RESEARCH ARTICLE
Akt1 signaling coordinates BMP signaling and
β
-catenin activity to
regulate second heart field progenitor development
Wen Luo1, Xia Zhao1, Hengwei Jin1, Lichan Tao1, Jingai Zhu1, Huijuan Wang1, Brian A. Hemmings2and Zhongzhou Yang1,3,*
ABSTRACT
Second heart field (SHF) progenitors exhibit continued proliferation and delayed differentiation, which are modulated by FGF4/8/10, BMP and canonical Wnt/β-catenin signaling. PTEN-Akt signaling regulates the stem cell/progenitor cell homeostasis in several systems, such as hematopoietic stem cells, intestinal stem cells and neural progenitor cells. To address whether PTEN-Akt signaling is involved in regulating cardiac progenitors, we deletedPtenin SHF progenitors. Deletion ofPtencaused SHF expansion and increased the size of the SHF derivatives, the right ventricle and the outflow tract. Cell proliferation of cardiac progenitors was enhanced, whereas cardiac differentiation was unaffected by Pten deletion. Removal of Akt1 rescued the phenotype and early lethality of Pten deletion mice, suggesting that Akt1 was the key downstream target that was negatively regulated by PTEN in cardiac progenitors. Furthermore, we found that inhibition of FOXO by Akt1 suppressed the expression of the gene encoding the BMP ligand (BMP7), leading to dampened BMP signaling in the hearts ofPtendeletion mice. Cardiac activation of Akt also increased the Ser552 phosphorylation ofβ-catenin, thus enhancing its activity. Reducing β-catenin levels could partially rescue heart defects ofPten deletion mice. We conclude that Akt signaling regulates the cell proliferation of SHF progenitors through coordination of BMP signaling andβ-catenin activity.
KEY WORDS: Second heart field, Akt, BMP,β-catenin, Mouse
INTRODUCTION
Lineage tracing and retrospective clonal analysis have identified two populations of cardiac progenitors during early mouse heart development (Kelly et al., 2001; Vincent and Buckingham, 2010; Watanabe et al., 2012). These two pools of cardiac progenitors are localized in the first heart field (FHF, or primary heart field) and the second heart field (SHF, or anterior heart field) (Bruneau, 2008; Harvey, 2002; Olson, 2006). Although the FHF contributes mainly to the left ventricle (LV), the SHF develops into the right ventricle (RV), the inflow tract and the outflow tract (OFT) (Black, 2007).
Starting at embryonic day (E) 8.5, the migration of SHF progenitors from the splanchnic mesoderm (SM) and pharyngeal mesoderm (PM) into the linear heart tube is essential for heart development in mice (Watanabe et al., 2012). Genetic studies in mice have revealed that disruption of SHF formation and
migration severely impaired heart development. For instance, each deletion ofIsl1,Tbx5,Mef2c andNkx2.5(Nkx2-5–Mouse Genome Informatics) affects the SHF development, resulting in developmental heart defects, with a single ventricle (the left ventricle) and the absence of SHF derivatives, including the RV and the OFT, being observed (Dodou et al., 2004; Hu et al., 2004; Lin et al., 1997; Prall et al., 2007; Xu et al., 2004).
SHF progenitors exhibit continued proliferation and a delay in differentiation (Srivastava, 2006; Watanabe et al., 2012).Fgf8and Fgf10, the first molecular marker of the murine SHF, are the two important regulators that promote SHF proliferation (Ilagan et al., 2006; Kelly et al., 2001; Park et al., 2006; Watanabe et al., 2010, 2012). Canonical Wnt/β-catenin signaling also drives SHF progenitor cell proliferation (Klaus and Birchmeier, 2009; Klaus et al., 2007; Kwon et al., 2007). Bone morphogenetic protein (BMP) signaling is required to induce the SHF formation and to subsequently inhibit cardiac cell proliferation (Klaus et al., 2007; McCulley et al., 2008; Prall et al., 2007).
PTEN-Akt signaling regulates the stem cell/progenitor cell homeostasis (Cully et al., 2006; Oudit and Penninger, 2009; Sussman et al., 2011; Walsh, 2006). In several stem/progenitor cell systems, such as hematopoietic stem cells, intestinal stem cells (ISCs) and neural progenitor cells, deletion ofPtencauses increased cell proliferation through Akt activation (Groszer et al., 2001; He et al., 2007; Li and Clevers, 2010).
