Diving animals frequently show flexibility or variability in their diving behaviour and performance. This variability can be observed within individuals (Beck et al., 2000), among individuals (Ropert-Coudert et al., 2003) or between populations (Tremblay and Cherel, 2003). The determinants of this variability are diverse. For example, variability in prey species may require a modification in dive profile (Takahashi et al., 2003), or, on a larger scale, temporal variability in the location of prey can lead to changes in foraging location and hence preferred diving depths (Charrassin and Bost, 2001). Some changes in diving behaviour have a physiological basis. Larger individual Magellanic penguins (Spheniscus magellanicus) tend to dive deeper and for longer than smaller birds (Walker and Boersma, 2003). The buoyancy, depth profile, stroking patterns and efficiency of dives by grey seals (Halichoerus grypus) and Weddell seals (Leptonychotes weddellii) are dependent partly on body condition (Beck et al., 2000; Sato et al., 2003). In both of these species, leaner animals tend to stroke less frequently and descend more rapidly than fatter, more buoyant animals and either dive for longer (Beck et al., 2000) or have a shorter recovery time at the surface between dives (Sato et al., 2003).
Green et al. (in press a) showed that the foraging and diving
behaviour of macaroni penguins [Eudyptes chrysolophus (Brandt 1837)] varies substantially throughout their annual cycle. Specifically, during the winter months, the penguins tend to dive to greater depths and for greater durations than during the summer. However, in an earlier study of their diving energetics, Green et al. (2003) showed that female macaroni penguins may dive close to the limits of aerobic metabolism during the summer months. Calculated aerobic dive limit (cADL) is a quantity often used to assess the energetic cost of dives (Butler and Jones, 1997). cADL is the time at which an animal is estimated to have exhausted its usable oxygen stores while submerged, and is calculated from the total usable oxygen stores divided by a measure of the rate of oxygen consumption (VO∑) while submerged. In macaroni penguins,
during the summer, cADL was estimated to be 138·s, and 95.3% of all dives were less than this threshold (Green et al., 2003). However, during the middle of winter, mean dive duration of foraging dives by female penguins was approximately 143·s (Green et al., in press a). This suggests either that the previous estimate of cADL was incorrect or that, during the winter months, the penguins were either subject to considerable ecological constraints that forced them to dive Published by The Company of Biologists 2005
doi:10.1242/jeb.01679
Macaroni penguins were implanted with data loggers to record heart rate (fH), abdominal temperature (Tab) and
diving depth during their pre-moult trip (summer) and winter migration. The penguins showed substantial differences in diving behaviour between the seasons. During winter, mean and maximum dive duration and dive depth were significantly greater than during summer, but the proportion of dives within the calculated aerobic dive limit (cADL) did not change.
Rates of oxygen consumption were estimated from fH. As winter progressed, the rate of oxygen consumption during dive cycles (sVO∑DC) declined significantly and mirrored the
pattern of increase in maximum duration and depth. The decline in sVO∑DC was matched by a decline in minimum
rate of oxygen consumption (sVO∑min). When sVO∑min was
subtracted from sVO∑DC, the net cost of diving was
unchanged between summer and winter. We suggest that the increased diving capacity demonstrated during the winter was facilitated by the decrease in sVO∑min.
Abdominal temperature declined during winter but this was not sufficient to explain the decline in sVO∑min. A
simple model of the interactions between sVO∑min, thermal
conductance and water temperature shows how a change in the distribution of fat stores and therefore a change in insulation and/or a difference in foraging location during winter could account for the observed reduction in sVO∑min
and hence sVO∑DC.
Key words: macaroni penguin, Eudyptes chrysolophus, diving, seasonal change, oxygen consumption, thermoregulation, cADL.
Summary
Introduction
Do seasonal changes in metabolic rate facilitate changes in diving behaviour?
J. A. Green
1,*, I. L. Boyd
2, A. J. Woakes
1, C. J. Green
3and P. J. Butler
11School of Biosciences, University of Birmingham, Edgbaston, Birmingham B15 2TT, UK,2Sea Mammal Research
Unit, Gatty Marine Laboratory, University of St Andrews, St Andrews, Fife KY16 8LB, UK and 3British Antarctic Survey, High Cross, Madingley Road, Cambridge CB3 0ET, UK
*Author for correspondence (e-mail: [email protected])
beyond their aerobic limits or else their physiological capability to dive increased.
In the present study, we compare diving behaviour and energetics during the summer/pre-moult foraging trip and winter/migratory period of macaroni penguins in order to determine what might cause the observed changes in diving behaviour. Specifically, we ask: (1) when do the observed large-scale seasonal changes in diving behaviour occur; (2) is variation in diving behaviour caused by variation in the energetic cost of diving, or vice versa; and (3) what physiological and/or ecological factors might cause variation in the behaviour and energetics of diving?
Materials and methods
Study animals
Study animals were captured and equipped with data loggers (DLs) at the British Antarctic Survey (BAS) base on Bird Island, South Georgia. All birds used in the experiments were breeding adults from the macaroni penguin colony at Fairy Point on the north side of Bird Island. Although the United Kingdom Animal (Scientific Procedures) Act 1986 does not apply to South Georgia, where this work was conducted, we followed its provisions, especially those set out by the Home Office in the Official Guidance on the operation of the Act. As our benchmark, we followed guidance to researchers using similar methods in the United Kingdom. Our procedures also conformed to the SCAR Code of Conduct for Use of Animals for Scientific Purposes in Antarctica.
Deployment of DLs took place during the austral summers of 2001/02 and 2002/03, and retrieval of DLs took place during the austral summers of 2002/03 and 2003/04. Study birds were identified and captured using the procedures described by Green et al. (2004), and DLs were surgically implanted into the abdominal cavity using previously established techniques (Stephenson et al., 1986; Green et al., 2003). Long-term implantation of these DLs has previously been shown to have no detectable adverse effects on the behaviour, breeding success and survival of this species (Green et al., 2004). In 2001/02, DLs were deployed during the chick rearing phase in January and February (N=39). In 2002/03, DLs were implanted during the incubation phase in November (N=19), chick rearing period in January and February (N=12) and moult phase in March (N=12). DLs were retrieved in the breeding season following implantation. In the 2002/03 breeding season, 34 of 39 penguins with DLs returned after the winter. In the 2003/04 breeding season, 43 of 43 penguins with DLs returned after the winter. In both years, return rates were not significantly different to control groups or previous data for this colony (Green et al., 2004). In both seasons, DLs that had failed during the winter migration were removed during the courtship phase, while DLs which had not failed during the winter were removed during the following moult phase. DLs were removed using the same procedures as during implantation and, after recovery, the penguins resumed their normal activities.
