Copyright © 2003, American Society for Microbiology. All Rights Reserved.
emm
Typing of M Nontypeable Invasive Group A Streptococcal
Isolates in Israel
Allon E. Moses,
1* Carlos Hidalgo-Grass,
2Mary Dan-Goor,
2Joseph Jaffe,
1Ilanit Shetzigovsky,
1Miriam Ravins,
2Zinaida Korenman,
3Ronit Cohen-Poradosu,
1and Ran Nir-Paz
1Department of Clinical Microbiology and Infectious Diseases, Hadassah University Medical Center,1Department
of Clinical Microbiology, Hebrew University Medical School,2and Israel Ministry of Health Streptococcal
Reference Laboratory,3Jerusalem, Israel
Received 28 April 2003/Returned for modification 16 June 2003/Accepted 22 July 2003
We performedemmtyping of M nontypeable invasive group A streptococcal (GAS) isolates collected in a
prospective population-based study in Israel. One hundred twenty of 131 isolates (92%) hademmsequences
compatible with GAS, consisting of 51 differentemmtypes. Eleven isolates were found to be group G
strep-tococcus. Of the 120 isolates, 55 (46%) belonged to 32 types for which there were no typing sera available in the Streptococcal Reference Laboratory in Israel. The other 65 (64%) isolates, consisting of 19 types, had sera
available and therefore could have been serotyped. Forty-three isolates had T andemmtypes which were not
correlated according to standard M-typing protocols and were therefore missed. The principal effect ofemm
typing was the addition of 32 types not previously identified in Israel and the discovery of new associations
betweenemmand T types.emmtyping did not significantly change the proportion of M types; the five most
common types were 3, 28, 2, 62, and 41. Twenty different types comprised 80% of all isolates. No newemm
sequences were discovered.emmtyping emphasized the unusually low incidence of M1 strains causing severe
disease in Israel. As serological typing of GAS becomes more problematic due to lack of sera and the
appearance of new emmtypes, reference laboratories should replace M typing with emm sequence typing.
Development of a GAS vaccine relies on theemmtype distributions in different geographical locations. In our
study, 7% of isolates (types 41 and 62) are not included in a 26-valent vaccine that is being studied.
Group A streptococcus (GAS) causes a variety of human infections. These range from mild, self-limited diseases like pharyngitis and impetigo to severe, sometimes life-threatening illnesses such as bacteremia, necrotizing fasciitis, and toxic shock syndrome. Typing of GAS has long been the hallmark of both epidemiological studies and the understanding of diseases caused by different strains (2). M protein is a major virulence determinant of GAS that is associated with resistance to phagocytosis, adherence to cells, and virulence in a mouse model of necrotizing fasciitis (1, 24). Serological M typing was developed many years ago and was the only means for typing GAS. Initially, only 50 serotypes were described (12), but later several reference laboratories added some 30 more serotypes (12). This laborious method is becoming obsolete because it is time-consuming and expensive. Many centers have stopped producing specific antisera in rabbits. Sequence analysis of the hypervariable portion of the emmgene encoding M protein (emmtyping) has simplified GAS typing and has recently ex-panded the number of known GAS types from⬃80 to 124 (12). In various regions of the world, the percentage of M non-typeable strains varies from ⬎90% (20) to ⬍20% (15). The reasons for this variation include technical difficulties and a high prevalence of new emm types, for which serum is not available for M typing (15). In Israel, 67% of 21,517 GAS isolates (mostly from pharyngeal swabs taken over a 10-year
period) were M nontypeable (5). In a more recent study, Yagupsky and Giladi found 77% (10 of 13 cases) of GAS strains isolated from children with bacteremia to be M non-typeable (27). In a prospective population-based study of in-vasive GAS infections in Israel, conducted over a 2-year pe-riod, 133 of 409 cases (33%) were M nontypeable. These strains underwentemmsequencing and are the subject of this study.