To determine whether PTEN-Akt signaling is involved in the SHF regulation, we deleted Ptenin cardiac progenitor cells. We found that enhanced Akt signaling promoted SHF progenitor cell proliferation through the coordination of BMP signaling and
β-catenin activity.
RESULTS
Deletion ofPtenin the SHF progenitors results in SHF expansion and enlarged size of SHF derivatives
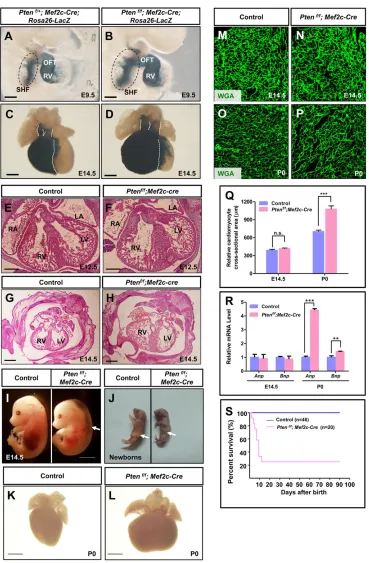
Mef2c-AHF-Cre(hereafter referred to asMef2c-Cre) mice express the Cre recombinase in the anterior heart field (AHF), a subset of the SHF which gives rise to OFT/RV (Verzi et al., 2005). This Cre line has been extensively utilized to delete genes for investigation of the SHF development in mice (Ai et al., 2007; Bai et al., 2013; Engleka et al., 2012; Xie et al., 2012). UsingMef2c-Cre,Ptenwas deleted in the SHF. Staining forβ-galactosidase clearly labeled the SHF and its derivatives, i.e. the OFT and the RV (Fig. 1A,B). Deletion ofPten resulted in a substantial expansion of SHF in the PM (Fig. 1A,B). The OFT and the RV in Pten deletion mice also showed significantly increased sizes compared with those of the control tissue (Fig. 1A,B). Quantitative analysis revealed that the SHF area (in the PM) and the length of the OFT inPten-deletion mice were significantly greater than in control mice (supplementary material Fig. S1A,B). Anatomical and histological analysis of embryos at later stages displayed significantly enlarged RV and arteries inPten Received 23 October 2014; Accepted 12 December 2014
1
MOE Key Laboratory of Model Animal for Disease Study, Model Animal Research Center, Nanjing Biomedical Research Institute, Nanjing University, Nanjing 210061, China.2Friedrich Miescher Institute for Biomedical Research, 4058 Basel, Switzerland.3Collaborative Innovation Center for Genetics and Development, Institute of Plant Biology, School of Life Sciences, Fudan University, Shanghai 200438, China.
*Author for correspondence ([email protected])
DEVEL
O
deletion mice (Fig. 1C-H). The heart function of these mice showed signs of impairment in the forms of edema (Fig. 1I). The majority of the mice died within 10 days after birth (Fig. 1S). Dead newborns were not presented with milk in the stomach or with hypertrophic heart (Fig. 1J-L). In addition, WGA staining showed no significant difference in cardiomyocyte size betweenPtendeletion and control mice by mid-gestational stage (E14.5) (Fig. 1M,N,Q), although at this stage cardiomyocyte proliferation was profoundly higher in Pten deletion mice than that of control mice (supplementary
material Fig. S1C-K). Furthermore, we observed enlarged sizes of cardiomyocytes in Pten deletion newborns compared with those of control mice (Fig. 1O-Q). Accordingly, expression of hypertrophic markers, such as Anp and Bnp (Nppa and Nppb, respectively–Mouse Genome Informatics), was not changed in the heart of Pten deletion mice at E14.5, but their levels were significantly increased inPtendeletion mice at birth (Fig. 1R).
[image:2.612.50.419.54.617.2]Taken together, these results indicate that PTEN inactivation affects SHF progenitors and SHF development.
Fig. 1. Deletion ofPtenin SHF progenitors resulted in SHF expansion.(A-D)β-galactosidase staining to visualize the SHF and its derivatives, the OFT and the RV, at E9.5 and E14.5, respectively. The SHF progenitors in the PM are circled by dashed lines in E9.5 mice. Deletion of
Ptencaused SHF expansion and enlarged size of the OFT and RV. (E-H) Histological analysis of the hearts at E12.5 and E14.5, respectively (H&E staining). (I) Gross analysis of mouse embryos. Arrow indicates edema. (J-L) Gross and microscopic analysis of embryos and their hearts. Arrows in J indicate the stomach. (M-P) WGA staining to display the cell size of cardiomyocytes at E14.5 and P0 (newborn). (Q) Quantification of cell size in M-P. (R) RT-qPCR analysis of gene expressions in heart tissue. (S) Survival curve. Scale bars: 50 µm.