Data loggers
The DLs used in the present study were designed by one of the authors (A.J.W.) and were the same instruments (Mk 3a, Mk 3b) used to study year-round diving behaviour of macaroni penguins (Green et al., in press a). Mk 3a instruments were used for all deployments during the 2001/02 season. Mk 3a instruments were also used for 13 of the deployments in the 2002/03 season, with Mk 3b used for the remainder. Both instruments had dimensions of 362811·mm and weighed 18·g before and 21·g after encapsulation in paraffin wax to provide waterproofing and a silicone coating for biocompatibility. Both instruments could record hydrostatic pressure for conversion to dive depth, heart rate (fH), body attitude (upright or prone) and abdominal temperature (Tab).
Mk 3a DLs had a 32·Mb memory capacity and were programmed to record dive depth every 2·s, fH and body attitude every 10·s and Tabevery 30·s for 453·days. Mk 3b DLs
had a larger memory capacity (64·Mb) and were programmed to record dive depth every 1·s, fHand body attitude every 10·s and Tab every 15·s for 542·days. Mk 3a DLs had a depth
resolution of approximately 0.3·m, while a technical problem with the Mk 3b DLs meant that, although they had a depth resolution of approximately 0.09·m, they failed to record further depth changes deeper than approximately 25·m (Green et al., in press a). All DLs were individually calibrated and had a temperature resolution of approximately 0.06°C. The time that each logger was started, implanted, removed and stopped was carefully noted as GMT. After retrieval, data from all DLs were downloaded onto a computer before being transferred to a UNIX workstation or PC for further analysis.
Dive analysis
While evaluating dive records, dives with maximum depths of <2.4·m were ignored during analyses since, between the surface and this depth, wave action, recorder noise and the interaction between temperature and pressure degraded depth accuracy making it impossible to accurately characterise dives this shallow. No distinction was made between foraging and non-foraging (travelling or searching) dives, and hence all dives with maximum depth of 2.4·m were used in all analyses. In total, 1·616·403 dives were analysed and, for each dive, maximum depth, dive duration and subsequent surface duration were extracted.
Data analysis
attempt), (6) winter. In the current project, only data from the pre-moult trip (N=54 birds) and winter (N=46 birds) phases were used for analysis and are referred to as summer and winter, respectively. The data were further filtered to reduce variability. The duration of the pre-moult trip varied slightly between individuals, so only the first 16·days of the pre-moult trip were used, as this was the minimum duration of this phase in all birds. For winter, the first 13·days of winter were removed as, until this time, some birds still returned to the colony regularly and had not engaged in their full migration, despite the completion of moult. The duration of the winter phase was limited to 190·days, as this was the minimum duration of this phase. For analyses involving dive depth, only data from the 2001/02 season were used (N=31 for summer, N=26 for winter).
The body mass (Mb) of both male and female macaroni
penguins fluctuates during the breeding season, by a factor of up to two (Croxall, 1984). Mb at the end of winter is
substantially greater than at the beginning (Croxall, 1984) but, other than this, Mbis unknown during the winter period. To
understand the relative changes in metabolism, it is beneficial to calculate the rate of oxygen consumption (VO∑),
independently of the potentially confounding effects of fluctuating Mb and body size. In recent years, there has been
much attention on the correct way to account for the effects of Mb in physiological investigations (e.g. Packard and
Boardman, 1999). Unusually, in macaroni penguins, the mass exponent of VO∑ is one, whereas there is no relationship
between heart rate (fH) and Mb(Green et al., 2001; J. A. Green,
C. R. White and P. J. Butler, unpublished data). We found that using mass-specific VO∑(sVO∑) accounts for all of the variation
between individuals in VO∑ at a given fH in this species and
allows the construction of predictive relationships between fH
and sVO∑(Green et al., 2001). Therefore, in the current study,
the use of mass-independent sVO∑ not only accounts for
variation in body size and mass between individuals but also allows us to account for the confounding effects of variable body mass within individuals at different times of year. sVO∑
(in ml·min–1·kg–1) was estimated from fH using the equations of Green et al. (2005), and the standard error of the estimate (S.E.E.) was calculated following the method of Green et al. (2001).
In an effort to evaluate potential changes in minimum or maintenance metabolic rate (MMR), a running average of sVO∑
was computed for each 12-min period throughout each day. The period with the minimum value was assigned as the basal rate of oxygen consumption for that day (sVO∑min). Twelve
minutes was selected as the interval to evaluate sVO∑min, as it
was the inflection point in a plot of running average size against minimum fH (Withers, 2001). In an effort to correct for the possible effects of anapyrexia (a regulated decrease in body temperature) on MMR, sVO∑min was normalised to a
temperature of 39°C (sVO∑minC). sVO∑minC was calculated
according to van’t Hoff principles, assuming an apparent Q10
of 3 (Heldmaier and Ruf, 1992).
Mean abdominal temperature was calculated for each day.
To gain a better understanding of the many physiological processes leading to changes in body temperature, a running average of Tab was calculated every 12·min for each day for
each penguin. The minimum and maximum value for each day for each animal were then extracted (Tab,min and Tab,max,
respectively). Tab,maxwas used to represent the normal, core
body temperature as it was independent of the effects of circulatory changes, regional hypothermia or metabolic suppression associated with diving (Ponganis et al., 2003; Butler, 2004). These factors can conspire together to reduce Tab, and the magnitude and duration of this decrease are
dependent on the duration of diving bouts (Green et al., 2003). sVO∑ while submerged cannot be measured during dives
(Costa, 1988), but sVO∑DC, the sVO∑ of a complete dive cycle
(dive plus the subsequent surface period), can be estimated from mean fH of that dive cycle (Fedak, 1986; Bevan and Butler, 1992; Butler, 1993). Calculated aerobic dive limit (cADL) is usually calculated by dividing useable oxygen stores by an estimate of VO∑ while submerged (Butler and Jones,
1997). Green et al. (2003) used sVO∑DC as a measure and
indicator of VO∑ while submerged and showed that cADL is
not necessarily a fixed quantity. In macaroni penguins, cADL increased with increasing dive duration, because sVO∑DC
decreased with increasing dive duration. As a result of this, in the present study, cADL was calculated, at each observed dive duration, and cADL described in the text is the threshold at which dive duration exceeded the cADL calculated at that dive duration. cADL was calculated using oxygen stores of 58·ml·kg–1, as in a previous study (Green et al., 2003). 95% confidence intervals of cADL were calculated by repeating this process but substituting sVO∑DCwith the upper and lower 95%
confidence intervals of sVO∑DCat each dive duration.
Estimates of sVO∑ are quoted in the text ± S.E.E., and
comparisons of these data were made using Woolf’s test for differences, which is most appropriate for the analysis of data derived from a predictive relationship (R. L. Holder, personal communication). All other data were analysed using general linear model (GLM), and means are quoted as ±S.E.M. Results were considered significant at P<0.05, and the significance of statistical tests is quoted in the text.