MATERIALS AND METHODS
Source of GAS strains.GAS isolates were collected in a population-based study of invasive GAS done in Israel from 1997 to 1999 (19). Isolates were sent to our study center in Jerusalem by 24 of the 25 acute-care hospitals in Israel; 409 isolates were obtained from normally sterile sites and sent for traditional sero-logic M and T typing to the Israel Ministry of Health Streptococcal Reference Laboratory (MOH), and 402 strains were available for T typing. M typing was performed according to a decision analysis that relies on a known correlation between T and M types and which is similar to that used by Johnson and Kaplan (17). After T typing, only correlated M antisera were used for M typing. T nontypeable strains were not M typed. All strains that were M and T nontypeable were further analyzed byemmtyping (3).
emmtyping.PCR of streptococcal isolates was performed according to the recommendations of the Division of Bacterial and Mycotic Diseases, Centers for Disease Control and Prevention (CDC),Streptococcus pyogenes emmsequence database (http://www.cdc.gov/ncidod/biotech/strep/doc.htm). The primers used for amplification of GAS DNA were primer 1 (5⬘ TATTCGCTTAGAAAA TTAA 3⬘) and primer 2 (5⬘GCAAGTTCTTCAGCTTGTTT 3⬘).
According to the CDC recommendations the sequence of the sense strand of theemmhypervariable coding region was determined. The PCR product was sequenced by automated sequencing, using primer 1 (Hy Laboratory Ltd., Re-hovot, Israel). The sequence of bases 65 to 165 was submitted (using the Strep-tococcal Group A Subtyping Request Form, Blast 2.0 Server) to the National
* Corresponding author. Mailing address: Hadassah Medical Cen-ter, P.O. Box 12000, Jerusalem 91120, Israel. Phone: 9722-6776540. Fax: 972-2-6758915. E-mail: [email protected].
4655
on May 15, 2020 by guest
http://jcm.asm.org/
Centers for Disease Control Biotechnology Core Facility Computing Laboratory, where theemmtype was determined.
Correlation protocol.T type-M oremmcorrelation was based upon a preset protocol used by the MOH; this protocol is a combination of the common correlations in Israel and the correlations published by Johnson and Kaplan (17). We also compared T and M oremmcorrelation to the CDC protocol, which is based on both data from theS. pyogenes emmsequence database (http://www.c-dc.gov/ncidod/biotech/strep/emmtypes.htm) and recent publications (2, 4).
Differentiation between GAS and GGS.Differentiation between GAS and group G streptococcus (GGS) was achieved by subjectingS. pyogenesor Strep-tococcus dysgalactiaesubspeciesequisimilisto both a pyrrolidonyl aminopepti-dase test and a Rapid ID 32 STREP test (bioMerieux, Marcy l’Etoile, France). Statistical analysis.Statistical analysis was done with SPSS software (release 11.0.1). The2test or Fisher exact test was used for differences in proportions where required, and the Mann-Whitney U test was used for nonparametric comparisons. A two-sidedPvalue of⬍0.05 was considered significant. Two sets of comparisons were made between the isolates. In the first, we compared typeable and nontypeable strains. In the second we compared isolates bearing the same M oremmtype to evaluate if there are any parameters attributed to failure of M typing. The comparisons included demographics of patients, the clinical source of the isolate, and the month and year of the study. Comparisons were also made for strains from different regions in Israel by allocating patients to four distinct geographical regions.
RESULTS
emmtyping was performed on 131 of the 133 isolates which were nontypeable by M serotyping. The other two strains were not sequenced for technical reasons. All the isolates were PCR amplified, and the resulting sequences matched that of the
emmgene by homology analysis. The correspondingemmtype was received from the CDC facility. For 11 of 131emm-typed
isolates that were found to haveemmgenes characteristic of GGS, we repeated the serogrouping. Six of these isolates gave a positive reaction with group G antigen, indicating that the isolates were indeed GGS. The other five gave a positive re-action with the group A antigen. In contrast to this finding, both a pyrrolidonyl aminopeptidase test and a Rapid ID 32 STREP test confirmed that these five isolates were S.
dysga-lactiae. Thus, the five isolates were GGS presenting a group A
antigen.
