number of organizer centres upon the metabolic index· was determined and using the coefficient of regression (which was highly significant: t
=
28 · 3 at p=
0 · 01) the formula giving the relationship between organizer centres and metabolic indices was determined. Substituting di q-erent values of metabolic index the corresponding theoretical value for the number of organizer centres was calculated and these values were used to determine the position of the trend line.!85
TABLE V
Maximum tiller number per plant, number of leaves on the main shoot and number of spikelets in the spike of the main shoot
Variety
N.P. 710 Rival Hansa
NDC LDV NDC LDV NDC LDV
-Max. Tiller No ... 8·5 5·0 21·0 6·5 40·0 12·0
Max. Leaf No.
..
7·5 7·5 15·0 7·5 18·5 12·3No. of Spikelets 20 11 23 15 22 18
-NDC, Normal day-unvernalized: LDV, Long day-vernalized.
Similar relationship between metabolic index and cell elongation is shown in Fig. 9. Here also regression of cell elongation on meta-bolic index was worked out and from the resulting regression equa-tion statistically computed values of ceil elongaequa-tion were obtained for marking the position of the trend line. Values for metabolic indices are plotted against the corresponding values of ceil elongation. It is clear that the experimentaiiy determined values of ceil elongation closely correspond with the trend line.
0
... o ><"'
...,_
NDC lOV !\J.P. 710
RIVAL G)
HANSA ~
•
XI 2 3 4
METABOLIC iN DE X (Y)
186
0
~1
><L{)
~I'-z
0 i=
<( l'J0
ZLfl
0
_ j
w
--'"'
- ' N
w
u
J. J. CHINOY
•
0•
00 Ell .0 @X X 0
0
.
.x
y. Ell•
Ell0
Ell
0 GJ
•••
0 X')(•
0 tile t• )( .x Ell
Ell
0
X"" 19·2Y-0•7
2
METABOLIC
3 4
I NDE X (Y)
5
FIG. 9. Linear regression of cell elongation on metabolic index. Key to the graph same as in Fig. 8.
Mean increment (per week) in the laying down of organizer centres as well as in cell elongation both in the stem and the spike together with the mean metabolic index during periods of vegetative and repro-ductive differentiation are presented in Table VI for the three wheat varieties under NDC and LDV treatments. There is a tremendous rise in cellular activity (as shown both by the laying down of organizer centres as well as cell elongation) during the period of reproductive differentiation paralleled by a considerable increase in the value of metabolic index.
From the foregoing it will appear that intensification of metabolic activity is brought about by the increasing concentration of ascorbic acid as well as its utilization in the shoot apex and this results in an outburst of tremendous growth activity in the cells of the shoot apex.
GENERAL DISCUSSION
Experimental evidence pertaining to the role of ascorbic acid metabolism in plants has been reviewed elsewhere (Bonner, 1957; Aberg, 1958; Mapson, 1958, 1960; Knox, 1960; Tonzig and Marre, 1961). It is, therefore, not necessary to discuss the question in detail here except in relation to growth and development.
Synthesis and Function of Ascorbic Acid in Plants
TABLE VJ
Mean increment per week in organizer centres as well as in cell elongation both in the stem and the spike and mean
metabolic index during periods of vegetative
Organizer Centres
and reproductive differentiation
Variety
N.P. 710 Rival Han sa
NDC LDV NDC LDV NDC LDV
v* 13 10 33 10 46 19
r* 158 118 112 102 154 159
Cell Elongation in
Stem v 3 · 5 1 4 · 0 2 · 6 8 · 8 1 · 9 3 · 6
r 10·8 20·0 11·9 14·2 13·2 13·5
Cell Elongation in
Spike v 0·6 2·5 0·3 1·3 0·2 0·1
Metabolic Index
r 17·7 31·6 16·2 19·2 19·5 19·9
v
r
0·34
I ·28
0·39
1·60
0·46
2·80
0·41
1·81
0·86
4· 17
0·67
3·39
*
v, the period of vegetative differentiation; r, the period of reproductive differentiation; NDC, Normal day-unvernalized; LDV, Long day-verl'alized.18~ J. J. CHINOY
Nanda, 1959; Michniewicz, 1961 a). There is also evidence in support of the fact that AA-concentration rises earlier in the embryo subjected to vernalization treatment (Chinoy and Nanda. 1959; Michniewicz,
1961).
With the emergence of the shoot above ground the relationship between AA and photosynthesis assumes great importance. A close relationship between photosynthesis and AA formation has been postulated by a number of workers (Giourd, 1938; Reid, 1938; Aberg, 1945, 1957; Prokoshew and Dantsheva, 1946; Popovskaja, 1950; Smith, 1952; Haagen. 1953; Asselbergs, 1957; Marre and Arrigoni. 1958; Mitsui and Oi. 1961 b).