DEVEL
O
Ptendeletion promotes the proliferation of SHF progenitor cells
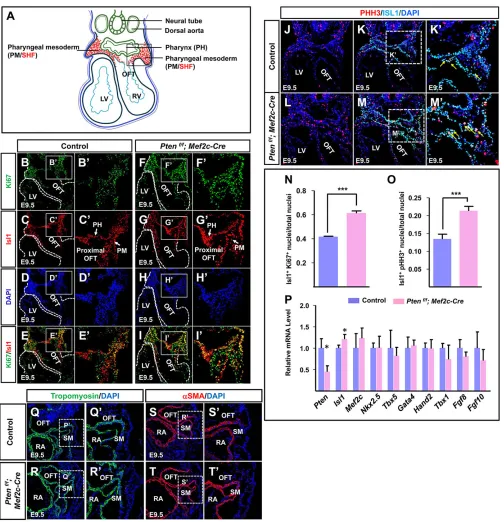
The enlarged SHF resulting fromPtenloss suggests enhanced cell proliferation of cardiac progenitors. Therefore, we assessed cell proliferation in the SHF. Islet1 (Isl1)-positive cells in the PM and proximal OFT indicate cardiac progenitor identity at E9.5. Thus, the proliferation of Isl1-positive cardiac progenitors in these regions was analyzed via immunofluorescence (IF) staining
(Cai et al., 2003). The results indicated that there were significantly more cells expressing Isl1 and the mitotic marker Ki67 (Mki67 – Mouse Genome Informatics) in Pten deletion embryos than in control embryos (Fig. 2A-I′,N). Co-staining of phosphorylated histone H3 ( pHH3) and Isl1 further confirmed these results (Fig. 2J-M′,O). In addition, loss ofPtenled to more
[image:3.612.57.561.152.674.2]β-gal- and Isl1-double-positive cells than in control animals (supplementary material Fig. S1L).
Fig. 2.Ptendeletion promotes the proliferation of SHF progenitors.(A) Schematic to display the horizontal section of E9.5 embryo. Boxed area marks the SHF for investigation (in red). (B-I) IF staining against Ki67/Isl1 of sections from E9.5 mouse embryos. Ki67/Isl1 double-positive nuclei are shown in yellow. B′-I′are higher magnification views of the boxed areas in B-I. (J-M) IF staining against PHH3/Isl1 of sections from E9.5 mouse embryos. PHH3/Isl1 double-positive nuclei are shown in yellow (indicated by arrows). K′and M′are higher magnification views of the boxed areas in K and M. (N) Quantification of Ki67/ Isl1 double-positive nuclei in B-I′. (O) Quantification of PHH3/Isl1 double-positive nuclei in J-M,K′and M′. (P) Gene expression levels examined by RT-qPCR. (Q-T) IF staining. Q′-T′are higher magnification views of the boxed areas in Q-T.
DEVEL
O
Next, we performed real-time quantitative PCR (RT-qPCR) to examine gene expression levels in the SHF region (Fig. 2A) of control and Pten deletion embryos. As expected, the expression levels ofPtenwere significantly decreased inPtendeletion mice compared with control mice at E9.5 (Fig. 2P). However, we observed profoundly enhanced Isl1 expression in Pten deletion mice compared with control mice, which was consistent with increased cell proliferation of SHF progenitors (Fig. 2P). The expression levels of the key cardiac transcription factors, including Nkx2.5,Hand2,Mef2c,Gata4,Tbx5 andTbx1, were comparable between control andPtendeletion mice (Fig. 2P). FGF8 and FGF10 are crucial signaling molecules promoting SHF progenitor proliferation, and their expression levels were not altered inPten
deletion mice (Fig. 2P). In addition, we investigated early cardiac muscle-specific genes, such as tropomyosin (Tpm) andα-smooth muscle actin (αSMA;Acta2–Mouse Genome Informatics) by IF staining and observed similar patterns in control andPtendeletion mice (Fig. 2Q-T′). These results suggest that cardiac differentiation was nearly normal inPtendeletion mice.