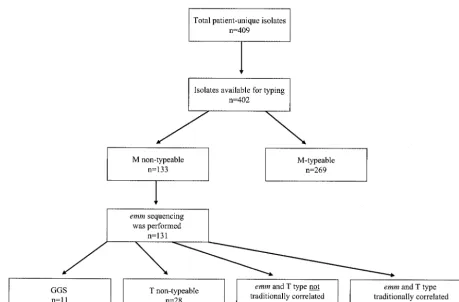
Among the 120 strains that hademmsequence typing com-patible with GAS (Fig. 1), there were 51 differentemmtypes (Table 1). Fifty-five of the 120 isolates belonged to 32 strain types for which there were no typing sera available in the MOH (types 30, 33, 42, 44, 51, 53, 61, 64, 65, 68, 71, 74 to 78, 81, 82, 84 to 87, 89, 92, 94, 95, 103, 113, 118, ST1815, ST3765, and ST5282). This was due to either newly characterizedemmtypes or known types for which antisera were no longer available at the MOH. The remaining 65 isolates, comprising 19 strain types (types 1, 2, 4, 5, 6, 9, 11, 12, 14, 18, 19, 22, 24, 26, 28, 29, 49, 59, and 60), for which sera were available, could have been serotyped by the conventional method. As shown in Fig. 1, for some isolates (49 of 120), the diagnosis should not have been missed, since the T and emmtypes were correlated. For an-other set of isolates (43 of 120), the association between the T and emmtypes did not agree with the protocol (Fig. 1), and therefore the strains could not have been M typed correctly.
[image:2.603.60.520.67.369.2]In the complete survey of 400 isolates, 84 had T-type and
FIG. 1. Number and distribution of GAS isolates in the study. “Traditionally” refers to M oremm types that correlate with the MOH scheme.
on May 15, 2020 by guest
http://jcm.asm.org/
TABLE 1. M andemmtypes and their corresponding T types
M/emmtype
No. of isolates No. of isolates for which M-correlates with T typeemm
New T types associated (no. of isolates) Total typedM a M NT
b⫺emm
typed CDC protocolsConforms toc MOH protocolsConforms to c T NT
1 9 5 4 6 6 0 8 (1), 28 (1), 55 (1)
2 28 26 2 23 28 0 6 (5)
3 104 104 0 84 104 0 1 (20)
4 10 4 6 7 8 0 12 (1), 15/17/23 (1), 28 (1)
5 9 6 3 9 9 0
6 5 3 2 2 4 1 2 (2)
9 9 6 3 7 7 0 15/17/23 (1), 28/56 (1)
11 6 3 3 6 6 0
12 11 10 1 10 11 0 11 (1)
13 4 4 0 4 4 0
14 8 2 6 1 4 3 3 (1), 49 (3)
15 5 5 0 0 5 0 15/17/23 (5)
18 9 4 5 4 5 1 5 (1), 6 (1), 12 (1), 15/17/23 (1)
19 5 3 2 0 3 2 8 (1), 15/17/23 (2)
22 7 1 6 5 5 0 15/17/23 (2)
24 1 0 1 0 0 0 15 (1)
25 1 1 0 1 1 0
26 6 0 6 0 0 5 15/17/23 (1)
27 4 4 0 4 4 0
28 49 41 8 45 48 0 4 (3), 49 (1)
29 5 4 1 0 5 0 28/56 (5)
30 1 0 1 0 0 1
33 2 0 2 1 1 1
41 13 13 0 11 13 0 1 (2)
42 1 0 1 0 0 1
44 1 0 1 1 1 0
49 2 1 1 2 2 0
51 1 0 1 1 1 0
53 1 0 1 0 0 0 28/56 (1)
55 1 1 0 1 1 0
59 4 0 4 3 2 1
60 1 0 1 0 0 0 12 (1)
61 2 0 2 2 0 0
62 18 18 0 17 18 0 5/27/44 (1)
64 1 0 1 1 0 0
65 1 0 1 0 0 0 15/17/23 (1)
68 1 0 1 0 0 0 28 (1)
71 1 0 1 0 0 0 14 (1)
74 3 0 3 2 1 1
75 4 0 4 3 3 1
76 4 0 4 3 3 0 4 (1)
77 3 0 3 2 2 1
78 3 0 3 2 2 0 5 (1)
81 4 0 4 1 1 2 49 (1)
82 1 0 1 1 0 0
84 1 0 1 0 0 0 11 (1)
85 2 0 2 0 0 2
86 2 0 2 2 0 0
87 2 0 2 2 0 0
89 1 0 1 1 0 0
92 2 0 2 1 0 1
94 1 0 1 0 0 0 28 (1)
95 1 0 1 0 0 1
103 2 0 2 0 0 2
113 1 0 1 0 0 0 3/13/B3264 (1)
118 2 0 2 1 0 1
ST1815 1 0 1 0 0 0 8 (1)
ST3765 1 0 1 0 0 0 3/13/B3264 (1)
ST5282 1 0 1 0 0 0 3 (1)
Group G 11 0 11 0 0 9 2 (1), 5 (1)
Total 400 269 131 279 318 37 (84)
aTaken from Moses et al. (19). bNT, nontypeable.
cThe M type oremmsequence correlates with previously known M- and T-type association according to traditional protocols (CDC or MOH).
