Effects of adolescent binge ethanol exposure on striatal parvalbumin
By
Rayyanoor Jawad
Senior Honors Thesis Department of Chemistry
University of North Carolina at Chapel Hill
April 6, 2020
Approved:
Donita L. Robinson , Thesis Advisor Kevin Weeks, Reader
Acknowledgements
Firstly, I would like to thank the members of my committee, who took the time to review my thesis paper and participate in my defense, despite major drawbacks in the semester. My faculty advisor, Dr. Donita Robinson, provided me with this rich opportunity and offered unhindered guidance and support, but most importantly, trusted me to take on this project. I would like to thank and appreciate my postdoc mentor, Dr. Alex Gomez – A, without whom I could not have faced this wonderful challenge, who was present every step of the way with answers and comfort, and from whom I learned to think more critically about our study. My postdoc mentor, Dr. Carol Dannenhoffer, who enthusiastically taught me how to use the microscope, cell-count, slice brains, perform perfusions, and even how to do a bit of coding. I am eternally grateful for all the skills and training I received from her, including her gift of joy. Next, I would like to thank members of the Crews Lab, Dr. Ryan Vetreno, who graciously took the time to teach me how to perform IHC, use his equipment, and answer my plethora of questions about the staining process, and Dr. Victoria Macht, who would stop in her tracks to help me with any tangible or conceptual troubleshooting, a true expert on IHC and a great teacher. Last, but not least, Kate Spencer, a research technician in the Crews Lab, who answered all the questions I had while doing IHC staining and comforted me amidst my stress of getting the stain right the first time. The following work could not have been completed without these individuals.
Abstract
Introduction
According to a national survey, about 60% of college students from the ages of 18 to 22 years old reported drinking in the past month. About two-thirds of the drinking students reported binge drinking, which entails more than four drinks (NIAAA, 2015) and a blood ethanol concentration (BEC) ≥0.08 g/dl (HHS, 2006): the BEC value that is the legal alcohol driving-limit (Fell, 2014). Indeed, binge drinking constitutes the majority of drinking among high school and college students (NIAAA, 2015).
Adolescence, which is the period from peri-puberty to adulthood, is a critical period for brain development. During this time, the human brain undergoes substantial changes in the frontal and parietal regions where higher cognitive functions, such as cognitive control and planning ostensibly originate (Fuhrmann, 2015). The high level of brain plasticity during adolescence makes adolescents sensitive to alcohol and drugs (Ladouceur, 2012). Although some consequences of binge drinking are known, such as its link to accidents and violence, less is known about its consequences on adolescent brain development and how impairments in memory and cognition persist into adulthood.
Researchers in alcohol studies have found that the combination of low sensitivity to alcohol sedation, high risk-taking behavior, and reward seeking contributes to high rates of binge drinking in rodent and human adolescents (Crews et al, 2016; Spear and Swartzwelder, 2014). Studies show adolescent intermittent ethanol (AIE) exposure in rodents can alter synaptic physiology, sensitivity to alcohol, and behavior in ways that are reminiscent of an adolescent phenotype, even into adulthood (Crews et al, 2016; Spear and Swartzwelder, 2014).
a knockout of the δ subunit-containing GABAA receptor (δ-GABAAR) exhibit less ethanol consumption, less sensitivity to ethanol withdrawal, and less ethanol-induced seizure protection than wild-type mice. In the cerebral cortex, a subpopulation of interneurons is GABAergic. Some of the GABAergic interneurons contain the marker parvalbumin (PV), an intracellular Ca2+ binding protein that influences neuronal excitability and activity (Davies, 2003). In the striatum, PV interneurons regulate the activity of medium spiny neurons (MSNs), the principal neurons of the striatum, and the excitatory/inhibitory (E/I) balance to maintain the efficiency of cortico-striatal information processing driving appropriate behavioral responses (Lee, 2017).
al, 2019).