[image:4.612.77.530.55.530.2]PTEN is a negative genetic regulator of Akt1 signaling PTEN is a lipid phosphatase antagonizing the PI3K signaling, which mediates several signal transduction pathways, including the mitogen-activated protein kinase (MAPK) signaling pathway and the PDK1 signaling pathway (Cantley, 2002). PDK1 phosphorylates and subsequently activates a panel of kinases, such as Akt,
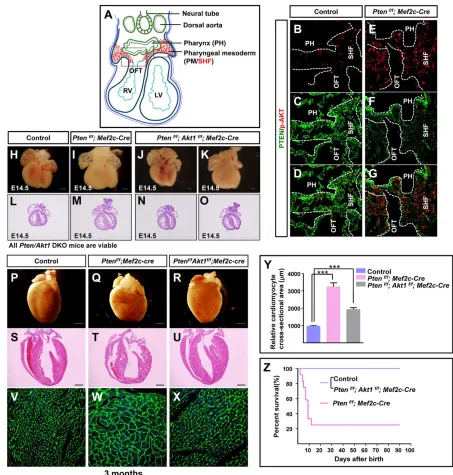
Fig. 3. PTEN is a negative genetic regulator of Akt1 signaling.(A) Schematic to display the horizontal section of E9.5 embryo. Boxed areas mark the SHF progenitor pool for investigation (in red). (B-G) IF staining against phospho-Akt S473 and PTEN. (H-O) Microscopic and histological analysis of mouse embryos and hearts. (P-U) Microscopic and histological analysis of mouse postnatal hearts. (V-X) WGA staining to display the cardiomyocyte size in mice of 3 months age. (Y) Quantification of cell size in V-X. (Z) Survival curve. Survival rate is identical between control andPtenf/f;Akt1f/f; Mef2c-Cre mice. Note that the data for the control andPtenf/f; Mef2c-Cre mice are from the same experiment as those shown in Fig. 1S.
DEVEL
O
PKC (Prkcg–Mouse Genome Informatics), SGK (Sgk1–Mouse Genome Informatics) and S6K (Rps6k – Mouse Genome Informatics) (Pearce et al., 2010). Although Akt is a key downstream target that is negatively regulated by PTEN, genetic evidence is lacking regarding the relationship between PTEN and Akt during mouse embryogenesis and organogenesis.
To examine whether Akt activity is promoted in the heart tissue of Ptendeletion mice, we performed IF staining in E9.5 embryonic hearts. Phosphorylation of Akt at serine 473 (S473) should indicate Akt activation, and IF staining of Akt phospho-S473 revealed a stronger signal in Pten deletion SHF than in that of controls (Fig. 3A-G).
Next, we tested the genetic relationship between PTEN and Akt during heart development. The deletion ofAkt1fromPtendeletion mice generated Pten/Akt1 double-deletion mice (Ptenf/f; Akt1f/f; Mef2c-Cre). Anatomic and histological analysis revealed comparable heart sizes betweenPten/Akt1double-deletion mice and control mice, indicating that the increased heart size ofPten deletion mice was rescued by abrogation of Akt1 (Fig. 3H-O). The majority of Pten deletion mice died within 10 days after birth and showed hypertrophic hearts. However, the Pten/Akt1 double-deletion mice were able to survive for more than 3 months and showed significantly reduced cardiomyocyte size compared withPten deletion mice, as evidenced by histological analysis (Fig. 3P-Z).
Similarly, we also observed that Mesp1-Cre-mediated Pten deletion (Ptenf/f; Mesp1-Cre) caused heart defects and embryonic
lethality beginning at E9.5, and complete lethality by E12.5. However, the deletion ofAkt1fromPtendeletion mice generated thePten/Akt1 double-deletion mice (Ptenf/f; Akt1f/f; Mesp1-Cre), which were able to develop to full term, with some progeny surviving for 3-4 months (supplementary material Fig. S2A,D-G). Furthermore, we found that deletion ofAkt2orAkt3also delayed the embryonic lethality ofPten deletion mice (supplementary material Fig. S2B,C). Histological analysis revealed nearly normal heart development in Pten/Akt1 double-deletion mice, although a slightly enlarged heart was observed (supplementary material Fig. S2A).
Taken together, these results indicate that Akt1 is the key downstream target of PTEN and is negatively regulated by PTEN during heart development. The increased size of the SHF and the defective SHF development found in Pten deletion mice are a consequence of enhanced Akt activity.