on May 15, 2020 by guest
http://jcm.asm.org/
emm-M-type correlations different from those usually sug-gested by the CDC protocols (Table 1). For 41 of the 84 isolates, M typing was performed but the M-T correlations of the MOH were different from those used by the CDC (Table 1). The remaining 43 isolates were designated nontypeable and were therefore emm typed. Among the 120 strains, the five most commonemmtypes, accounting for 24% of the isolates, were types 28, 4, 14, 22, and 26. Interestingly, none of the strains wereemm3, indicating that M serotyping succeeded in identifying all M3 isolates. This was also true for a number of other types, including 13, 15, 25, 27, 41, and 62.emmtyping of nontypeable isolates did not significantly affect the overall pro-portions of the different M types: the five most common types before and afteremmtyping were 3, 28, 2, 62, and 41 (Fig. 2). These accounted for 75.1% of the M typeable isolates (n ⫽
269) but only 51.6% of the isolates typed byemmand M (n⫽
400). Twenty different types comprised 80% of all 400 isolates. There was no difference between the M andemmtypes with regard to the patient’s age, the month of isolation, the infected organ, or the hospital or region in Israel. This was also true for strains that should have been typeable according to their T types. Thus, these demographic characteristics could not pro-vide an explanation for the failure to M type these isolates. There was a tendency toward a higher proportion of nontype-able strains in the second year of the study (P⬍0.001; Mann-Whitney U test). The nontypeable strains did not correlate with other factors: age of patients, organ infected, hospital of isolation, or region in Israel.
We analyzed the T types of isolates and their correlations to demographical and clinical characteristics of patients. Interest-ingly, T types 3/13/B3264, 15/17/23, and 28/56 occurred mainly in the first year of the study (P ⬍ 0.001), while T type 3 occurred mostly in the second year of the study (P⬍0.001). Type 3/13/B3264 was also isolated more frequently in areas outside of the Jerusalem district (26.1 versus 11.3%; P ⫽
0.001). No correlation between T types and other parameters tested was found to be statistically significant.
DISCUSSION
We performedemmtyping of M nontypeable isolates col-lected in a prospective population-based study of invasive GAS
infections in Israel. The results of this study demonstrate that a considerable proportion of strains were considered M non-typeable because appropriate antisera were unavailable, either because sera had never been prepared (new serotypesemm82 and above [Table 1]) or because the sera were no longer available (e.g., M53 and M64). Technical problems in the pro-duction of high-quality sera or in serotyping may also affect the ability to M type isolates.emmtyping of GAS in Israel did not change the distribution of the five most prevalent types. The principal impact ofemmtyping was the addition of 32 new type strains to the Israel database and the discovery of new associ-ations betweenemmand T types. There were no newly discov-eredemmsequences among our strains.