Results
Diving behaviour
considerably greater variability, maximum dive depth showed a similar pattern of change (Fig.·2A). Mean dive depth also showed a similar pattern of change but was up to 50% greater during the middle of winter than during summer. During late-winter, mean dive depth appeared to be closer to mean dive depth during summer (Fig.·2B).
To investigate these differences in more detail, mean and maximum dive duration and depth were derived for each bird for the summer (276·641 dives) and for the middle of the two stable periods during mid- and late-winter (phasedays 69–85, 121·681 dives; phasedays 149–165, 113·174 dives, respectively). These periods are illustrated in Fig.·1. There was no difference between the sexes in mean and maximum dive duration and depth. Mean dive duration was significantly different between each of these periods (GLM with Tukey post-hoc tests; F2,113=95.1, P<0.0001; Table·1). Mean
maximum dive duration was also significantly different between each of these periods (GLM with Tukey post-hoc tests; F2,113=154.7, P<0.0001;
Table·1). Mean dive depth was significantly greater during mid-winter than during summer and late-winter (GLM with Tukey post-hoc tests; F2,113=12.5, P<0.0001; Table·1). Mean
maximum dive depth was not significantly different between mid-and late-winter, but both were significantly greater than mean maximum dive depth during summer (GLM with Tukey post-hoc tests; F2,113=13.8, P<0.0001; Table·1). Abdominal temperature and metabolic
rate
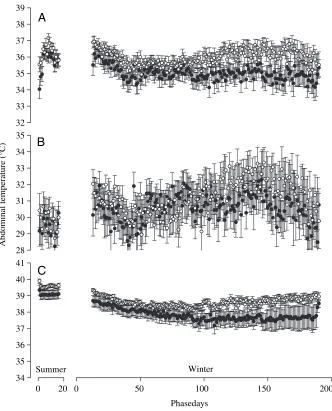
There was some variation between individuals in the pattern of change of daily abdominal temperature (Tab,D),
but Fig.·3A shows the mean for all animals. In female penguins, Tab,D
decreased by approximately 1°C during early winter from the summer level then remained at this level for the rest of the winter. In male penguins, Tab,Ddeclined by approximately 0.8°C
at the beginning of winter from the mean summer level but later increased 0 20
Phaseday
0 50 100 150 200
60 80 100 120 140 160 180
Summer Winter
140 160 180 200 220 240 260
A
B
Dive duration (s)
Fig.·1. (A) Mean daily maximum dive duration (±S.E.M.) and (B) mean daily dive duration (± S.E.M.) recorded from female (filled symbols) and male (open symbols) macaroni penguins during the summer/pre-moult and winter/migration phases of their annual cycle. The solid black bars represent the three sampling periods used in the analysis of changes in dive duration and other variables.
0 20
Dive depth (m)
Phaseday
0 50 100 150 200
Summer Winter
0 15 30 45 60 75 60 75 90 105 120 135
A
B
Fig.·2. (A) Mean daily maximum dive depth (±S.E.M.) and (B) mean daily dive depth (±S.E.M.)
to approximately the same as during summer. Changes in minimum abdominal temperature (Tab,min) also varied between
individuals but a general pattern was identified (Fig.·3B). In both male and female penguins, Tab,min was variable
during winter but was up to 2°C higher than during summer. Significant changes in maximum abdominal temperature (Tab,max) were far more consistent
between individuals. Tab,max declined
steadily over the first 100·days of winter to a level around 1.5°C lower than that during summer. Tab,maxthen remained at
this level for the rest of the winter in female penguins but increased slightly, while remaining below summer levels, in male penguins (Fig.·3C). GLM was used to further investigate the changes in temperature (Tab,D, Tab,min or Tab,max) for
summer, mid-winter and late-winter as described above. In each analysis, temperature (Tab,D, Tab,minor Tab,max) was
the dependent variable, with sex and season as factors (Table·2). Mean Tab,D
was significantly greater in males than females but not significantly different between the seasons. Mean Tab,min was
not significantly different between the seasons or sexes. Mean Tab,max was
significantly greater in males than females and was significantly lower during mid-winter and late-winter than during the summer.
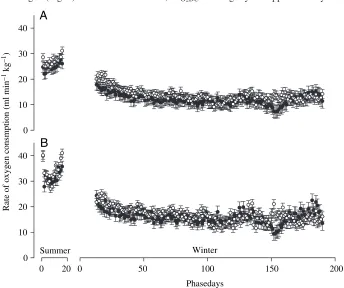
Minimum sVO∑ showed an inverse
pattern to that of diving behaviour (Fig.·4A). sVO∑min was lower during
winter than summer. During winter, sVO∑min decreased initially until
approximately day 50 and then remained
relatively constant at around 50% of the summer value until the end of winter. Normalising sVO∑minfor the effects of body
temperature had little effect on this pattern (Fig.·4B). Mean sVO∑minduring summer was significantly greater than sVO∑min
during mid-winter (days 69–85) whether normalised or not (Woolf’s test for differences; Table·3). The magnitude of the
decrease in sVO∑minfrom summer to winter only changed from
51% to 49% with normalisation.
Energetic cost of diving
[image:5.612.49.569.88.161.2]Changes in maximum dive duration and depth as winter progressed were mirrored by changes in the energetic cost of Table 1. Mean and mean maximum (±S.E.M.) dive duration and dive depth at different times of year in macaroni penguins
Duration (s) Depth (m)
Period Mean Maximum N Mean Maximum N
Summer 97.3±1.4λ,γ 161.8±1.7λ,γ 54 31.8±1.4λ 78.8±2.8λ,γ 31
Mid-winter 134.0±2.3ϕ,λ 214.0±2.5ϕ,λ 38 45.0±2.9λ,γ 104.5±4.5λ 22
Late-winter 114.7±3.0ϕ,γ 200.4±3.5ϕ,γ 24 33.7±1.9γ 93.3±3.9γ 17
Significant differences between pairs within each column are represented by the following symbols λ,γ,ϕ.
32 33 34 35 36 37 38 39
Phasedays
0 50 100 150 200
0 20 34
35 36 37 38 39 40 41
Abdominal temperature (°C)
28 29 30 31 32 33 34 35
C
A
B
Summer Winter
Fig.·3. (A) Mean daily abdominal temperature (± S.E.M.), (B) mean daily minimum
abdominal temperature (±S.E.M.) and (C) mean daily maximum abdominal temperature (±
S.E.M.) recorded from female (filled symbols) and male (open symbols) macaroni penguins
[image:5.612.232.564.189.600.2]diving. For any given dive duration, mean sVO∑DCdecreased as
winter progressed, whereas during summer, sVO∑DC was
unchanged (Fig.·5). At the start of winter, sVO∑DCwas slightly
less than that during the summer and decreased over the first part of the winter migration before reaching a stable level after approximately 50·days. This stable level was then maintained for most of the winter before increasing slightly at the end. As with mean dive duration, mean sVO∑DCwas
derived for each bird for the summer and for mid- and late-winter (phasedays 69–85 and phasedays 149–165, respectively) at three ranges of dive duration (41–50·s, 91–100·s, 141–150·s). With the exception of 141–150·s dives in females, sVO∑DC
was significantly different among these periods at each range of dive duration in both sexes (Woolf’s test for differences; Table·4). Post-hoc Z-tests with Bonferroni corrections showed that, where there was a significant difference, sVO∑DC was the same
during both winter periods but both of these were significantly lower than sVO∑DCduring summer.