PTEN-Akt signaling suppresses BMP-SMAD signaling through FOXO
[image:5.612.86.527.56.417.2]BMP signaling is required for cardiac progenitor specification, proliferation and differentiation (Brand, 2003; Klaus et al., 2012; Mercola et al., 2011; Olson and Schneider, 2003; van Wijk et al., 2007). To understand how enhanced Akt activity gives rise to SHF expansion, we studied BMP signaling in the SHF ofPtendeletion mice. IF staining of phospho-SMAD1/5, an indicator of the activation of BMP signaling, showed a markedly reduced signal in the SHF ofPtendeletion hearts (Fig. 4A-I).
Fig. 4. PTEN-Akt signaling suppresses BMP-SMAD signaling.(A-H) IF staining against phospho-SMAD1/5 in the SHF at E9.5. (A′-H′) Higher magnification views of the boxed areas in A-H. (I) Quantification of p-SMAD1/5 nuclei in the SHF of A-H′. (J) Western blot analysis. (K,L) RT-qPCR analysis ofBmpligands and receptors.
DEVEL
O
Western blot analysis also revealed a consistent change in phospho-SMAD1/5 (Fig. 4J; supplementary material Fig. S3A,B). Among the∼20Bmpligands,Bmp2,4,7and10are expressed in the SHF (Liu et al., 2004; Wang et al., 2010; Yuasa and Fukuda, 2008). We examined the expression levels of theseBmpligands and their receptors in the SHF via RT-qPCR, and the results revealed significantly reduced levels of Bmp7 in Pten deletion mice compared with those of the controls (Fig. 4K). However, the expression levels of BMP receptors, including Alk1-3, Alk6and Bmpr2, were comparable between control andPtendeletion mice (Fig. 4L).
Collectively, these data indicate that BMP signaling is dampened in the SHF ofPtendeletion embryos.
Akt1 signaling was hyperactivated uponPten deletion, and we observed nuclear localization of active Akt. Therefore, we hypothesized that the expression ofBmp 7was suppressed by Akt signaling at the transcriptional level. The forkhead box O (FOXO) family of transcription factors are well-established Akt substrates,
and Akt signaling might repress the expression of BMP signaling components through inhibition of FOXO (Evans-Anderson et al., 2008). To test this hypothesis, we first investigated the phosphorylation status of FOXO in heart tissue via IF staining. The levels of FOXO phosphorylation in thePtendeletion hearts were profoundly enhanced compared with control hearts (Fig. 5A-G). Phosphorylation of FOXO by Akt translocates FOXO from the nucleus to the cytosol. Accordingly, we observed more cytosolic phospho-FOXO in the Ptendeletion hearts than in control hearts (Fig. 5A-G; and supplementary material Fig. S4). Furthermore, western blot analysis revealed significantly enhanced FOXO phosphorylation levels in the heart ofPtendeletion mice compared with control mice (Fig. 5H).
[image:6.612.90.516.55.479.2]To test whether FOXO activates the transcription ofBmp7, we ligated a luciferase reporter into the upstream cis-elements of the Bmp7gene, which contains a FOXO-binding consensus sequence (Fig. 5I). This luciferase reporter assay showed that constitutively active FOXO1 (FOXO1-AAA) strongly activated the transcription
Fig. 5. Akt regulatesBmp7expression through the FOXO transcription factor.(A-F) IF staining against phospho-FOXO1/3 in the SHF at E9.5. More pFOXO1/3 is localized in the cytosol ofPtendeletion mice. C′,F′are higher magnification views of the boxed areas in C,F. (G) Quantification of pFOXO1-positive nuclei. (H) Western blot analysis. (I)Bmp7-luciferase construct. There is a FOXO binding site at the−637 bp position in theBmp75′-upstream region. (J) Luciferase reporter assay. In theBmp7mutant, the FOXO binding motif of GTAAATAT was changed to GTACCTAT (highlighted in red).
DEVEL
O
of the luciferase reporter, whereas mutation of the FOXO-binding consensus sequence blocked this activation (Fig. 5I,J).
Taken together, these results demonstrate that Akt signaling suppresses BMP signaling through phosphorylation and inhibition of FOXO.