Although there is evidence of the deleterious effects of various models of alcohol exposure on different brain regions, the effects of adolescent alcohol exposure on PV neurons are unknown. Given the ability of perinatal alcohol exposure to reduce the expression of PV neurons in the striatum, and the ability of AIE exposure to impair behavioral flexibility and alter PV expression in the DG, the current study tested the overarching hypothesis that AIE-induced reductions in GABAergic PV interneuron markers are associated with AIE reductions in behavioral flexibility and functional connectivity. The findings from this study may inform future neurobiological targets for alcohol abuse disorder treatment in humans.
To compare PV interneuron marker levels in AIE and water (control) rats, we use immunohistochemistry (IHC), in which an unlabeled primary antibody binds to the target antigen in a cross-section of tissue. A secondary antibody reacts with the primary antibody and is labeled with a fluorescent dye (immunofluorescence method) or other tags, such as various enzymes (immunoenzyme method). The secondary antibody is typically derived from an animal that has been primed with immunoglobulin G (IgG) from the host of the primary antibody. This dual antibody method is sensitive due to amplification of the detective signal by binding multiple molecules of the second antibody to the primary antibody. Several factors may interfere with IHC results such as fluorescence, antibody binding capability, biotin, and several endogenous substances, such as endogenous peroxidase. Therefore, it is necessary to block background staining to avoid false positive staining (Chen et al, 2010).
Method
Animals
temperature and humidity-controlled vivarium with a 12:12 hour light cycle, lights on at 0700. Litters were culled to 10 pups by postnatal day (P) 3. Littermates were weaned to 2 per cage on P21. Adolescent ethanol was administered as described below (Madayag et al, 2017).
Adolescent Intermittent Alcohol (AIE) Paradigm
The exposure regimen used by the Neurobiology of Adolescent Drinking in Adulthood Consortium administered ethanol for 2 days, followed by 2 days of no exposure (5 g/kg ethanol, 25 percent v/v in water, postnatal day (P) 25–55) as a model for AIE to achieve binge-like BECs (>0.10 g/dL) (Broadwater et al, 2017). Rats were assigned in a balanced design to either AIE or water control. The first study consisted of 11 AIE and 9 water rats, and the second study consisted of 8 AIE and water rats each. This AIE regimen has been shown to produce a peak BEC value 242±15.4 mg/dl for males, occurring at 60 min. post-gavage in trunk and tail blood samples (Madayag et al, 2017).
Studies
Study 1
Previously stained brain tissue for PV ranged from about Bregma of 2.28 mm to -1.92 mm of the basal forebrain region, anterior to posterior. This sample consisted of young adult P80 brain tissue from control (n=9) and AIE (n=11) male rats.
Study 2
Perfusion, brain tissue preparation
Animal subjects were anesthetized with a lethal dose of urethane (1.5 g/kg) and transcardially perfused with 0.1 M phosphate-buffered saline (PBS, pH 7.4), followed by 4% paraformaldehyde in PBS. Brains were excised, and post-fixed in 4% paraformaldehyde for 24h at 4C and then, stored for 4 days in 30% sucrose solution.
Immunohistochemistry (IHC)
Brain samples from Studies 1 and 2 were used to perform full brain 5-parvalbumin immunoreactive (5-PV+IR) immunohistochemistry. The samples were treated with 20% glycerol and 2.0% dimethylsulfoxide to prevent freezing artifacts and using MultiBrain Technology, brains were embedded in a gelatin matrix. The MultiBrain blocks were sectioned coronally with 40 μm thickness and collected into 12-well plate containing cryoprotectant (30% glycol/30% ethylene glycol in PBS). The following procedures were administered while samples were on an orbital shaker. Samples were quenched with 0.6% H2O2 (30% stock) and washed with Phosphate-buffered saline (PBS) at room temperature (RT). To block endogenous peroxidase activity, free-floating MultiBrain sections were incubated in normal GOAT serum at RT. Sections were incubated overnight 4C in primary antibody rabbit anti-PV (Abcam – Cat. #ab11427). The following day, sections were washed with PBS, incubated in goat anti-rabbit IgG secondary antibody solution, washed again in PBS, and incubated in avidin-biotin complex (ABC) solution at RT. After washing again, immunoreactivity was visualized by staining sections with diaminebenzidine tetrahydrochloride (DAB) and H2O2 and washed again. Sections were mounted on gelatinized glass slides, air-dried, dehydrated in alcohol, cleared with xylene, and coverslipped.