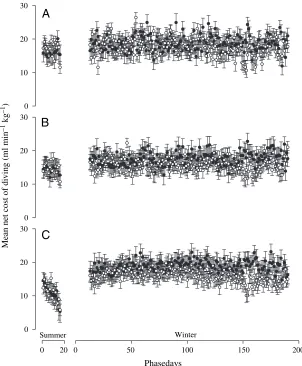
The net cost of diving above MMR or maintenance metabolism (sVO∑net)
was estimated by subtracting mean sVO∑minfrom mean sVO∑DCfor each day
(Fig.·6). sVO∑net was derived for each
[image:6.612.42.560.99.167.2]bird for the summer and for mid- and late-winter (phasedays 69–85 and phasedays 149–165, respectively) at Table 2. Mean daily abdominal temperature (Tab,D), minimum abdominal temperature (Tab,min) and maximum abdominal
temperature (Tab,max) in macaroni penguins at three different seasons of their annual cycle
Female Male
Season N Mean Tab,D Mean Tab,min Mean Tab,max N Mean Tab,D Mean Tab,min Mean Tab,max
Summer 27 35.8±0.33 29.4±0.68 39.1±0.27λ,γ 25 36.3±0.29 29.9±0.64 39.6±0.11λ,γ
Mid-Winter 19 35.0±0.47 31.0±0.84 37.7±0.37γ 18 35.6±0.36 30.6±0.74 38.4±0.13γ Late-Winter 12 34.7±0.53 30.2±0.90 37.6±0.58λ 10 36.2±0.49 32.0±1.09 38.7±0.21λ
GLM was used to investigate the change in temperature. Tab,D and Tab,maxwere significantly greater in male penguins (P=0.027 and 0.005,
respectively). There was significant effect of season on Tab,maxonly (P<0.0001). Differences between pairs of temperatures as indicated by post-hocTukey tests are represented by the following symbols λ,γ.
Phasedays
0 50 100 150 200
0 20 0
10 20 30 40
Rate of oxygen consmption (ml min
–1
kg
–1
)
0 10 20 30 40
B
A
Summer Winter
Fig.·4. (A) Mean daily minimum rate of oxygen consumption (± S.E.E.) and (B) mean daily minimum rate of oxygen consumption adjusted for maximum daily body temperature (±S.E.E.)
recorded from female (filled symbols) and male (open symbols) macaroni penguins during the summer/pre-moult and winter/migration phases of their annual cycle.
Table 3. Mean minimum rate of oxygen consumption (sVO∑min) and mean minimum rate of oxygen consumption adjusted for body temperature (sVO∑minC) during summer and mid-winter in male and female macaroni penguins
Sex Season N sVO∑min Z P sVO∑minC Z P
Female Summer 28 24.9±2.0 32.0±2.1
Female Winter 19 11.6±2.2 4.39 <0.0001 15.7±2.2 5.37 <0.0001
Male Summer 25 27.2±2.1 33.3±2.1
Male Winter 19 13.7±2.2 4.45 <0.0001 17.5±2.2 5.17 <0.0001
Z-tests (with Bonferroni corrections for multiple tests) were used to compare estimates of sVO∑minor sVO∑minCbetween summer and
[image:6.612.43.387.260.548.2] [image:6.612.39.561.643.710.2]three ranges of dive duration (41–50·s, 91–100·s, 141–150·s). With the exception of 141–150·s dives in females, sVO∑net was not significantly different
among these periods at each range of dive duration in both sexes (Woolf’s test for differences; Table·4). Post-hoc Z -tests with Bonferroni corrections showed that in dives of 141–150·s duration in females, sVO∑net was the same during
both winter periods but both of these were significantly higher than sVO∑DC
during summer.
If sVO∑DC at a given dive duration
decreases, then if oxygen stores remain unchanged, cADL should increase. cADL and the proportion of dives exceeding cADL were calculated for the three sampling periods used previously (Fig.·7). cADL is derived from several estimated quantities. It is therefore difficult to make statistical comparisons, other than to examine the extent of overlap in the ranges of 95% confidence intervals. Fig.·7 indicates that in both males and female penguins, cADL was up to 45% greater during the mid- and late-winter than during summer. In female penguins, the proportion of dives within the cADL was the same during all three phases whereas in male penguins the proportion of dives within the cADL was the same during summer and mid-winter but was greater during late-mid-winter.
Discussion
Comparison with earlier work In our earlier study of the diving physiology of macaroni penguins
(Green et al., 2003), our measurements of fH were approximately 30% lower than those in the present study. After conversion, our estimates of sVO∑DCwere approximately 45%
lower in the previous study. We therefore derived an estimate of cADL during the summer of the present study that was 36% less than that in the previous study and which, because of the shape of the distribution of dive durations, resulted in 58% fewer dives being within the cADL. Some of the difference in sVO∑DC may be due to natural variability in metabolic rate
and/or the demands of diving in different seasons, but re-examination of both data sets has unearthed other differences. We conclude that rates of oxygen consumption are higher in the present study because we have improved our methods of filtering out periods when the heart rate measurements are unreliable. The percentage of dives within the cADL in the present study now matches that found in other studies of diving
birds (Butler, 2001) and suggests that VO∑while submerged is
substantially less than sVO∑DC. While cADL remains a useful
comparative measure, both within and between species, this result once again highlights the importance of obtaining improved estimates of both usable oxygen stores and VO∑while
submerged if cADL is to truly reflect the duration of dives at which usable oxygen stores are exhausted (Green et al., in press b).
Changes in diving behaviour
In both sexes, diving behaviour during the pre-moult trip was relatively consistent, with little change in mean and maximum dive duration and depth. During the winter migration, however, the picture was rather different and two distinct stable phases can be identified. At the start of winter, both mean and maximum dive durations and depth were 20
30 40 50 60
Phasedays
0 50 100 150 200
0 20 20
30 40 50 60
Mean rate of oxygen consumption of dive cycle (ml min
–1
kg
–1)
20 30 40 50 60
C
A
B
Summer Winter
greater than those during the summer and proceeded to increase further until around day 50. These higher levels were maintained until around day 100. During this mid-winter period, mean and maximum depth and duration were significantly greater than those during the summer. There then followed a period of change where mean depth, mean duration and maximum duration decreased to other lower stable levels. During this late-winter stable period, maximum depth was unchanged but mean depth and mean and maximum duration were significantly lower. Indeed, mean depth was equal to that during summer.
