Model-driven elucidation of the inherent capacity of Geobacter sulfurreducens for electricity generation
Full text
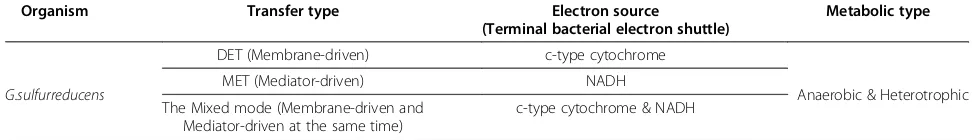
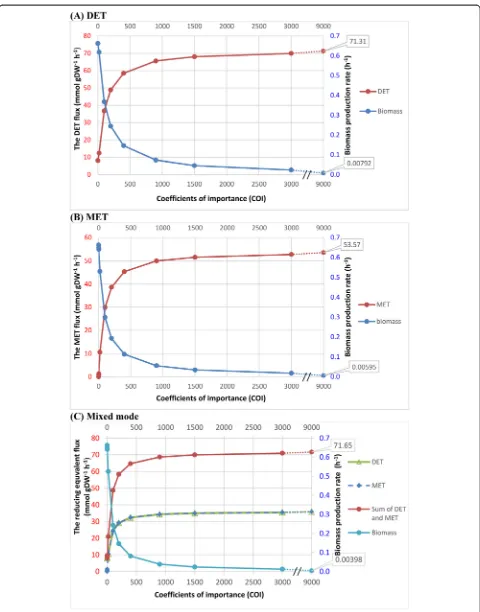
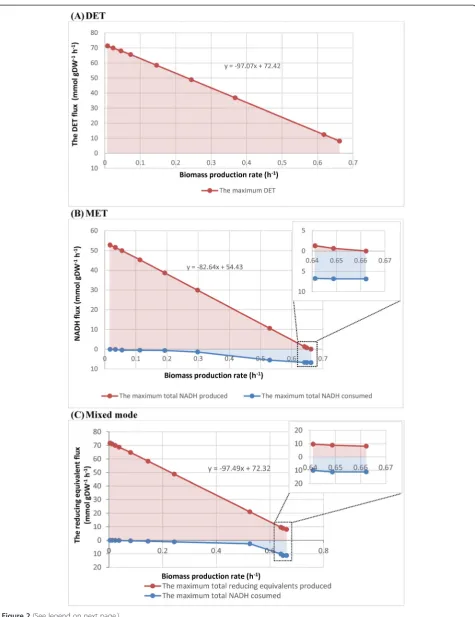
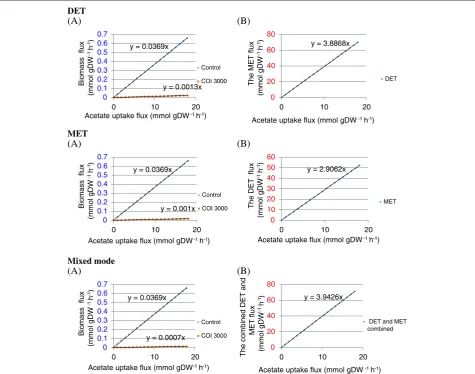
Figure



Related documents
Online 2455 3891 Print 0974 2441Vol 9, Suppl 2, 2016 A REVIEW ON ANTIOXIDANT METHODS SUNITHA DONTHA* Department of Pharmaceutical Chemistry, Malla Reddy College of Pharmacy,
F., Variance components and genetic parameters for milk production of Holstein cattle in Antioquia (Colombia) using random regression models, Rev. Gharahveysi, S., Irani
, 2015). In this study chemical kinetic study is carried out to understand the optimum time the reaction required to complete. In this study antioxidant property of
In the context of Web-page recommendation, the input data is Web logs that record user sessions on a daily basis. The user sessions include information about users‘ Webpage
Abstract—Reversible logic has become very promising for low power design using emerging computing technologies.Sequential circuits can be built by replacing the
Fishers from the villages in Lila, Pamilacan and Jagna in Bohol, those from Sagay and Guinsiliban in Camiguin and Magallanes in Limasawa hunted manta rays.. The fishers from Lila
Cell viability and cell attachment were studied by MTT assay and scanning electron microscopy (SEM). In vitro examinations revealed the amount of bone-like apatite
The study sought to examine whether knowledge influences the perception of customers towards non-interest banking services of Deposit Money Banks (DMBs) in Jimeta metropolis,