For image capture and analysis, BioQuant Nova Advanced Image Analysis system (R&M Biometric, Nashville, TN) was used. Images were captured using an Olympus BX50 Fluorescence Microscope and Sony DXC-390 video camera linked to a computer. To ensure exactness of data acquisition, the instruments were background corrected and normalized to present light levels. Assessment of PV was performed in the striatal subregions: DMS, DLS, and VS defined in Fig. 3 and 4.
Statistical analysis
The total number of PV+IR cells was assessed in each striatal subregion and data were collected as the total number of cells per area for each animal (cells/mm2). The experimenter was blinded to the rat group. Unpaired t-tests were used to compare cell quantification between AIE and water-exposed rats. One-way analyses of covariance (ANCOVAs) were conducted to compare the effect of the covariate, striatal subregions, on PV+IR cell expression in both studies.
Results
Study 1
0.755. The one-way ANCOVA also suggested that the covariate, exposure, was non-significantly related to PV+IR expression, F(1) = 3.405, p = 0.071.
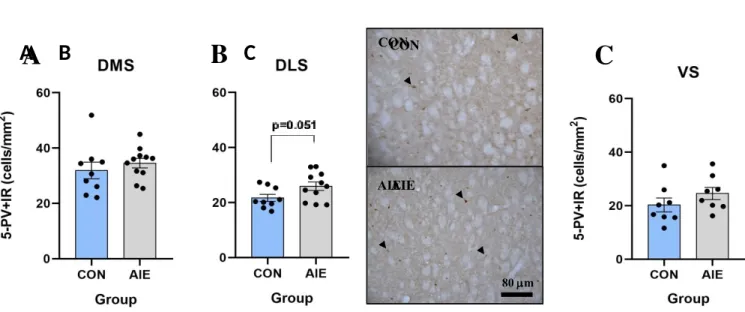
The photomicrographs in Fig. 1B are representative of their corresponding data. Looking at the photomicrograph in Study 1, there is a representative AIE-induced trend increase of PV+IR interneuron expression in the DLS region compared to the control.
Study 2
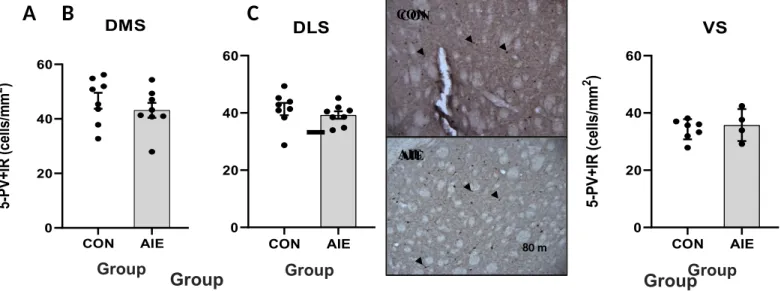
A replication study was necessary due to marginal effects found in Study 1. Results from Study 2 (Figure 2) show that AIE did not alter PV+IR interneurons in any striatal region. Specifically, AIE and control subjects did not differ significantly in the DMS [t(14) = 0.8769, p = 0.3953], DLS [t(14) = 0.8196, p = 0.4262], or VS [t(9) = 0.5528, p = 0.5939] PV+IR interneuron expression. The df value for the VS is less than that for DLS and DMS because there was a smaller sample size for the VS. Note that, the PV+IR cell count was higher across the striatum in Study 2 compared to Study 1. A one-way ANCOVA confirmed that the covariate, structure, was non-significantly related to PV+IR expression, F(1) = 2.769, p = 0.104. The one-way ANCOVA also confirmed that the covariate, exposure, was non-significantly related to PV+IR expression, F(1) = .346, p = 0.560.