Arnon and his co-workers (1954, 1956, 1957, 1958) have ascribed the role of an electron carrier to AA in the oxidative chain of photo-synthetic phosphorylation in illuminated chloroplasts or as a factor stabilizing the activity of the chloroplasts. Mitsui and Oi (1961 a)
have found significant correlations between seasonal variations in photosynthetic phosphorylation, photochemical transphosphorylation, the Hill reaction of spinach leaves harvested in different seasons and the ability of these leaf extracts to produce AA from its precursor.
The photosynthetic activity as well as carbohydrate content of leaves of the rice plant have also been shown to register a significant rise with the onset of flowering (Dastur and Chinoy, 1932). In a number of other crop plants the carbohydrate content of the leaves as well as of shoot apices (especially the invert sugar content) attained a significantly higher level in LDV plants compared to NDC ones (unpublished records). This is significant considering the fact that a higher concentration of sucrose or other sugars is essential for rapid biosynthesis of AA (Chinoy, Grover and Nanda, 1961).
Reid (1937, 1941 a, 1941 b, 1941 c, 1943) found ascorbic acid in the greatest concentration in actively growing zones of plants. In some experiments a sevenfold increase in AA content was registered in elongating root-tips. She suggested that this accumulation preceded and conditioned cell elongation.
A mass of data which has accumulated during the course of the last ten years in the writer's laboratory on the interaction between AA and auxin, exogenous application of AA to the shoot apices of plants, biosynthesis of AA by different organs of plants as well as on the AA-content and its utilization during the course of the growth period of different crop plants such as Triticum, Hordeum, Avena, Secale, Triticale (of Arne Muntzing), Cicer, Cajanus, Arachis, Linum and others grown under different photoperiodic and vernalization treatments also points conclusively to the important role of ascorbic acid in growth and development (Chinoy, Grover and Sirohi, 1957 a; Chinoy, Nanda and Garg, 1957, 1958; Garg, Chinoy and Nanda, 1958, 1958 a;
Evidence presented in the present paper lends full support to the fact that ascorbic acid is continuously being formed durieg germination as well as photosynthesis. Vernalization and long photoperiod help in raising the level of AA at an earlier date in the shoot apices of all the three varieties compared to the level of AA in the shoot apex of corresponding NDV plants. Evidence for the rising trend in the AA content of leaves and other organs with the approach of flowering has been obtained earlier in the writer's laboratory (Chinoy, Singh and Sirohi, 1958) as well as by other workers (Venkataramani. 1950; Zachershichina, 1960; Michniewicz. 1961; Shkol'nik and Abdura-shitov, 1962).
Enzymes and Ascorbic Acid Utilization
A number of enzyme systems capable of catalyzing the oxidation of ascorbic acid in plant extracts have been discovered: (I) those with indirect reaction, such as polyphenolase, laccase, cytochrome oxidase and peroxidase; and (2) that catalyzing a direct reaction between AA and oxygen, riz., AA-oxidase (see Bonner, 1957; and Mapson, 1958 for further discussion and references).
It may also be noted here that Sisakyan and Filippovich ( 1951, 1953) have demonstrated qualitative changes not only in the oxidative systems but also in dehydrogenase systems during vernalization, which suggests a correlation between redox systems in the plant with its development. From the data already reported (Chinoy, Grover and Sirohi, 1957 a; Chinoy, Singh and Sirohi, 1958) as well as those pre-sented in this paper (Figs. 1-7 and Table I) it appears that ascorbic acid is an important constituent of the redox system influencing develop-ment. Shkol'nik and Abdurashitov (1962) also ascribe a similar role to AA.
It will not be out of place to record here the Regulator-Anti-oxidant Hypothesis of Siegel and Porto ( 1961) which seeks to relate regulation of growth. development and aging of plants to the balance of oxidants (electron acceptors) and antioxidants (electron donors) in the cell tissue or organism.
Redox systems regulated by glutathione and others may also be involved in growth of some plants. It has, however, been postulated that retardation of AA-oxidation keeps the glutathione in a reduced form and as a consequence of which growth is accelerated (Marre and Arrigoni, 1957).
Relationship Between Ascorbic Acid. Nucleic Acids. Proteins and Growth
190 J. J. CH!NOY
processes. Further a parallelism has been established between higher AA-content and a higher DNA/RNA ratio in leaves during flowering (Zachershichina, 1960).
Although much remains to be elucidated with regard to the complex process of protein synthesis there remains little doubt that Ribonucleic acids (RNA) are essential for protein synthesis (Brachet, 1952; Linderstr<f>m-Lang, 1952; Brown, Reith and Robinson, 1952; Mckee, 1952; Loomis, 1953; Ebert, 1954; Fruton, 1954; Lipmann, 1954; Brown and Robinson, 1955; Mirsky and Allfrey, 1957; Askonas, Simkin and Work, 1960; Hoagland, 1960).
Cell growth and differentiation in both the plant and the animal are inexorably bound with the formation and degradation of proteins and enzyme proteins. An increase in protein content takes place during certain stages of cell growth especially flowering (Kursanov and Bryushkova, 1940; Pearsall and Billimoria, 1937). Increased protein content is correlated with increased metabolic activity of the cells (Brown eta/., 1952). Prokoshew and Petrochenko (1948) as well as Kiryukhin (1961) have also established relationship between protein content and ascorbic acid in potato tubers.