[image:8.612.44.560.100.329.2]The driving force behind these changes in behaviour is unclear but is likely to depend on the distribution of prey species with respect to depth. Antarctic krill (Euphausia superba) is the dominant species in the zooplankton assemblage of the Southern Ocean (Everson, 2000). Macaroni penguins feed predominantly on Antarctic krill, at least in the summer (Croxall et al., 1997), but in decreasing amounts in recent years (Barlow et al., 2002). Antarctic krill vary Table 4. Results from Woolf’s test for differences between (1) estimates of VO∑of completed dive cycles (sVO∑DC±S.E.E.) and (2)
the net cost of diving (sVO∑net±S.E.E.) at three different periods of the annual cycle
Dive duration sVO∑DC(ml· sVO∑net(ml·
Sex range (s) Season O2 ·min–1·kg–1) χ2 P O2·min–1·kg–1) χ2 P
Female 41–50 Summer 40.6±2.1λ,γ 15.8±2.0
Mid-winter 29.2±2.2λ 17.6±2.2
Late-winter 28.9±2.4γ 18.54 <0.0001 19.2±2.4 1.19 0.5518
91–100 Summer 39.0±2.1λ,γ 14.1±2.0
Mid-winter 27.7±2.2λ 16.1±2.2
Late-winter 29.6±2.5γ 15.55 0.0004 19.8±2.4 3.30 0.1920
141–150 Summer 35.7±2.1 10.9±2.0λ,γ
Mid-winter 29.6±2.2 18.0±2.2λ
Late-winter 30.9±2.5 4.54 0.103 21.1±2.4γ 11.60 0.0030
Male 41–50 Summer 44.2±1.5λ,γ 16.9±2.0
Mid-winter 32.2±1.5λ 18.5±2.2
Late-winter 28.3±1.7γ 55.05 <0.0001 15.2±3.2 0.77 0.6807
91–100 Summer 42.4±1.5λ,γ 15.1±2.0
Mid-winter 29.4±1.5λ 15.7±2.2
Late-winter 27.0±1.7γ 57.31 <0.0001 13.9±3.2 0.21 0.9006
141–150 Summer 38.7±1.4λ,γ 11.5±2.0
Mid-winter 30.8±1.5λ 17.0±2.2
Late-winter 26.5±1.7γ 32.12 <0.0001 13.4±3.2 3.39 0.1838
Tests were repeated at three ranges of dive duration in male and female macaroni penguins. Significant differences between pairs determined by Z-tests with Bonferroni corrections within each dive range are represented by the following symbols λ,γ,ϕ.
0 10 20 30
Phasedays
0 50 100 150 200
0 20 0
10 20 30 0 10 20 30
C
A
B
Summer Winter
Mean net cost of diving (ml min
–1
kg
–1)
Fig.·6. Mean net cost of diving (±S.E.E.) for dives of
[image:8.612.44.346.375.741.2]annually, seasonally, diurnally and geographically in their location in the water column (Godlewska, 1996) but usually undertake diurnal vertical migration; spending the night close to the surface and the day at deeper depths. Despite this, during the day, krill will nearly always be located within the diving depth range of macaroni penguins. The mean depth of the krill varies substantially between locations in the Atlantic sector of the Southern Ocean, Scotia Sea and waters around South Georgia (Everson, 1984; Godlewska, 1996). The mean depth in a review of studies varied from 28 to 156·m and the amplitude of migration from 2.5 to 59·m, depending on the timing and location of the studies. Indeed, some data suggest that close to South Georgia, pressure from predatory fish feeding on the shelf bottom causes diurnal vertical migration to be reversed, with krill found relatively close to the surface (around 50·m) during the day and dispersed throughout the water column at night (Everson, 1984).
There is little information on the foraging location of macaroni penguins, especially during winter. During the chick rearing season, the penguins undertake short foraging trips and remain close to South Georgia over the continental shelf (Barlow and Croxall, 2002). During the longer incubation foraging trip, the penguins range further afield to far deeper waters in the Polar Frontal Zone (Barlow and Croxall, 2002). The incubation foraging trip is of a similar duration and for a similar purpose (rapidly to replenish or increase body reserves) to the pre-moult foraging trip, so it seems reasonable to assume for the time being that the penguins forage in a similar location. The location of macaroni penguins during the winter is currently unknown. However, it seems likely that during the winter they migrate to the open ocean away from the vicinity of South Georgia. A difference in the depth of krill swarms in the open ocean is therefore a likely explanation for the
difference in mean diving depth between summer and winter. Figs·1 and 2 suggest that there was a further change in the mean depth of the krill from mid- to late-winter, due either to a difference in the location of the penguins or to the behaviour of the krill. In late-winter, mean depth, mean duration and maximum duration declined while maximum depth did not change. The decrease in maximum duration was only slight (6.5%; Table·1) and there were no changes at this time in the physiological parameters measured (Figs·3–6), implying that the diving capacity of the penguins did not change. As a result, they were able to dive more comfortably within their limits. This is reflected in an increase in the proportion of dives within the cADL between mid-winter and late-winter (significantly so in male penguins), despite no significant change in the cADL (Fig.·7).
Energetic cost of diving
The increases in mean and maximum dive depth and duration from summer to winter, and the continued progressive increases through the first part of winter, were inversely matched precisely by changes in the energetic cost of diving. As mean and maximum dive duration increased, mean sVO∑DC
for a given dive duration decreased (Fig.·5). It appears therefore that increased dive duration and depth were facilitated by a decrease in the energetic cost of diving. A progressive improvement in diving ability due to a reduction in energetic costs has not previously been observed in a mature diving animal. Juvenile diving animals commonly show an improvement in their ability to dive during development (Burns, 1999; Ponganis et al., 1999; Noren et al., 2001). Other studies have shown that oxygen stores can vary seasonally in adult animals (MacArthur, 1990) but that any apparent advantage of this is cancelled out by an increase in VO∑while
diving (MacArthur et al., 2000). Increased oxygen availability during diving was induced in tufted ducks (Aythya fuligula) by training them to dive for longer (Stephenson et al., 1989). However, a similar increase induced by training in muskrats (Ondatra zibethicus) was again cancelled out by an increase in VO∑ during diving (MacArthur et al., 2003). In the current
study, we do not know how, or even whether, oxygen stores varied. However, the proportion of dives in excess of the cADL did not vary between summer and mid-winter (Fig.·7), suggesting that whether or not the estimate of oxygen stores we used is accurate, it did not vary. The penguins must have been under pressure to dive deeper and for longer during winter, and apparently this increase was accommodated solely by the decreased energetic cost of diving. This decrease must be achievable only in the time scale and under the conditions experienced in winter, or else the penguins would modify their energetic costs in the same way during the summer and we would see more dives within the cADL at this time.