The prevalence of M nontypeable isolates varies among dif-ferent studies and may be related to the geographical distri-bution of the types. In a report of 4,760 GAS isolates from Canada (26), 15.4% were nontypeable, while in the United Kingdom (7) only 7.35% of 16,909 strains were nontypeable. In one study in Thailand, 80% were found to be nontypeable (23). This may be due to the presence of newly characterizedemm
types. Indeed, there are several reports indicating that new
emmtypes appear at a higher rate in certain countries in Asia and Latin America (15, 16). The number of different M and
emm types (59 types) found in our cohort was considerably higher than that found elsewhere (10). This is a relatively high number of types, taking into account the small population (6,000,000) and the small area of Israel. This variation may be due to the large number of immigrants arriving in Israel during recent years, originating from widely separated geographical regions throughout the world. Among the four geographical areas in Israel, there was no difference in M or emm type distribution. In one study of invasive and noninvasive disease in Hong Kong, there were a total of 32 M types (13). In a study of throat isolates from children in Rome, there were 22 differ-ent emm types (8), while in Aachen, Germany, 216 isolates comprised 18 differentemmtypes (6). In a report from Thai-land, 40 invasive GAS strains were found to belong to 24 differentemmsequences (22). In a study of throat isolates and isolates from sterile sites in Mexico, there were 31 different
emmtypes (10). Similar to findings in the United States, where among 2,002 invasive GAS isolates the 5 most common types accounted for 49.5% of the isolates (21), in our study the 5 most frequent isolates (Fig. 2) accounted for 51.6% of all strains and 20 types accounted for 80% of the isolates.emm
types 3 and 28 belonged to the five most frequent isolates in both countries. In Israel, types 12, 41, and 62 were among the five most common types, whereas in the United States they were not present in the 10 most frequent types (21).
We could not find a correlation between M oremmtype and clinical features, such as patient gender, age, or source of infection. However, the power to detect these associations may be limited by the small number of isolates within the specific M types. This is in contrast to the findings of Tyrrell et al., who described clusters of M types as correlating with age (26). We could find no apparent reason for the higher rate of nontype-able strains in the second year of the study. There was no change in laboratory technique, and there was no difference between the frequencies of T nontypeable and T type M or
emmdiscordance in the 2 years of the study.
The unusually low incidence of M1 isolates causing severe
FIG. 2. Proportions of the five most prevalent GAS types after
emmtyping (M andemmtyping of 400 isolates).
on May 15, 2020 by guest
http://jcm.asm.org/
GAS disease in Israel (19) was corroborated byemm typing and is considerably lower than that described in many other geographical areas (9, 11, 21, 25).
The association of certain T types with specific M types is a well-known epidemiological observation. This association is the basis for using initial T typing for simplification of the M-typing process (17). We found correlations between T and M types which were previously not considered to associate with each other. This finding, together with unavailable sera at the MOH, explains why 60 of 131 isolates (46%) were M nontype-able. There are several notableemmtypes (2, 3, 14, 15, 18, 28, and 29) that were found to be associated with unusual T types (Table 1). The association of 20 M3 isolates with T1 is remark-able and is rarely seen in the United States (2, 4, 17). These M3 strains may represent former M1 clones and warrant further study.
Nine of the 11 GGS isolates in our collection were T non-typeable. Five of these 11 GGS isolates were found to have a group A cell wall carbohydrate antigen and group G strepto-coccalemmsequences, and two of them were T typeable. This phenomenon may be due to transfer of the group A (18) and T antigens from GAS toS. dysgalactiae. The population-based nature of our study may give an indication of the prevalence of this finding in Israel, if we assume that strains that were M typeable would not give a positive M-type result on a GGS.
emmsequencing seems to be a reliable means for distinguish-ing between GAS and GGS.
As serological typing of GAS becomes more problematic due to lack of sera and the appearance of new emm types, reference laboratories should replace M typing withemm se-quence typing. For the development of a vaccine for GAS, it is pertinent to know theemmtype distribution in different geo-graphical locations. In our study, 7% of strains (types 41 and 62) are not included in a 26-valent vaccine that is being studied (14).