To accurately measure PV+IR expression in Study 2 and avoid bias in the data, tissue that was not represented in both exposure groups within Study 2 was excluded and tissue was recounted randomly. The photomicrographs in Fig. 2B are representative of their corresponding data. Looking at Study 2’s photomicrographs, there is nearly an equal expression of PV+IR interneurons in the DLS.
The present study was the first to report the effects of AIE on striatal PV interneurons. Following analyses with unpaired t-tests in Studies 1 and 2, we observed that AIE-exposed rats had no significant effect on PV+IR interneuron expression in DMS, DLS, and VS subregions compared to the control. Additionally, one-way ANCOVA analyses revealed a non-significant main effect of the covariate, striatal subregion, on PV+IR expression.
We demonstrate here that PV interneurons in the DMS, DLS, and VS subregions are not affected by binge level alcohol exposure and are unlikely to play a significant role in the behavioral deficits associated with each subregion. According to Fig. 1, which shows the results from Study 1, we found that AIE does not significantly alter PV+IR interneuron expression in any striatal subregion. However, Fig. 1B indicated a trend increase in PV+IR interneuron count. These results suggested an increase in the feed-forward inhibition on MSNs in the DLS, a striatal subregion that mediates motor skill learning and conditioned responding to reward-associated stimuli. To test the robustness of the data, Study 2, a replication study, was conducted. The results from Study 2, shown in Fig. 2, revealed a non-significant main effect of exposure across the striatal subregions and no trend increase in the DLS region. Therefore, it is unlikely that AIE exposure altered PV+IR expression in the striatum.
amplifies the target signal due to having multiple binding sites. It is possible that other components of the IHC protocol may have differed to some extent between the two studies because they were conducted by different researchers. Furthermore, a fresh batch of primary antibody was used in Study 2, while an older batch was used in Study 1, which may have contributed to the darker stain and easy detectability of PV+IR interneurons.
Several limitations of the overall study must be addressed. The difference in IHC staining mentioned above may be presenting false positives in Study 2, however, given a consistently non-significant main effect of exposure and structure in both studies and low variability in the data, we deem this is unlikely. Another limitation is that some anterior sections of brain tissue that were stained and counted in Study 1 were not available to stain and count for Study 2, thus limiting the range of slices for Study 2. The limited tissue range was a reason for concern, because we observed that PV+IR interneuron expression was not homogeneous from the anterior to the posterior slices, and this was confirmed by a different study, which found that PV interneurons are more abundant rostrally than caudally in rat brains (Wu, 2000). Even though we are not able to directly compare both studies due to the lack of homogenous tissue representation from anterior to posterior, this was not the focus of the overall study. Another limitation is that the areas selected for each tissue slice while counting may have been inconsistent because the striatal subregions were manually divided into sections as shown in Figs. 3 and 4. To more accurately assess cells/mm2, a more systematic approach to the striatum’s division into its subregions could be implemented. One such approach is using Nikon’s NIS elements feature, which allows for automated and consistent area selection among slices.
alterations in specific striatal markers and correlate the results to behavioral changes. Another possibility for future projects could include the investigation of sex differences of striatal PV+IR interneuron expression after AIE exposure.
References
1. Broadwater, M. A., Lee, S.‐H., Yu, Y., Zhu, H., Crews, F. T., Robinson, D. L., and Shih, Y.‐
Y. I. (2018) Adolescent alcohol exposure decreases frontostriatal resting‐state functional connectivity in adulthood. Addiction Biology, 23: 810–823. doi:10.1111/adb.12530.
2. Chen, Xiao et al. “Double staining immunohistochemistry” North American journal of medical sciences vol. 2,5 (2010): 241-5.
3. Crews, Fulton T et al. “Adolescent Alcohol Exposure Persistently Impacts Adult Neurobiology and Behavior” Pharmacological reviews vol. 68,4 (2016): 1074-1109.
4. Davies, Martin. “The role of GABAA receptors in mediating the effects of alcohol in the central nervous system” Journal of psychiatry & neuroscience : JPN vol. 28,4 (2003): 263-74.
