Obersteiner, Andrea (2017): Pollen associated microbiome and its relationship to pollution and allergens. Dissertation, LMU München: Fakultät für Biologie
Full text
Figure

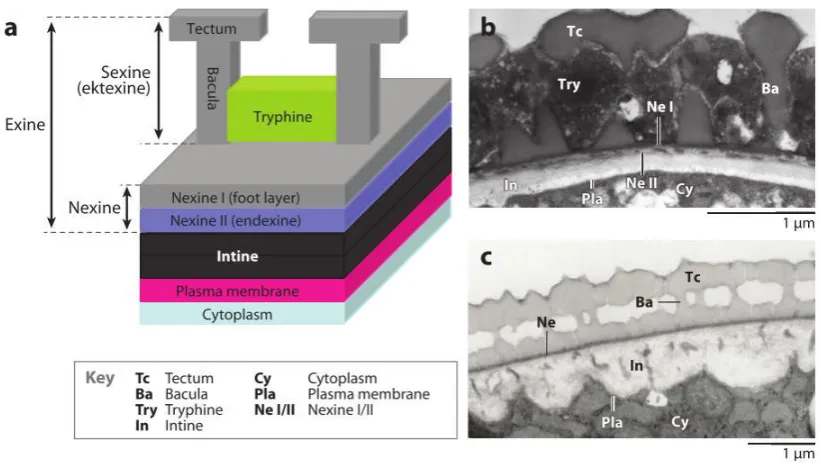
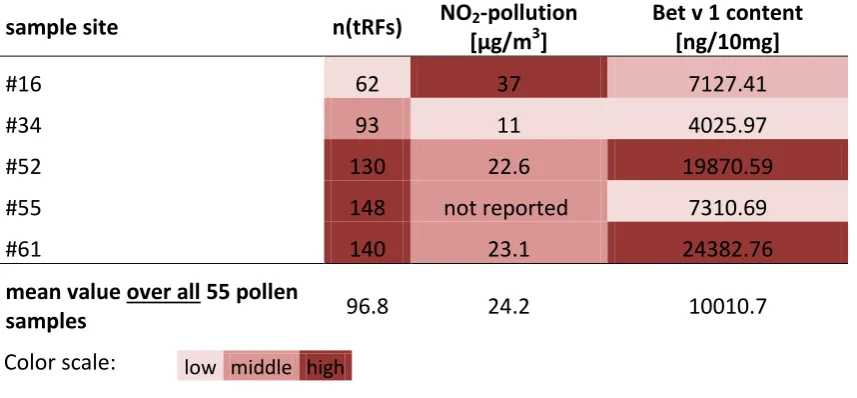

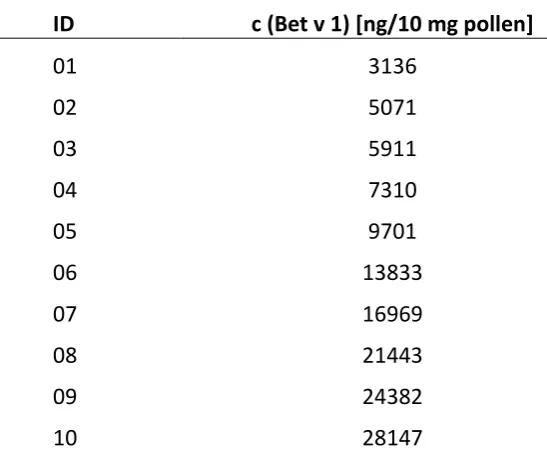
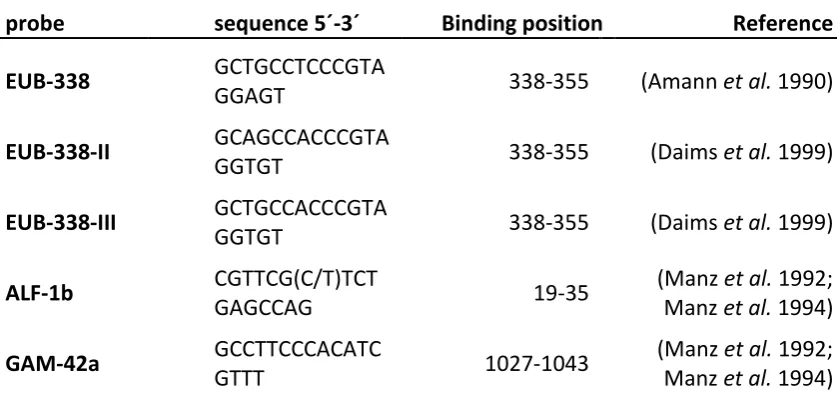
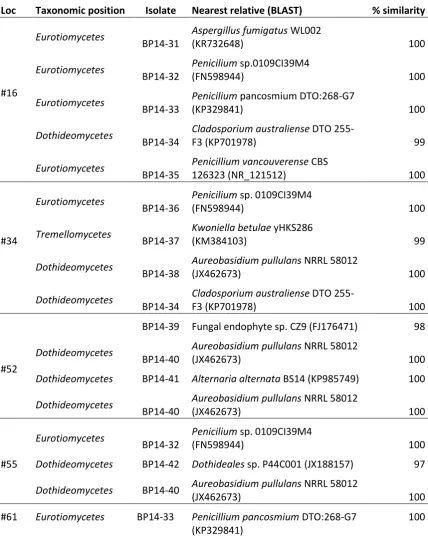


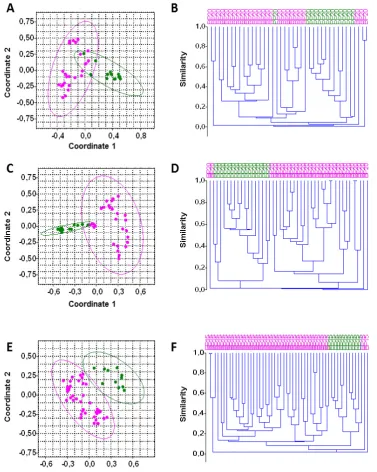
Related documents
caninum in large dairy farms in Isfahan Province, Iran, to realize if the parasite is an important agent to induce abortion in these herds and finally if infected
Results: When Caco-2 cell cultured with TO901317 alone, the mRNA levels of ABCA1, apoA1, apoM, LRH-1 and SHP1 were significantly increased with dose-dependent manners ( p <
Ten polypeptides of our SmD1 copurified protein set remain at the status “conserved hypothetical.” The fact that this set contained nearly all known trypanosome snRNP proteins and
time Event management is about planning, organization and execution of a project/activity such as parties, weddings, product launches, conferences, etc. Today, it
It has a number of pharmacological effects including antimicrobial, antioxidant, anti-cholinesterase, improvement of cognitive performance and mood, reducing
Legend: Comparison of abundances of the bacteria predicted to contain glutaminase gene (glsA, K01425) and urease gene subunits alpha (ureC, K01428), beta (ureB, K01429) and gamma
The solar PV system is simulated such that the PV module charge the battery through the controller and battery also provides the power to the load when the solar radiation
The result indicates a general decrease in liquid limit of the contaminated soil samples with the liquid limit of OPPCG samples having a decrease of 80.65% at 12%