ACKNOWLEDGMENT
This study was funded by the Chief Scientist grant of the Israel Ministry of Health to A.E.M.
REFERENCES
1. Ashbaugh, C. D., H. B. Warren, V. J. Carey, and M. R. Wessels.1998. Molecular analysis of the role of the group A streptococcal cysteine protease, hyaluronic acid capsule, and M protein in a murine model of human invasive soft-tissue infection. J. Clin. Investig.102:550–560.
2. Beall, B., R. Facklam, T. Hoenes, and B. Schwartz.1997. Survey ofemmgene sequences and T-antigen types from systemicStreptococcus pyogenes infec-tion isolates collected in San Francisco, California; Atlanta, Georgia; and Connecticut in 1994 and 1995. J. Clin. Microbiol.35:1231–1235.
3. Beall, B., R. Facklam, and T. Thompson.1996. Sequencingemm-specific PCR products for routine and accurate typing of group A streptococci. J. Clin. Microbiol.34:953–958.
4. Beall, B., R. R. Facklam, J. A. Elliott, A. R. Franklin, T. Hoenes, D. Jackson, L. Laclaire, T. Thompson, and R. Viswanathan.1998. Streptococcal emm types associated with T-agglutination types and the use of conserved emm gene restriction fragment patterns for subtyping group A streptococci. J. Med. Microbiol.47:893–898.
5. Bergner-Rabinowitz, S., and M. Ferne.1978. Type distribution of beta-hemolytic streptococci in Israel: a 10-year study. J. Infect. Dis.138:152–159. 6. Brandt, C. M., B. Spellerberg, M. Honscha, N. D. Truong, B. Hoevener, and R. Lutticken.2001. Typing ofStreptococcus pyogenesstrains isolated from throat infections in the region of Aachen, Germany. Infection29:163–165.
7. Colman, G., A. Tanna, A. Efstratiou, and E. T. Gaworzewska.1993. The serotypes ofStreptococcus pyogenespresent in Britain during 1980–1990 and their association with disease. J. Med. Microbiol.39:165–178.
8. Dicuonzo, G., G. Gherardi, G. Lorino, S. Angeletti, M. De Cesaris, E. Fiscarelli, D. E. Bessen, and B. Beall.2001. Group A streptococcal geno-types from pediatric throat isolates in Rome, Italy. J. Clin. Microbiol.39: 1687–1690.
9. Efstratiou, A.2000. Group A streptococci in the 1990s. J. Antimicrob. Che-mother.45(Suppl.):3–12.
10. Espinosa, E. L., Z. Li, G. D. Barreto, C. E. Jaimes, S. R. Rodriguez, V. Sakota, R. R. Facklam, and B. Beall.2003. M protein gene type distribution among group A streptococcal clinical isolates recovered in Mexico City, Mexico, from 1991 to 2000, and Durango, Mexico, from 1998 to 1999: overlap with type distribution within the United States. J. Clin. Microbiol. 41:373–378.
11. Facklam, R., B. Beall, A. Efstratiou, V. Fischetti, D. Johnson, E. Kaplan, P. Kriz, M. Lovgren, D. Martin, B. Schwartz, A. Totolian, D. Bessen, S. Holl-ingshead, F. Rubin, J. Scott, and G. Tyrrell.1999.emmtyping and validation of provisional M types for group A streptococci. Emerg. Infect. Dis.5:247– 253.
12. Facklam, R. F., D. R. Martin, M. Lovgren, D. R. Johnson, A. Efstratiou, T. A. Thompson, S. Gowan, P. Kriz, G. J. Tyrrell, E. Kaplan, and B. Beall.2002. Extension of the Lancefield classification for group A streptococci by addi-tion of 22 new M protein gene sequence types from clinical isolates: emm103 to emm124. Clin. Infect. Dis.34:28–38.