Undoubtedly therefore the high ascorbic acid content as well as its rapid utilization in the actively growing and differentiating cells are of great significance when taken in conjunction with the known role of nucleic acids in protein metabolism, high nucleic acid content of actively dividing cells, as well as the relationship between protein content and the formation of new protoplasm. The induction and acceleration of growth and flowering by feeding the plant with nucleic acid derivatives and other substances which represent or simulate nuclear constituents or which drastically affect these constituents (Levan, 1939, 1951; Butenko, 1960; Chailakhyan eta!., 1961); as well as by the exogenous application of ascorbic acid (Havas, 1935; Havas and Gal, 1936; Dennison, 1940; Chailakhyan, 1956; Zemle-nookhin, 1956; Chinoy. Nanda and Garg, 1957, 1958; Garg, Chinoy and Nanda, 1958, 1958 a; Shkol'nik and Steklova, 1958) highlight the importance of AA formation and utilization in growth and develop-ment of plants in relation to nucleic acid-protein metabolism.
On the basis of the foregoing discussion the following "Ascorbic Acid-Nucleic Acid-Protein Metabolism Concept of Growth and Development in Plants" is being hazarded with the full realization of the gaps in our knowledge, but in the hope that it may prove useful in imparting some coherence to the various hormonal concepts of growth and development advanced so far.
Ascorbic Acid-Nucleic Acid-Protein Metaholism Concepr of Grmt•th and De1·elopment
Long-day plants.-ln long-day plants vernalization helps in raising the level of AA in the shoot apex earlier which appears to have an inductive effect upon the cells for continued greater formation of AA and transference of this enhanced ability to future generations of cells (Chinoy, Nanda and Garg, 1957). The leaves of vernalized plants when placed under a long photoperiod produce photosynthetic material for a prolonged period and simultaneously give rise to AA. Oxidizing agents of AA are also produced under the influence of light. However the balance is in favour of increasing AA-concentration under suitable temperature and photoperiod and consequently the process of aging (maturation) of cells is retarded. The plastic materials are drawn towards the organizer centre with higher AA-content (unpublished records). When the AA-content of the shoot apex reaches a critical level it initiates reactions in the cell leading to accelerated formation of nucleic acids. DNA/RNA ratio increases. An increase in protein content takes place as a result of increased production of nucleic acids. This increase in protein production necessarily implies increased metabolic activity of the cell during growth.
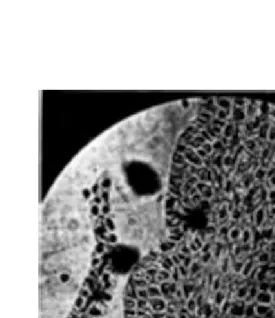
Such a state of acceleration of metabolic activity is rapidly trans-ferred to successive cell generations together with an improved ability for the biosynthesis of AA. A stage is reached when the metabolic activity increases to such an extent that two growth organizer centres develop where formerly only one had developed leading to the stage of square ridges and then double ridges (Fig. 10). Henceforward the shoot apex has entered the period of reproductive differentiation and the rate of formation of growth organizer centres is greatly accele-rated (Fig. 8 and Table VI). The primordia of different parts of the spike are laid down in rapid succession (Table VI). A stage in the rapidity of the formation of organizer centres, cell division and cell elongation (Figs. 8, 9, and Table VI) is reached when the chromosome material does not get sufficient time to grow and divide normally and thus meiosis results. Probably other changes such as that of viscosity are also involved (Tonzig, 1952; Tonzig and Trezzi, 1954; Chinoy, Grover and Sirohi, 1957 a; Tonzig and Marre, 1961).
192 J. J. CHINOY
growth and development the AA of the cell is immobilized. An indirect support for this contention is obtained from the work of Nanda and Hamner (1959); also see Hamner (1960) who have shown that when Biloxi Soybean a short-day plant is exposed to long photo-cycles at a comparatively lower temperature floral bud initiation takes place. At a higher temperature intervention of a dark period induces flowering in a short-day plant by reducing the oxidative processes and by accelerating AA formation and accumulation thus ensuring an adequate level of metabolic activity for flowering.
The known retarding action of auxin on flowering in short-day plants is probably an indirect one by stimulating oxidative enzyme systems. At the normal temperature of the experiment (which is above 20° C.) exogenous application of auxin may also bind the available free endogenously produced ascorbic acid giving rise to ascorbigen which has been found to be inactive so far from the point of view of growth and development (Basu, 1961).
The data presented in the present paper clearly indicate that the winter wheat Hansa requires a higher AA-concentration as well as a higher level of AA-utilization for the coversion of the shoot apex from the vegetative to the reproductive state compared with the cor-responding levels in N.P. 710 and Rival-both spring varieties. This is a very significant fact supporting the above trend of thought when we take into consideration the fact that vernalization of winter cereal (in this case winter rye Petkus) can be brought about as readily by darkness as by cold temperature (Purvis and Gregory, 1937). When winter wheat Hansa was vernalized and put under long photoperiod AA-concentration at the time of the change in the shoot apex as well as maximum AA-content were both appreciably lowered (Fig. 6) approaching those of spring varieties.