PTEN-Akt signaling regulatesβ-catenin activity
Wnt signaling regulates early cardiogenesis and is involved in SHF development. Deletion of the Wnt effectorβ-catenin in the SHF caused impaired RV formation (Kwon et al., 2007). It was previously reported that phosphorylation of β-catenin at the C-terminal S552 by Akt promotes β-catenin nuclear localization and activation in ISCs (He et al., 2007). We first investigated
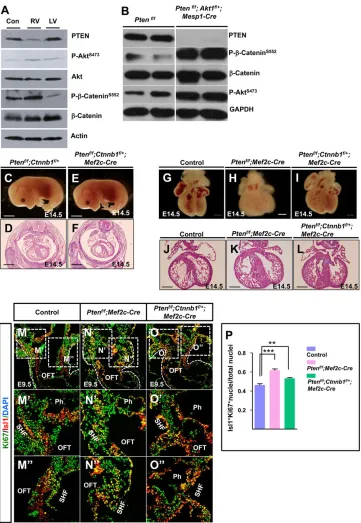
β-catenin phosphorylation levels in heart tissues through western blot analysis and found much higher phospho-β-catenin levels in the hearts of Ptenf/f; Mef2c-Cre mice than in control hearts (Fig. 6A). Ptenf/f; Akt1f/+; Mesp1-Cre mice also showed strong phosphorylation ofβ-catenin in the heart (Fig. 6B).
Next, we tested whether reducing β-catenin activity could reverse the phenotype of Pten deletion mice. We found that partial deletion of β-catenin partially reduced the heart size of Pten deletion mice (Fig. 6C-L) and also decreased cell proliferation of the SHF progenitors of Pten deletion mice (Fig. 6M-P).
[image:7.612.50.411.55.578.2]Collectively, these results showed that β-catenin activity was elevated inPtendeletion mice, which disrupted heart development.
Fig. 6. PTEN-Akt signaling regulates
β-catenin activity.(A,B) Western blot analysis ofβ-catenin S552 phosphorylation in heart tissues. (C-L) Microscopic and histological analysis of embryos and their hearts. (M-O) IF staining against Ki67/Isl1 at E9.5. M′-O′and M″-O″are higher magnification views of the boxed areas in M-O. (P) Quantification of Isl1/Ki67 double-positive nuclei in M-O″.
DEVEL
O
Reducing Akt signaling in SHF progenitors
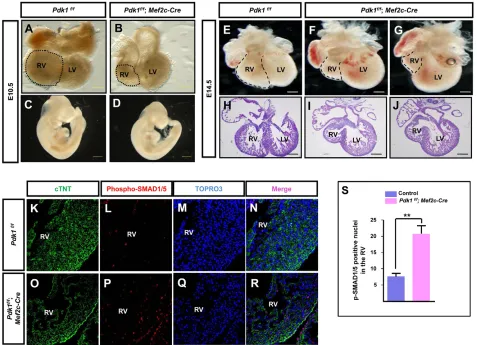
PDK1 is indispensable for Akt activation during heart development (Feng et al., 2010). We deleted Pdk1 in the SHF and observed impaired development of the RV and the OFT (Fig. 7A-J). IF staining of phospho-SMAD1/5 revealed an increased signal in the hearts of Pdk1deletion mice compared with the controls (Fig. 7K-S).
DISCUSSION
Our findings provide new insights into SHF development, which is a fundamental, but poorly understood, developmental event that governs OFT formation and ventricular expansion in mammals. We found that enhancing Akt signaling throughPtendeletion caused expansion of the SHF and overgrowth of SHF derivatives, the OFT and the RV, whereas the inhibition of Akt signaling upon PDK1 and Akt inactivation results in a smaller OFT and RV. Thus, our findings demonstrate a crucial role of Akt signaling in SHF development. Furthermore, we uncovered a mechanism underlying the coordination of BMP and Wnt-β-catenin signaling to regulate SHF proliferation and differentiation (supplementary material Fig. S5A).
Genetic relationship between PTEN and Akt1 in SHF development
One property of SHF progenitors is continued cell proliferation (Watanabe et al., 2012). FGF signaling drives the cell proliferation of SHF progenitors. Among the numerous FGF ligands, FGF8 and 10 are important regulators involved in SHF development. FGF10
was the first identified molecular marker of the murine SHF, and mouse genetic studies have revealed that both FGF8 and FGF10 promote SHF progenitor proliferation through activation of the MAPK/ERK pathway (Watanabe et al., 2010). In a variety of mouse and human tissues, FGFs exert their functions by modulating two pivotal signaling pathways: the MAPK/ERK signaling pathway and the PI3K-Akt signaling pathway. Therefore, FGF ligands, including FGF4, 8 and 10, are most likely the upstream intercellular growth factors that activate PI3K-Akt signaling in the SHF.