Although seasonal variation in sVO∑ while diving has not
been demonstrated previously, some authors have speculated on its existence (Bennett et al., 2001). In the current study, we show that, in macaroni penguins, variation in sVO∑while diving
is due almost entirely to variation in sVO∑min. It is not possible
A
20 40 60 80 100 120 140 160
B
Phase
Summer Mid-winter Late-winter % of dives within calculated aerobic
dive limit
Calculated aerobic
dive limit (s)
0 20 40 60 80
to measure basal metabolic rate (BMR) in an active animal, but sVO∑min provides an approximation of the maintenance
requirements of the penguins while at sea, excluding diving behaviour. sVO∑minshowed a pattern of change which matched
that of sVO∑DC(Fig.·4) and there are good linear relationships
between mean daily sVO∑minand sVO∑DC(r2=0.73 and 0.83 for
dives of 100·s duration in females and males, respectively). It seems clear that the decrease in sVO∑DCand increase in diving
capacity are facilitated by this drop in minimum or basal metabolic rate.
Seasonal change in basal metabolic rate
Seasonal variation in BMR has been demonstrated in many species. Among diving animals, captive female grey seals (Boily, 1996; Boily and Lavigne, 1997), harbour seals (Phoca vitulina) and harp seals (Phoca groenlandica; Renouf and Gales, 1994) show a marked underlying seasonal variability in resting metabolic rate, although the pattern of this variability varies between species.
The maintenance of a high core body temperature in endotherms carries a high metabolic cost (Bennett and Ruben, 1979). In water, which has a thermal conductance 25greater than air, these metabolic costs are likely to be even higher, even for an animal as well adapted to an aquatic lifestyle as a penguin. In several species of penguins, resting VO∑ is
approximately twice as high when they are in relatively cold water than when they are in air (Stahel and Nicol, 1982; Culik et al., 1991; Bevan et al., 1995). Other studies have shown that penguins have no thermoneutral zone in water, and metabolic rate increases with decreasing water temperature (Stahel and Nicol, 1982; Barré and Roussel, 1986). In little penguins (Eudyptula minor), this trend occurs until a critical temperature, beyond which metabolic rate increases sharply (Stahel and Nicol, 1982).
Resting sVO∑during summer of macaroni penguins in water
at 6.8°C, recorded using respirometry, was 27.0 and 24.5·ml·O2·min–1·kg–1 for females and males, respectively
(Green et al., 2005). These values are similar to sVO∑minduring
summer in the present study. However, by mid-winter, sVO∑min
was closer to values obtained from the same penguins while resting in air (10.7 and 9.7·ml·O2·min–1·kg–1for females and
males, respectively; Green et al., 2005). In fasted king penguins (Aptenodytes patagonicus), resting VO∑in water was
approximately 2that in air. However, in well-fed penguins, resting VO∑in water was substantially lower and was the same
as resting VO∑in air (Fahlman et al., in press). It is suggested
by these authors that a complex interaction between nutritional state, vasoconstriction, fat deposition and fat mobilisation causes a decrease in thermal conductance (the inverse of insulation) of around 25% and therefore a substantial reduction in metabolic rate.
Thermoregulation
Perhaps macaroni penguins are also able to reduce the cost of thermoregulation during the winter. Thermoregulatory costs can be reduced by either decreasing the temperature gradient
(by reducing body temperature or increasing external temperature) or by decreasing the rate of heat transfer or thermal conductance. Temporarily decreasing core temperature (anapyrexia) while inactive on a daily or seasonal basis is used by some endotherms to conserve energy when the temperature is very low and/or food is scarce (Nedergard and Cannon, 1990). Even in a period of high activity, barnacle geese (Branta leucopsis) were found to save considerable amounts of energy for their spring migration through having a reduced Tab (Butler and Woakes, 2001). However, in the
present study, mean Tab,Ddid not change from the summer to
winter (Table·2) and the modest decline in Tab,max (a proxy for
body temperature) of approximately 1.5°C (Table·2) was not nearly sufficient to explain the decrease in sVO∑min (Fig.·4).
Perhaps more likely is that, by migrating northwards, macaroni penguins are able to forage in warmer waters during winter than they are during the summer. Factors controlling the sea surface temperature (SST) are beyond the scope of this article but, as an example, at the longitude of Bird Island (approx. 38° W) during July, SST increases by approximately 0.92°C for each degree of latitude travelled north from 53° S (the latitude of Bird Island) to 38° S and beyond.
before the moult when they will be worn and undoubtedly less effective.
Interactions between Tab, water temperature and insulation
can be summarised in a simple model of thermal conductance changes. Conductance (Cin W·deg.–1·m–2) can be calculated simply as C=MO∑(Tb–Ta)–1A–1, where Tband Taare core body
and ambient body temperature, respectively (Tab and SST in
this case), and A is the surface area of the bird in m2. MO∑
(metabolic rate) is calculated from sVO∑min using an
equivalence of 18.889·J·ml–1·O2 (Green et al., 2002). Surface
area is given by Meeh’s formula, with a constant of 0.77 (Barré and Roussel, 1986). While there is much debate on the correct constant for use in penguins (Luna-Jorquera et al., 1997), the value has no effect in our model, which looks at relative effects of insulation and Tain isomorphic individuals. Exact mass is
unknown but it is assumed to be the same for summer and winter at 4·kg. Varying mass has little effect on the conclusions of the model. We do not know how temperature changes with depth where the penguins are located, so Tais assumed to be
equal to SST. During the pre-moult trip, if we assume that the penguins forage close to the Maurice Ewing Bank (longitude –41.5°, latitude –51.5°) then SST during March 2002 was 5.8°C. Conductance can therefore be calculated for the pre-moult trip. If we assume that C is equal in the winter or decreases by 10, 20, 30 or 40% (i.e. insulation increases) then we can calculate what percentage of the decrease that we observe in sVO∑min might be explained by increasing water
temperatures, i.e. reducing ∆T, the temperature difference between Taand Tb(Fig.·8).