This Ascorbic Acid-Nucleic Acid-Protein Metabolism Concept for Growth and Development of Plant derives further substantiation from a mass of data showing close correlations between numerous growth characters, metabolic activity and flowering. 1 n fact the developmental process of a plant governs the magnitude and the rate of growth in its various components (see Chinoy eta!., 1959, for references).
Relationship between AA, AA-Utilization and Auxin
193
high level of its utilization. The auxin activity sharply increases after anthesis right up to milk-ripe stage and thereafter declines. During the period of grain development the high auxin activity is paralleled by lower AA-concentration and a decline in the level of its utilization. This close correspondence between auxin concentration on the one hand and AA-concentration and its utilization on the other leads one inevitably to the conclusion that the auxin activity of the extracts of the shoot apex may be essentially due to the action of ascorbic acid on growth.
It is difficult to visualize diverse pathways of induction and stimu-lation of growth and metabolic activity by regulatory substances of the plant as well as the synthetic ones. It is more probable that auxins and gibberellins function by stimulating the formation of ascorbic acid in leaves and growing regions of a plant (Chinoy and Nanda, 1959 and unpublished records; Michniewicz, 1961 a).
Discussing the metabolic reactions to auxin Marre and Arrigoni (1957) have postulated a scheme in which it has been shown that auxin inhibits AA-oxidation and thereby decreases the concentration of dehydro-ascorbic acid. This results in an increase in the ratio of reduced/ oxidised glutathione, which stimulates dehydrogenase activity and respiration, affects SH groups of enzymes and cofactors as well as influences transpeptidation reactions and protein synthesis resulting in growth.
Finally another important physiological action of auxin may be considered in the light of the present postulation. Tissue culture work has revealed that indole acetic acid brings about marked changes in the nuclear material, and that the action of auxins in growth is intimately concerned with nucleic acid metabolism. Thus at lower concentra-tions auxins favour the production of DNA while at concentraconcentra-tions optimum for growth greater production of RNA takes place. Marked increases in DNA are associated with active cell division and increased RNA content is associated with cell enlargement (Silberger, 1953; Si1berger and Skoog, 1953; Porter, 1953; Naylor, 1953; Naylor et al., 1954; Jablonski and Skoog, 1954; Skoog, 1954). In the light of what has been said above it appears highly probable that the changes in nuclear material ascribed to auxin may be due to the accelerated for-mation and utilization of ascorbic acid under the influence of auxin as already shown earlier.
In conclusion, therefore, it can be said that high ascorbic acid concentration and its rapid utilization at the time of the transformation of the shoot apex from the vegetative to the reproductive state and subsequently right up to the stage of first anthesis, the synchronous fall in the auxin content during this entire period of reproductive differentiation, the increased DNA/RNA ratio during reproductive differentiation, increased protein and carbohydrate contents of repro-ductive parts, considerable increase in the production of organizer
i94 J. J. CHINOY
centres as well as in cell elongation during reproductive differentiation and also the high metabolic index during reproductive differentiation constitute the broad experimental base upon which the Ascorbic Acid-Nucleic Acid-Protein Metabolism Concept for Growth and Deve-lopment of Plants rests; and further that, flowering is merely an accentuated phase of growth activity of a plant.
SUMMARY
Unvernalized and vernalized seed of three vanet1es of Triticum aestivum N.P. 710 (Indian early flowering variety), Rival (late spring variety) and Hansa (very late winter variety), both from Svalof, were grown under normal day (of 11 hr.) and long day (of 24 hr.). Besides the usual growth analysis the development of the spike was studied by making periodic measurements of different organs. From the periodic data of tiller and leaf numbers as well as from dates of organ initiation and development, weekly production of organizer centres were calculated for the period of vegetative differentiation (from trans-plantation to double ridges) as well as for the period of reproductive differentiation (from double ridges to first anthesis). Similarly cell elongation was determined by calculating the stem height and spike length on any given sampling day as per cent. of the maximum.
Weekly determinations of ascorbic acid concentration (mg. AA;g. fresh wt.), ascorbic acid-utilization (per cent. AA oxidised in 1 hr.) and auxin content (r.g.r. of Avena coleoptile section elongation) in vegeta-tive and reproducvegeta-tive shoot apices as well as in different organs of the developing spike were carried out.
Ascorbic acid concentration as well as its uti.ization increased and reached a significantly higher level at the time of the change in the shoot apex from the vegetative to the reproductive state. This enhanced formation and utilization of ascorbic acid continued throughout the period of reproductive differentiation (from double ridges to first anthesis). Synchrorwus with this the auxin level in the shoot apex reached a significantly lower level at the commencement of the period of reproductive differentiation and thereafter up to anthesis.
Production of organizer centres and cell elongation increased tremendously during this period.
development as the constant) were found to be highly significant. The present data are discussed in the light of known facts of nucleic acid-protein metabolism.