13. Ho, P. L., D. R. Johnson, A. W. Yue, D. N. Tsang, T. L. Que, B. Beall, and E. L. Kaplan.2003. Epidemiologic analysis of invasive and noninvasive group A streptococcal isolates in Hong Kong. J. Clin. Microbiol.41:937–942. 14. Hu, M. C., M. A. Walls, S. D. Stroop, M. A. Reddish, B. Beall, and J. B. Dale. 2002. Immunogenicity of a 26-valent group A streptococcal vaccine. Infect. Immun.70:2171–2177.
15. Jamal, F., S. Pit, R. Facklam, and B. Beall.1999. Newemm(M protein gene) sequences of group A streptococci isolated from Malaysian patients. Emerg. Infect. Dis.5:182–183.
16. Jamal, F., S. Pit, D. R. Johnson, and E. L. Kaplan.1995. Characterization of group A streptococci isolated in Kuala Lumpur, Malaysia. J. Trop. Med. Hyg.98:343–346.
17. Johnson, D. R., and E. L. Kaplan.1993. A review of the correlation of T-agglutination patterns and M-protein typing and opacity factor production in the identification of group A streptococci. J. Med. Microbiol.38:311–315. 18. Katsukawa, C., A. Tamaru, and Y. Morikawa.2002.Streptococcus dysgalac-tiaesubsp.equisimilispossessing Lancefield’s group A antigen. Kansensho-gaku Zasshi76:155–160.
19. Moses, A. E., S. Goldberg, Z. Korenman, M. Ravins, E. Hanski, and M. Shapiro.2002. Invasive group a streptococcal infections, Israel. Emerg. In-fect. Dis.8:421–426.
20. Nakashima, K., S. Ichiyama, Y. Iinuma, Y. Hasegawa, M. Ohta, K. Ooe, Y. Shimizu, H. Igarashi, T. Murai, and K. Shimokata.1997. A clinical and bacteriologic investigation of invasive streptococcal infections in Japan on the basis of serotypes, toxin production, and genomic DNA fingerprints. Clin. Infect. Dis.25:260–266.
21. O’Brien, K. L., B. Beall, N. L. Barrett, P. R. Cieslak, A. Reingold, M. M. Farley, R. Danila, E. R. Zell, R. Facklam, B. Schwartz, and A. Schuchat. 2002. Epidemiology of invasive group a streptococcus disease in the United States, 1995–1999. Clin. Infect. Dis.35:268–276.
22. Pimtanothai, N., P. Orataiwun, S. Nilgate, C. Suankatay, and P. Nunthapisud.2002.emmtypes of invasive group A streptococcal isolates from Thai patients at King Chulalongkorn Memorial Hospital from 1995– 1999. J. Med. Assoc. Thai.85(Suppl. 1):S371–S377.
23. Pruksakorn, S., N. Sittisombut, C. Phornphutkul, C. Pruksachatkunakorn, M. F. Good, and E. Brandt.2000. Epidemiological analysis of non-M-type-able group AStreptococcus isolates from a Thai population in northern Thailand. J. Clin. Microbiol.38:1250–1254.
24. Ravins, M., J. Jaffe, E. Hanski, I. Shetzigovski, S. Natanson-Yaron, and A. E. Moses.2000. Characterization of a mouse-passaged, highly encapsulated variant of group A streptococcus in in vitro and in vivo studies. J. Infect. Dis. 182:1702–1711.
25. Schwartz, B., R. R. Facklam, and R. F. Breiman.1990. Changing epidemi-ology of group A streptococcal infection in the USA. Lancet336:1167–1171. 26. Tyrrell, G. J., M. Lovgren, B. Forwick, N. P. Hoe, J. M. Musser, and J. A. Talbot.2002. M types of group A streptococcal isolates submitted to the National Centre for Streptococcus (Canada) from 1993 to 1999. J. Clin. Microbiol.40:4466–4471.
27. Yagupsky, P., and Y. Giladi.1987. Group A beta-hemolytic streptococcal bacteremia in children. Pediatr. Infect. Dis. J.6:1036–1039.