Modelling thermal conductance in this way assumes that animals are passive bodies, and studies have shown that such
an assumption can oversimplify the complexity of changes in conductance in animals (Hind and Gurney, 1997). Furthermore, heat generated during locomotion may be used in thermoregulation, while movement itself will alter conductance (De Vries and Van Eerden, 1995; Hind and Gurney, 1997). However, penguins do not dive to forage at night (Wilson et al., 1993; Green et al., in press a) and therefore at this time probably remain inactive at the water surface. Indeed, in the present study, sVO∑min was nearly always
recorded during the hours of darkness. Therefore, while changes in conductance are unlikely to provide the full explanation for the decrease in MMR during winter, Fig.·8 suggests that this will be an important component. This simple model shows us that small increases in insulation, particularly internal insulation, and/or water temperature can have a large effect on the reduction of metabolic rate. The progressive decrease in sVO∑min at the start of winter and the increase
towards the end of winter could be explained by the penguins moving from cooler to warmer waters and vice versa or a progressive improvement in body condition and insulation. In great cormorants (Phalacrocorax carbo carbo), water temperature, body temperature and body fat thickness were found to be major contributors to diving energetics (Grémillet et al., 1998). Further work on the winter location, body condition and mass of penguins will assist us in assessing the relative contribution of the effect of a change in thermal conductance and/or ∆Tin macaroni penguins.
List of symbols
fH heart rate
C conductance
cADL calculated aerobic dive limit
Mb body mass
MMR minimum metabolic rate
MO∑ metabolic rate
sVO∑ mass-specific rate of oxygen consumption
sVO∑DC mass-specific rate of oxygen consumption during
dive cycles
sVO∑min minimum mass-specific rate of oxygen
consumption
sVO∑minC minimum mass-specific rate of oxygen
consumption normalised to 39°C sVO∑net net cost of diving
Ta ambient temperature
Tab abdominal temperature
Tab,D mean daily abdominal temperature Tab,max daily maximum abdominal temperature Tab,min daily minimum abdominal temperature
Tb body temperature
VO∑ rate of oxygen consumption
The authors would like to thank Nick Warren, who assisted in the retrieval of the data loggers, and the rest of the science team at Bird Island, especially Jane Tanton. The authors would also like to thank Dr Craig White for helpful Change in water temperature (°C)
0 2 4 6 8 10 12 14 16
% of decrease explained
0 20 40 60 80 100
Ta–Tb
16 18 20 22 24 26 28 30
+40% +30%+20% +10% 0%
Fig.·8. A simple model indicating to what extent a change in water temperature, or ∆T [ambient temperature (Ta) – body temperature
(Tb)], could account for the observed decrease in minimum
mass-specific rate of oxygen consumption (sVO∑min) from summer to winter
discussions and comments on the manuscript. We are grateful to the British Atmospheric Data Centre, which provided us with access to the Met Office Sea Surface Temperature Data. This work was funded by NERC, under their Antarctic Funding Initiative (AFI), with logistical support provided by the British Antarctic Survey.
References
Barlow, K. E. and Croxall, J. P.(2002). Seasonal and interannual variation in foraging range and habitat of macaroni penguins Eudyptes chrysolophus at South Georgia. Mar. Ecol. Prog. Ser. 232, 291-304.
Barlow, K. E., Boyd, I. L., Croxall, J. P., Reid, K., Staniland, I. and Brierley, A. S.(2002). Are penguins and seals in competition for Antarctic krill at South Georgia? Mar. Biol. 140, 205-213.
Barré, H.(1984). Metabolic and insulative changes in winter- and summer-acclimatized king penguins chicks. J. Comp. Physiol. 154, 317-324. Barré, H. and Roussel, B.(1986). Thermal and metabolic adaptation to first
cold-water immersion in juvenile penguins. Am. J. Physiol. 251, R456-R462.
Beck, C. A., Bowen, W. D. and Iverson, S. J.(2000). Seasonal changes in buoyancy and diving behaviour of adult grey seals. J. Exp. Biol. 203, 2323-2330.
Bennett, A. F. and Ruben, J. A. (1979). Endothermy and activity in vertebrates. Science206, 649-654.
Bennett, K. A., McConnell, B. J. and Fedak, M. A.(2001). Diurnal and seasonal variations in the duration and depth of the longest dives in Southern elephant seals (Mirounga leonina): possible physiological and behavioural constraints. J. Exp. Biol. 204, 649-662.
Bevan, R. M. and Butler, P. J.(1992). Cardiac output and blood flow distribution during swimming and voluntary diving of the tufted duck (Athya Fuligula). J. Exp. Biol. 168, 199-217.
Bevan, R. M., Woakes, A. J., Butler, P. J. and Croxall, J. P.(1995). Heart rate and oxygen consumption of exercising gentoo penguins. Physiol. Zool.
68, 855-877.
Blem, C. R.(1990). Avian energy storage. Curr. Ornithol. 7, 59-113. Boily, P.(1996). Metabolic and hormonal changes during the molt of captive
gray seals (Halichoerus grypus). Am. J. Physiol. 270, R1051-R1058. Boily, P. and Lavigne, D. M.(1997). Developmental and seasonal changes
in resting metabolic rates of captive female grey seals. Can. J. Zool. 75, 1781-1789.
Burns, J. M.(1999). The development of diving behavior in juvenile Weddell seals: pushing physiological limits in order to survive. Can. J. Zool. 77, 737-747.
Butler, P. J.(1993). To what extent can heart rate be used as an indicator of metabolic rate in free-living marine mammals. Symp. Zool. Soc. Lond. 66, 317-332.
Butler, P. J.(2001). Diving Beyond the Limits. News Physiol. Sci. 16, 222-227.
Butler, P. J.(2004). Metabolic regulation in diving birds and mammals. Resp. Physiol. Neurobiol. 141, 297-315.
Butler, P. J. and Jones, D. R.(1997). Physiology of diving of birds and mammals. Physiol. Rev77, 837-899.
Butler, P. J. and Woakes, A. J.(2001). Seasonal hypothermia in a large migrating bird: saving energy for fat deposition? J. Exp. Biol. 204, 1361-1367.
Charrassin, J.-B. and Bost, C.-A.(2001). Utilisation of the oceanic habitat by king penguins over the annual cycle. Mar. Ecol. Prog. Ser. 221, 285-297.
Costa, D. P.(1988). Methods for studying the energetics of freely diving animals. Can. J. Zool. 66, 45-52.
Croxall, J. P.(1984). Seabirds. In Antarctic Ecology(ed. R.M. Laws), pp. 533-619. London: Academic Press.
Croxall, J. P., Prince, P. A. and Reid, K.(1997). Dietary segregation of krill-eating South Georgian seabirds. J. Zool. (Lond.)242, 531-536.
Culik, B. M., Wilson, R. P., Dannfield, R., Adelung, D., Spairani, H. J. and Coria, N. N. C.(1991). Pygoscelid penguins in a swim canal. Polar Biol.
11, 277-282.
Everson, I.(1984). Zooplankton. In Antarctic Ecology(ed. R. M. Laws), pp. 463-490. London: Academic Press.
Everson, I.(2000). Krill: Biology, Ecology and Fisheries. Oxford: Blackwell Science.
Fahlman, A., Butler, P. J., Handrich, Y., Woakes, A. J. and Schmid, A. (in press). Metabolism and thermoregulation during fasting in king penguins, Aptenodytes patagonicus, in air and water. Am. J. Physiol.