A new concept of growth and development, viz., 'the Ascorbic Acid-Nucleic Acid-Protein Metabolism Concept of Growth and Development in Plants' has been advanced on the basis of the high ascorbic acid concentration and its rapid utilization at the time of the change in the shoot apex from the vegetative to the reproductive state and also during the period of reproductive differentiation, the synchronous fall in auxin content, the increased DNA/RNA ratio during the same period, increased protein and carbohydrate contents of reproductive parts, marked enhancement in the production of organizer centres and in cell elongation as well as the high metabolic index during reproductive differentiation. In accordance with this concept the formation and utilization of ascorbic acid in the shoot apex of a plant stimulates the production of nucleic acids which are known to participate in the synthesis of proteins. With increased production of proteins protoplasm increases and growth activity results in the form of laying down of organizer centres, cell division and enlargement. Under suitable environmental conditions the formation and utilization of ascorbic acid is greatly accelerated and as a consequence of which nucleic acid-protein metabolism is also considerably stimulated resulting in a tremendous upsurge in the rate of formation of organizer centres, cell division and cell enlarge-ment. This brings about a profound change in the differentiation of the shoot apex. Ultimately the cellular activity is accelerated to such an extent in some cells that the chromosome material does not get sufficient time to grow and divide normally and thus meiosis results.
ACKNOWLEDGEMENTS
I wish to thank my associates K. K. Nanda, Narendra Singh, R.
Grover, G. S. Sirohi and 0. P. Garg for their help with experimental work.
DEDICATION
I dedicate this work to the memory of my revered teacher Professor F. G. Gregory.
REFERENCES
l. ABERG, B. (1945). Effects of light and temperature on the ascorbic acid co rt...:nt of green plants. Ann. Roy. Agri. Col/. Sweden, 13, 239-73.
CAssel-bergs, 1957.)
2. - - - - . (1949). Changes in the ascorbi~ acid content of darkened leaves as infbenced by temperature, sucrose application, and severing from the plant. Physiol. Plant., 2, 164-83.
196 J. J. CHINOY
4. ABERG, B. (1958). Ascorbic acid. Encyc/op. Pl. Physiol., 6, 479-99. EJ.: W. Ruhland, Springer-Verlag, Berlin.
5. ALI·ZADE, M. A. (1959). Dependence of nucleic acid content in tea plant on growth of buds and young shoots. Fizio/. Raste., 6(1), 48-52.
6. ARNON, D. I., WHATLEY, F. R. AND ALLEN, M. B. (1954). Photosynthesis by isolated chloroplasts. II. Photosynthetic phosphorylation, the con-version of light into phosphate bond energy. J. A mer. Chem. Soc .. , 76, 6324-29.
7. - - - . (1956). Photosynthesis by isolated chloroplasts. IV. General con· cept and comparison of three photochemical reactions. Biochem. Biophys.
Acta, 20, 449-61.
8. - - - - . (1957). Triphospho-pyridine nucleotide as a catalyst of photo-synthetic phosphorylation. Nature, 180, 182-85.
9. ··----. (1958). Assimilatory power in photosynthesis. Science, 127, 1026-34.
10. AsKONAS, B. A., SIMKIN, J. L. AND WORK, T. S. (1960). Protein synthesis in cell-free systems. In Proteins. Proc. IV Intern!. Congr. Biochem., Vienna. Ed.: H. Neurath and H. Tuppy, Pergamon Press, London, pp. 181-96.
II. As>ELBEROS, E. A. M. AND F~ANCIS, F. J. (1932). Stud:es on the forml· tin of v;t1mh C in slices of potato tissue. Canadian J. Bot., 30, 665. 12. AsSELBERGS. E. A. M. (1957). Studies in the form1tion of ascorbic acid i 1
dJtached 'apple leaves. Plant Physiol., 32, 326-29.
13. B<\RsKAYA, E. I. AND OKNINA, E. Z. (1959). Nucleic acids during the for-mltion and growth of seeds of fruit cultures. Fiziol. Raste., 6 (1), 98-99.
14. - - - . (1959 a). Role of nucleic acids in growth processes and their effect on the dormant state of buds of fruit plants. Ibid., 6 (4), 457-63.
15. BAsu, S. (1961). Studies on Ascorbigen. Thesis, Calcutta University for the Ph.D. degree.
16. BHARANI, S. P., SHAH, Y. S. AND SHRINIVASAN, A. (1953). Biosynthesis of vitamin C during germination. III. Effects of sugars, Klebs' inter-mediates, amino-acids. B-vitamins and correlation with biogenesis of nico-tinic acid. Proc. Indian Acad. Sci., 37 B, 33.
17. - - - - . (1953 a). Biosynthesis of vitamin C during germination. IV. Eff..:cts of mitotoxic agents and certain enzyme inhibitors. Ibid., 37 B,
54-71.
18. BONNER, W. D., JR.. (1957). Soluble oxidases and their functions. Ann. Rev. Pl. Physiol., 8, 427-52.
19. BRACHET, J. (1952). The role of the nucleus and the cytoplasm in synthesis and mJrphogenesis. In structural aspects of cell physiology. Sym. Soc. Exptl. Bioi., No. 6, pp. 173-200.