5. De Giorgio, Andrea et al. “Long-term alterations of striatal parvalbumin interneurons in a rat model of early exposure to alcohol.” Journal of neurodevelopmental disorders vol. 4,1 18. 3 Jul. 2012, doi:10.1186/1866-1955-4-18
6. Everitt, Barry J., and Trevor W. Robbins. "From the ventral to the dorsal striatum: devolving views of their roles in drug addiction." Neuroscience & Biobehavioral Reviews 37.9 (2013): 1946-1954.
7. Fell, James C, and Robert B Voas. “The effectiveness of a 0.05 blood alcohol concentration (BAC) limit for driving in the United States.” Addiction (Abingdon, England) vol. 109,6 (2014): 869-74. doi:10.1111/add.12365
9. Hamilton, G. F., et al. "Neonatal alcohol exposure reduces number of parvalbumin-positive interneurons in the medial prefrontal cortex and impairs passive avoidance acquisition in mice deficits not rescued from exercise." Neuroscience 352 (2017): 52-63.
10. Ladouceur, Cecile D et al. “White matter development in adolescence: the influence of puberty and implications for affective disorders.” Developmental cognitive neuroscience vol. 2,1 (2012): 36-54. doi:10.1016/j.dcn.2011.06.002
11. Lee, Kwang, et al. "Parvalbumin interneurons modulate striatal output and enhance performance during associative learning." Neuron 93.6 (2017): 1451-1463.
12. Lui, Wen and Crews, Fulton. “Persistent Decreases in Adult Subventricular and Hippocampal Neurogenesis Following Adolescent Intermittent Ethanol Exposure.” Frontiers in Behavioral Neuroscience vol.11 (2017): 151.
13. Madayag, Aric C et al. “Sex and Adolescent Ethanol Exposure Influence Pavlovian Conditioned Approach” Alcoholism, clinical and experimental research vol. 41,4 (2017): 846-856.
14. Melon, Laverne C et al. “Interneuronal δ-GABAA receptors regulate binge drinking and are necessary for the behavioral effects of early withdrawal” Neuropsychopharmacology reviews vol. 44 (2019): 425–434.
15. National Institute of Alcohol Abuse and Alcoholism. “College Drinking.” Turning Discovery Into Health (2015).
16. George, Paxinos, and Watson Charles. "The rat brain in stereotaxic coordinates." Qingchuan Zhuge translate 32 (2007).
18. Shashkova, Sviatlana and Mark C Leake. “Single-molecule fluorescence microscopy review: shedding new light on old problems” Bioscience reports vol. 37,4 BSR20170031. 21 Jul. 2017, doi:10.1042/BSR20170031
19. Spear, Linda Patia, and H Scott Swartzwelder. “Adolescent alcohol exposure and persistence of adolescent-typical phenotypes into adulthood: a mini-review.” Neuroscience and biobehavioral reviews vol. 45 (2014): 1-8. doi:10.1016/j.neubiorev.2014.04.012
20. U.S Department of Health & Human Services: National Institutes of Health. “Underage Drinking.” Alcohol Alert (2006).
21. Vetreno, Ryan P et al. “Adolescent intermittent ethanol reduces serotonin expression in the adult raphe nucleus and upregulates innate immune expression that is prevented by exercise” Brain, behavior, and immunity vol. 60 (2016): 333-345.
22. Vetreno, Ryan P and Fulton T Crews. “Adolescent binge ethanol-induced loss of basal forebrain cholinergic neurons and neuroimmune activation are prevented by exercise and indomethacin” PloS one vol. 13,10 e0204500. 8 Oct. 2018, doi:10.1371/journal.pone.0204500
23. Vongvatcharanon, Uraporn et al. “Alcohol administration during adulthood induces alterations of parvalbumin and glial fibrillary acidic protein immunoreactivity in rat hippocampus and cingulate cortex” Acta histochemical vol. 112,4 (2010): 392-401.