There are three Akt proteins in mice and humans, Akt1, 2 and 3, and previous studies by our group have demonstrated that all of these proteins are expressed in embryonic heart tissues (Chang et al., 2010). Although deletion ofAkt1 is sufficient to suppress tumor development inPten+/−mice, genetic evidence of the relationship between PTEN and Akt1 during mouse embryonic development is lacking. In this study, we demonstrated that Akt1 is the key downstream target that is negatively regulated by PTEN, and we established the first genetic relationship between PTEN and Akt1 in the SHF and heart development.
[image:8.612.68.545.59.404.2]Pten is expressed throughout embryogenesis, and germline deletion of Pten causes embryonic lethality by E9.5, indicating that PTEN is essential for mouse embryonic development (Suzuki et al., 1998). Thus, it is possible that modulation ofPtenexpression or PTEN protein levels by signals other than FGF signaling impacts the SHF development.
Fig. 7. Reducing Akt signaling in SHF progenitors.(A-J) Microscopic analysis of E10.5 and E14.5 hearts and embryos. RVs are marked by dashed lines. (K-R) IF staining against phospho-SMAD1/5 in the RV at E14.5. (S) Quantification of p-SMAD1/5-positive nuclei in the RV.
DEVEL
O
Crosstalk between PTEN-Akt signaling and BMP signaling Previously, Li and colleagues reported that BMP suppresses PTEN phosphorylation that inactivates PTEN, resulting in enhanced PTEN activity in ISCs (He et al., 2004). Their results indicate that BMP suppresses Akt activity through PTEN in ISCs. In this study, we found that Akt signaling represses the expression ofBmpligand (Bmp7) in SHF progenitors. As a result, BMP signaling is reduced by Akt activation. There might be a reciprocal antagonizing effect between Akt and BMP signaling in SHF development. Whether BMP inhibits Akt signaling in the SHF needs further investigation.
Regulation of the stem cell/progenitor cell switch from quiescent to active state
Based on their own and others’research on stem cells, Clevers and Li proposed a model for the switch of stem cells from a quiescent state to an active state (Li and Clevers, 2010). In the quiescent state, BMP signaling is active, and Wnt signaling is turned off in stem cells. Once stem cells are activated for proliferation (the active state), BMP signaling is shut off and Wnt signaling is switched on. Nevertheless, how the switch from a quiescent/BMP-on state to an active/Wnt-on state is regulated remains elusive. Our findings suggest that Akt signaling is a mediator coordinating both BMP and Wnt signaling during the transition state of stem/progenitor cells (supplementary material Fig. S5B). In the future, it will be intriguing to test this hypothesis in adult stem cells, such as hair follicle, gut and bone marrow stem cells.
We observed enlarged LVs of Pten deletion mice from late gestational stage. Lineage tracing showed that Pten-deletion cardiomyocytes moved from the RV to the LV, suggesting that the mutant cardiomyocytes have gained migratory ability or dominated the RV due to increased proliferation. This hypothesis deserves further investigation.
MATERIALS AND METHODS Mice
The previously described mouse strains used in this study includedPten conditional mice (Suzuki et al., 2001),Mesp1-CreandMef2c-AHF-Cremice (Klaus and Birchmeier, 2009; Saga et al., 1999; Verzi et al., 2005),β-catenin -floxed mice (Huelsken et al., 2001) andAkt1andPdk1 conditional mice (Di et al., 2012; Feng et al., 2010; Zhao et al., 2014). All mouse lines were maintained in a B6 genetic background. The experimental animal facility has been accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International (AAALAC), and the Institutional Animal Care and Use Committee (IACUC) of Model Animal Research Center of Nanjing University, China, approved all animal protocols used in this study.
Histological analysis andβ-galactosidase staining
Mouse embryos were collected and fixed in 4% paraformaldehyde, then dehydrated, embedded in paraffin and sectioned. Histological sections were stained with hematoxylin and eosin (H&E) or used for other analyses, such as immunohistochemistry (IHC), immunofluorescence (IF) staining and terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assays. For β-galactosidase staining, embryos were harvested and pre-fixed for 2 h in 4% paraformaldehyde on ice, and β-galactosidase staining was subsequently performed as previously described (Prall et al., 2007).