20. BREGETOVA, L. G. (1952). Co.1t~nts of ascorbic acid in the lewes of cotton. Bioi. Abstr., 7202, 26, 679.
21. BROWN, R., REITH, W. S. AND ROBINSON, E. (1952). Mechanism of plant cell growth. In structural aspects of cell physiology. Sym. Soc. Exptl. Bioi., No. 6, pp. 328-47. Academic Press, Inc., New York.
22. - - - AND ROBINSON, E. (1955). V. Cellular differenti<ltion and the dJvelo;JmJnt of enyme proteins in pllnts. In Biological Specificity and Growth. Ed.: E. G. But:er, No. 5, pp. 93-118, Pri 1ceton University Press,
"
23. BuTENKO, R. G. (1960). Application of the method for cultivation of isolated terminal buds in a study of growth and organogenesis in plants. Fiziol.
Raste., 7(6), 715-23.
24. C~ \ILAKilYAN, M. KH., BUTENKO, R. G. AND LYUBARSKAYA, J. J. (1961). Effect of derivatives of nucleic acid metabolism on the growth and flowering of Perilla nankinensis. Ibid., 8 (1), 101-13.
25. CHAILAKHYAN, M. KH. (1956). Dok!. An. S.S.S.R., 3 (4), 894.
26. CHINO\', J. J. (1945). A simple micro-macerator for plant analysis. Curr. Sci., 14, 102-03.
27. - - - - . (1956). Determination of photothermic and vernalization quanta for the vegerative period of wheat. Physiol. Plant., 9, 1.
28. --AND NANDA, K. K. (1951). Effect of vernalization and photoperiodic treatments on growth and development of crop plants. I. Varietal
dif-ferences in flowering of wheat and its correlation with spike growth, under varying photoinductive and post-photoinductive treatments. Ibid., 4, 209.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
n
--,GROVER, R. AND SIROHI, G. S. (1957). Bio-assay of auxins on the basis of relative growth of Avena coleoptile sections. Ibid., 10, 86.
- - - - . (1957 a). A study of the interaction of ascorbic acid and indole-3-acetic acid in the growth of Avena coleoptile sections. Ibid., 10, 92.
- - , NANDA, K. K. AND GARG, 0. P. (1957). Effect of ascorbic acid on growth and flowering of Trigone!! a foenum-graecum and Bra;sica chinensis. Ibid., 10, 869.
- - - - . (1958). Effect of ascorbicacid on growthand flowering of Tri-gonella foenum-graecum and Brassica chinensis. Proc. Seminar on Modern
Developments in Plant Physiology, University of Delhi, pp. 43.
- - , GRoVER, R. AND NANDA, K. K. (1958). Sucrose as precurso of the regulatory sbstances in Avena coleoptile. Ibid., pp, 13,
- - . NARENDRA SINGH and SIROHI, 0. S. (1958). Interaction of ascorbic acid with indole-3-acetic acid in flowering of wheat. Ibid., p. 46.
- - NANDA, K. K., S!ROHI, G. S. AND SAWHNEY, K. L. (1959). Growth and phasic development of wheat. I. Vegetative period and photothermic requirement. Indian J. Plant Physiol., 2, 29-4 5.
_ _ AND - - . (1959). Auxin catalized biosynthesis of ascorbic acid by dry-excised embryo of wheat. Proc. IX. Intern!. Bot. Congr., Abstracts, 2, 69.
- - . GROVER, R. AND NANOA, K. K. (1961). Role of sucrose in the growth reaction of Avena coleoptile sections. Flora Bd., 150, H. 1,
s.
1-10.DASTUR, R. H. AND CHINOY, J. J. (1932). Carbon dioxide assimilation of
the leaves of the rice plant. Indian J. Agri. Sci., 2, 431.
DENNISON, R. (1940). Growth response of plants to riboflavin and ascorbic acid. Science, 92, 17.
EBERT, J. D. (1954). Some aspects of protein biosynthesis in development In Aspects of Synthesis and Order in Growth. Ed.: Dorothea Rudnick. Pp. 69-112. Princeton Uni. Press, Princeton, New Jersey.
FRUTON, J. S. (1954). Th~ biosynthesis of proteins and peptides. In Aspects of Synthesis and Order in Growth. Ed.: Dorothea Rudnick, No. II, pp. 15-42.
GARG, 0. P., CHINO\', 1. J. AND NANDA, K. K. (1958). A study of the effect
]9g J. J. CHINOY
development of Brissica chinensis. Proc. Seminar: Modern Development.~
in Plant Physiology, Delhi Univ., pp. 20.
43. GARO, 0. P. (1958 a). I1teraction of ascorbic acid and colchicine on growth aJd d~velopment of Brassica chinensis L. Fyton, 10 (1), 17-25, Part Ill. 44. GIROUD, A. (1938). L'acide ascorbique dans les cellules et les tissues.