Figures
Figure 1. Striatal immunoreactive parvalbumin (PV+IR) expression in adolescent intermittent alcohol (AIE) and control (CON)- exposed rats (CON=9, AIE=11) in Study 1. Results are expressed as mean with standard error of mean (SEM), compared with unpaired t-tests, and represent combined left and right striatal regions. (A) AIE leads to no significant differences in PV+IR expressing interneurons in the dorsomedial striatum (DMS, p=0.4372). (B) AIE leads to a trend increase in PV+IR expressing interneurons in the dorsolateral striatum (DLS, p=0.051). Representative photomicrographs of the DLS are shown. The tissue is Bregma of 1.56 mm cross-sections from CON and AIE-exposed animals. Arrows point to a few of the representative cells selected for counting. (C) AIE leads to no significant differences in PV+IR expressing interneurons in the ventral striatum (VS, p=0.234).
C
B
CONAIE
A
A
B
C
CONAIE
CON AIE CON AIE 0 20 40 60 DMS 5-P V +I R (c el ls /m m 2 ) Group CON AIE 0 20 40 60 DLS 5-P V +I R (c el ls /m m 2 ) Group CON AIE 0 20 40 60 VS 5-P V +I R (c el ls /m m 2 )
Group Group Group
Figure 2. Striatal immunoreactive parvalbumin (PV+IR) expression in adolescent intermittent alcohol (AIE) and control (CON)- exposed rats (CON=8, AIE=8) in Study 2. Results are expressed as mean with standard error of mean (SEM), compared with unpaired t-tests, and represent combined left and right striatal regions. (A) AIE leads to no significant differences in PV+IR expressing interneurons in the dorsomedial striatum (DMS, p=0.3953). (B) AIE leads to no significant differences in PV+IR expressing interneurons in the dorsolateral striatum (DLS, p=0.4262). Representative photomicrographs of the DLS are shown. The tissue is Bregma of 1.56 mm cross-sections from CON and AIE-exposed animals. Arrows point to a few of the representative cells selected for counting. (C) AIE leads to no significant differences in PV+IR expressing interneurons in the ventral striatum (VS, p=0.5939).
A
B
C
80 m CON
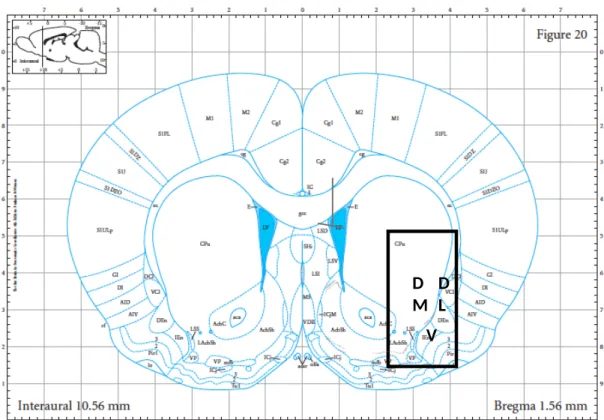
Figure 3. Coronal section of rat brain atlas of Paxinos and Watson (2007), showing dorsal medial striatum (left), dorsolateral striatum (right), and ventral striatum (bottom).
D M
D L
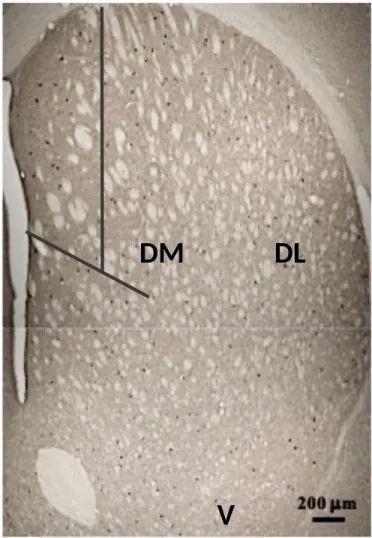
Figure 4. Coronal section of striatal tissue at 4 x 1 objective using an Olympus BX50 Fluorescence Microscope