Fedak, M. A.(1986). Diving and exercise in seals: a benthic perspective. In Diving In Animals And Man(ed. A. D. Brubakk, J. W. Kanwisher and G. Sundess), pp. 11-32. Trondheim: Tapir.
Godlewska, M. (1996). Vertical migrations of krill (Euphausia superba Dana). Pol. Arch. Hydrobiol. 43, 9-63.
Green, J. A., Butler, P. J., Woakes, A. J., Boyd, I. L. and Holder, R. L. (2001). Heart rate and rate of oxygen consumption of exercising macaroni penguins. J. Exp. Biol. 204, 673-684.
Green, J. A., Butler, P. J., Woakes, A. J. and Boyd, I. L.(2002). Energy requirements of female macaroni penguins breeding at South Georgia. Func. Ecol. 16, 671-681.
Green, J. A., Butler, P. J., Woakes, A. J. and Boyd, I. L.(2003). Energetics of diving in macaroni penguins. J. Exp. Biol. 206, 43-57.
Green, J. A., Tanton, J. L., Woakes, A. J., Boyd, I. L. and Butler, P. J. (2004). Effects of long-term implanted data loggers on macaroni penguins. J. Avian Biol. 35, 370-376.
Green, J. A., Boyd, I. L., Woakes, A. J., Warren, N. L. and Butler, P. J. (in press a). Behavioural flexibility during year-round foraging in macaroni penguins. Mar. Ecol. Prog. Ser.
Green, J. A., Halsey, L. G. and Butler, P. J.(in press b). To what extent is the foraging behaviour of aquatic birds limited by their physiology? Physiol. Biochem. Zool.
Green, J. A., Woakes, A. J., Boyd, I. L. and Butler, P. J. (2005). Cardiovascular adjustments during locomotion in penguins. Can. J. Zool.
83, 445-454.
Grémillet, D., Tuschy, I. and Kierspel, M.(1998). Body temperature and insulation in diving Great Cormorants and European Shags. Func. Ecol. 12, 386-394.
Heldmaier, G. and Ruf, T.(1992). Body Temperature and metabolic rate during natural hypothermia in endotherms. J. Comp. Physiol. 162, 696-706. Hind, A. T. and Gurney, W. S. C.(1997). The metabolic cost of swimming
in marine homeotherms. J. Exp. Biol. 200, 531-542.
Le Maho, Y., Delclitte, P. and Chatonnet, J.(1976). Thermoregulation in fasting emperor penguins under natural conditions. Am. J. Physiol. 231, 913-922.
Luna-Jorquera, G., Wilson, R. P., Culik, B. M., Aguilar, R. and Guerra, C.(1997). Observations on the thermal conductance of Adélie (Pygoscelis adeliae) and Humboldt (Spheniscus humboldti) penguins. Polar Biol. 17, 69-73.
MacArthur, R. A.(1990). Seasonal changes in the oxygen storage capacity and aerobic dive limits of the muskrat (Ondatra zibethicus). J. Comp. Physiol. 160, 593-599.
MacArthur, R. A., Humphries, M. M., Fines, G. A. and Campbell, K. L. (2000). Body oxygen stores, aerobic dive limits, and the diving abilities of juvenile and adult muskrats (Ondatra zibethicus). Physiol. Biochem. Zool.
74, 178-190.
MacArthur, R. A., Weseen, G. L. and Campbell, K. L.(2003). Diving experience and the aerobic dive capacity of muskrats: does training produce a better diver? J. Exp. Biol. 206, 1153-1161.
Nedergard, J. and Cannon, B. (1990). Mammalian hibernation. Philos. Trans. R. Soc. London Ser. B 326, 669-686.
Noren, S. R., Williams, T. D., Pabst, D. A., McLellan, W. A. and Dearolf, J. L.(2001). The development of diving in marine endotherms: preparing the skeletal muscles of dolphins, penguins, and seals for activity during submergence. J. Comp. Physiol. 171, 127-134.
Packard, G. C. and Boardman, T. J.(1999). The use of percentages and size-specific indices to normalize physiological data for variation in body size: wasted time, wasted effort? Comp. Biochem. Physiol. 122, 37-44. Ponganis, P. J., Starke, L. N., Horning, M. and Kooyman, G. L.(1999).
Development of diving capacity in emperor penguins. J. Exp. Biol. 202, 781-786.
Ponganis, P. J., Van Dam, R. P., Levenson, D. H., Knower, T., Ponganis, K. V. and Marshall, G.(2003). Regional heterothermy and conservation of core temperature in emperor penguins diving under sea ice. Comp. Biochem. Physiol. 135, 477-487.
Renouf, D. and Gales, R.(1994). Seasonal variation in the metabolic rate of harp seals: unexpected energetic economy in the cold ocean. Can. J. Zool.
72, 1625-1632.
Factors affecting stroking patterns and body angle in diving Weddell seals under natural conditions. J. Exp. Biol. 206, 1461-1470.
Stahel, C. D. and Nicol, S. C.(1982). Temperature regulation in the little penguin, Eudyptyla minor, in air or water. J. Comp. Physiol. 148, 93-100. Stephenson, R., Butler, P. J. and Woakes, A. J. (1986). Diving behaviour and heart rate in tufted ducks Aythya fuligula. J. Exp. Biol. 126, 341-359.
Stephenson, R., Turner, D. L. and Butler, P. J.(1989). The relationship between diving capacity and oxygen storage capacity in the tufted duck (Aythya fuligula). J. Exp. Biol. 141, 265-275.
Takahashi, A., Dunn, M. J., Trathan, P. N., Sato, K., Naito, Y. and Croxall, J. P.(2003). Foraging strategies of chinstrap penguins at Signy Island, Antarctica: importance of benthic feeding on Antarctic krill. Mar. Ecol. Prog. Ser. 250, 279-289.
Tremblay, Y. and Cherel, Y.(2003). Geographic variation in the foraging
behaviour, diet and chick growth of rockhopper penguins. Mar. Ecol. Prog. Ser. 251, 279-297.
De Vries, J. and Van Eerden, M. R.(1995). Thermal conductance in aquatic birds in relation to the degree of water contact, body mass and body fat: energetic implications of living in a strong cooling environment. Physiol. Zool. 68, 1143-1163.
Walker, B. G. and Boersma, P. D.(2003). Diving behaviour of Magellanic penguins (Speniscus magellanicus) at Punta Tombo, Argentina. Can. J. Zool. 81, 1471-1483.
Wilson, R. P., Pütz, K., Bost, C.-A., Culik, B. M., Bannasch, R., Reins, T. and Adelung, D.(1993). Diel dive depth in penguins in relation to diel vertical migration of prey: whose dinner by candlelight? Mar. Ecol. Prog. Ser. 94, 101-104.