Protoplasma mongraf. 16. (Assdbergs, 1957.)
45. Gl\1VBR, R., CBINOY, J. J. AND NANDA, K. K. (1958). Growth-regulating action of ascorbic acid in Avena coleoptile sections. Sym. " The Recent Advances in the Study of Plant Metabolism," Univ. AllaJ-.;_o,tad, pp. 41.
46. GREGORY, F. G. (1917). Third Annual Report, Experimental and R:search Station, Cheshunt.
47. ---...:., (1921). Studies in the energy relations of plants. I. The increase in the area of leaves and leaf surface of Cucumis sativus. Ann. Bot., 35,
93.
48. - - - . (1926). The effect of climatic conditions on gro'>' th of barley. Ibid., 40, I.
49. - - - - . (1928). P'wsiological aspects of the cotton plant in Gezira. Rept. Meeting Res. Workers, Gezira Farm, Sudan.
50. - - - - . (1928 a). StCJdies in the energy relations of plants. II. The effect of tem;Jerature on i'lcrease in area of leaf surface and in dry '>'eigl.t of
c~~umis sativus. P-1rt I. The effJct of temperature on the increase in area of l-::-1f SJtf-1ce. Ann. Bat., 42, 469.
51. - - AND CROWTHER, F. (1928 b). A physiological study of varietoJ diff.Jrences h plants. I. A study of the comp<mltive yields of barley varieti<.s with
diff~rent manurings. Ibid., 42, 757.
52. - - , - - AND L\ MBERT, A. R. (1932). The interrelation of factors control-li'lg tl-te production of cotton under irrigation in the Sudan. J. Agric. Sci.,
22, 617.
53. HAAOEN, U. (1953). The d.:ily changes of vitamin C-content in foliage leaves. Biol. Abstr., 29, 945; Abstr. 2754, 1955.
54. H\M~ER, K. C. (1960). P\otoperiodism and Circadi<m Rhythms. In Bb·
logical Clocks. Cold Spring Harbor Symposia on Quantztative Biology, 25, 269-77. The Biological Lal::oratory, Cold Spring Hu·bor, L.. I., New York. 55. HAVAS, L. (1935). Ascorbic acid (Vitamin C) ar;d the germination Frd
gro'.'ith of seedlings. Nature (Lond.), 136, 435.
56. - - AND GAL, I. (1936). Divergent physiological effects of synthetic and natural ascorbic acid. Ibid., 138, 586.
57. HOAGLAND, M. B. (1960). Enzymatic reactions between amino-acids and ribJ JJc.eic acids as intermediate steps in protein syrctl-_esis. In Proteins. Proc. IV. Intern!. Congr. Biochem., Vienna, Ed.: H. Neurath and H. Tuppy, Pergamon Press, London.
58. [sHERWOOD, F. A., CHEN, Y. T. AND MAPSON, L. W. (1953). Synthesis of L--l>..:orbic acid in plants and animals. Nature (Lond.), l'il, 31,8.
59. - - - - . (1954). Synthesi~ of L-ascorbic acid in plants and animals. Biochem.
J., 56, 1.
60. JABLONSKI, J. R. AND SKOOG, F. (1954). Ce!l e' brgement <mdcell division in <:x.;ised tobacco pith tissue. Phy.1iol. Plant .. 7, 16.
62. KNOX, W. E. (1960). Ascorbic acid. Functions of ascorbic <icid in erz~rrc
systems. Vitamin metabolism. Ed.: Umbreit and H. Molitor. Proc. IV Intern!. Congr. Bwchem., Vienna, pp. 307. Pergamon Press, New Y01k.
63. KUI~SANOV, A. L. AND BRYUSHKOVA, K. (1940). Biokhymia, 5, 188. (Mckee,
1953J .
64. LEVAN, A. (1939). Cytological phenomenon associated with root swellirg caused by growth substancef. Hereditas, 25, 87.
65. - - - - . (1951). Chemically induced chromosome reactions in Allium cepa and Vrcia faba. Cold Spr. Harbor Symp. Quant. Bioi., 16, 233.
66. LINDERSTRj11M-LANG, K. U. (1952). Proteins and Enzymes. Stanford Uni. Press, Stanford.
67. LIPMANN, F. (1954). On the mechanism of some ATP-Jinked reactions and certain aspects of protein S)nthesis. In the Mechanism of Enzyme Action. Ed: McElroy and B. Glass, 599-604. Johns Hopkins Press, Baltimore.
68. LOOMIS, W. E. (1953). In Growth and Drfferentiation in Plants. An Intro-duction and Summary. pp. 1-17. Iowa State Coil. Press, Ames., Iowa.
69. MAPSON, L. W., CRUICHSHANK, E. M. ANO CHEN, Y. T. (1949). Factors affecting synthesis of ascorbic acid in cress seedlings. BiochEm. J., 45, 171.
70. - - , IsHERWOOD, F. A. AND CHEN, Y. T. (1954). Biological synthesis of L-ascorbic acid by plant mitochondria. Ibid., 56, 21.
71. MAPS0\1, L. W. (1958). Metabolism of ascorbic acid in plants. Part I. FL!nctio:J.. Ann. Rev. Plant Physio!., 9, 119--50.
72. - - - - . (1960). The biosynthes;s of L-ascorbic acid in plants and animals In Vitamin Metabolism. Ed.: Umbriet, W. and Molitor. Pergamon Press, London.
73. MARRE, E. AND ARRIGONI, 0. (1957). Metabolic reactions to Auxin I. The eff~cts of auxin on glutathione and the effects of glutathione on growth of isolated plant parts. Physiol. Plant., 10, 289.
74. - - - - . (1958). Ascorbic acid and photosyntr.esis. I. "Monodd.ydrc-a>corbic acid'' reductase of chloroplasts. Biodem. Biophys. Acta, 30, 453-57.
75. McKEE, H. S. (1953). Structure and synthesis of protoplasm. In Growth and D fferentiation in Plants. pp. 305-38. Ed.: W. E. Loomis. The Iowa State College Press, Ames., Iowa.
76. MECHANIC, F. Y. (1958). The factors affecting the amount of AA in the leaves. Bioi. Abstr., No. 6912, 28, 636.
77. MtCHNIEWlCZ, M. AND ROWICKA, K. (1961). Investigations on ascorbic acid content in spring and winter wheat in time of germination and early growth. Acta Agrcbotanica, 10 (2), 19-34.
78. - - - . (1961). The infbence of vernalization on the content of ascorbic acid in winter wheats. Ibid., 10(2), 119-31.
79. - - - - . (1961 a). The influence of auxin and gibberellin on the ascorbic acid c;:nte:J.ts in w'1eat during the period of germination and early growth. Bioi.
Abstr., 36 (14), 45935.
200 J. J. CHINOY
81. MITSUI, A. ANO 01, Y. (1961 a). Endogenous changes of photochemical activities of spinach leaves. Plant and Cell Physiology, 2, 45-50.
82. - - - - . (1961 b). Light induced formation cf ascorbic acid in isolated chloroplasts. Ibid., 2, 99-104.
83. NANOA, K. K. ANO HAMNER, K. C. (1959). The effect of temperature, auxins, antiauxins and wme other chemicals on the endogenous rhythm affecting photoperiodic response of Biloxi soybean. Planta, 53, 53-68.
84. NATH, M. C., CH!TALE, R. P. AND BHAVANI, B. l1952). Biosynthesis of vitamin C and a new precursor. Nature, 170, 545.
85. NAYLOR, J. M. (1953). Seen from Skoog (1954).
86. - - , SANDER, G. AND SKOOG, F. (1954). IAA-induced mitosisandcell enlargement without cell division in excised tobacco pith tissue. Physiol.
Plant., 7(1), 25.
87. PEARSALL, W. H. AND BILLIMORJA, M. C. (1937). 31, 1743. (Mckee, 1953 .)
88. PoPOVSKAJA, E. M. (1950). To the formation and translocation of ascorbic acid in plants. Biokhymia, 15, 249-55.
89. PORTER, W. K. (1953). Seen from Skoog (1954).
90. PROKOSHEW, S. M. ANO DANTSHEVA, E. I. (1946). Factors of ascorbic acid biosynthesis. Biokhymia, 11, 481-91.
91. - - AND PETROCHENKO. (1948). Interrelation of protein and ascorbic acid in potato tubers. Dokl. Nauk. SSSR, 61, 313-16.
92. PURVIS, 0. N. AND GREGORY, F. G. (1937). Studies in the vernalization of cereals: A comparative study of winter rye by low temperature and short days. Ann. Bot. (N.S.), 1, 569.
93. RAY S. N. (1934). On the nature of the precursor of the vitamin C in the' veg·Jtable kingdom. I. Vitamin C in the growing pea seedling. Biochem. J., 28, 996.
94. REID, M. E. (1937). Localization of ascorbic acid in the cowpea plant at different periods of development. Amer. J. Bot., 445-47.
95. - - - - . (1938). The effect of light on the accumulation of ascorbic acid in young cowpea plants. Ibid., 25, 701-11.
96. - - - - . (1941 a). Relation of vitamin C to cell size in the growing region of the primary root of cowpea seedlings. Ibid., 28, 410-15.
97. - - - - . (1941 b). Metabolism of ascorbic acid in cowpea plants. Bull. Torrey Bot. Club, 68, 359-71.
98. - - - - . (1941 c). Relation of temperature to the ascorbic acid content of cowpea plants. Ibid., 68, 519-30.
99. - - - - . (1943). Interrelations of calcium and ascorbic acid to cell surfaces and intercellular substances and to physiological action. Physiol. Rev., 23, 76-99.
100. SHKOL'NIK, M. YA. AND STEKLOVA. (1958). cs~en in Shkol'nik and Abdura· shitov, 1962).
VOL. V, PL. IV
FIG. l 0. Microphotograph of a longitudinal section of the shoot apex of wheat. Note the square ridges showing the initiation of two organizer centres on each ridge indicating the commencement of reproductive differentiation.