IF and IHC staining
Embryos were fixed in 4% paraformaldehyde for no more than 2 h. After fixation, the embryos were rinsed in PBS and incubated in 30% sucrose at 4°C for at least 8 h. The embryos were then embedded in OCT medium in the required direction. The embedded embryos were snap-frozen in liquid nitrogen and stored at−80°C. Sections were cut at a thickness of 7-10μm. Subsequently, the sections were blocked with 10% goat serum and
incubated with appropriately diluted primary antibodies at 4°C overnight. Afterwards, the sections were washed three times in PBS for 5 min each and then incubated with secondary antibodies or other dyes at room temperature for 2 h. The fixed and stained sections were mounted with nail polishing oil for analysis. IHC staining was carried out according to manufacturer’s recommendations and detected with 3,3-diaminobenzidine (DAB) (Maixin-Bio). The following antibodies were used for IF staining: anti-p-Akt (Cell Signaling Technology, CST 4060; 1:200), anti-p-Smad1/5 (Cell Signaling Technology, CST 9516; 1:200), anti-p-β-catenin S552 (Cell Signaling Technology, CST 5651S; 1:200), anti-Ki67 (DakoCytomation, M724901; 1:200), anti-p-Histone H3 (Cell Signaling Technology, CST 9701S; 1:200), anti-αSMA (Sigma, c6198; 1:200), anti-p-FOXO (Cell Signaling Technology, CST 2880; 1:200), and Isl1 (1:200) and anti-Tropomyosin (1:200) from the Developmental Studies Hybridoma Bank (DSHB; University of Iowa, Iowa City, USA).
Western blot analysis
Embryonic hearts were harvested, snap-frozen in liquid nitrogen and stored until use. Tissue lysates were prepared in lysis buffer as previously described (Chang et al., 2010). Approximately 50 µg of protein per sample was separated via electrophoresis and transferred to a polyvinylidene difluoride (PVDF) membrane (Millipore). The membranes were blocked with TBST (50 mM Tris, 150 mM NaCl, 0.5 mM Tween 20, pH 7.5) and incubated with primary antibodies overnight. The samples were analyzed via immunoblotting with antibodies against t-Akt (Bioworld, BS1810; 1:1000), p-Akt (Cell Signaling Technology, CST 4060; 1:1000), Smad1 (Cell Signaling Technology, CST 6944; 1:500), p-Smad1/5 (Cell Signaling Technology, CST 9516; 1:500), p-β-catenin552 (Cell Signaling Technology, CST 5651S; 1:500) and anti-GAPDH (Bioworld, AP0063; 1:20,000). The membranes were then incubated with a secondary anti-rabbit or anti-mouse HRP-conjugated antibody, and the resulting signals were detected through enhanced chemiluminescence (ECL).
RT-qPCR
Total RNA from embryonic hearts or pharyngeal arches was isolated with TRIzol reagent (Invitrogen), and 1μg of RNA was reverse-transcribed using a reverse transcription system kit (Promega). RT-qPCR was performed using an ABI7900 thermocycler with the Roche SYBR Green RT-qPCR reagent. The expression of selected genes involved in cardiac development was examined, and the results were normalized to the expression of GAPDH.
Plasmid construction and luciferase reporter assay
A∼3.4 kb fragment, corresponding to theBmp75′flanking sequence, was amplified via PCR from mouse genomic DNA and cloned into the pGL3-Basic Vector (Promega) to generate theBmp7-luc vector. TheBmp7mut-luc vector, which contains a 2-bp mutation in the mouseBmp7FHRE box (AA to CC), was generated using the MutaBest kit (Takara). Luciferase assays were performed using the Dual Luciferase Assay System (Promega), following a standard protocol. At least ten independent transfections were conducted, and all assays were performed three times.
Statistical analysis
All results are presented as mean±s.e.m. Statistical analyses were performed using Student’st-test with GraphPad Prism 4.03 software. Multiple groups were tested via one-way ANOVA, and comparisons between two groups were performed using Student’st-test. AP-value of <0.05 was considered significant (*), andP-values of <0.01 (**) or <0.005 (***) were considered highly significant.
Acknowledgements
We thank Dario Alessi forPdk1floxed mice, Brian Black forMef2c-AHF-Cre mice
and Yeguang Chen for discussions. We thank Teng Teng for English editing and Heng Wang for drawing schematics.
Competing interests
The authors declare no competing or financial interests.
DEVEL
O
Author contributions
W.L. and Z.Y. conceived and designed the experiments; W.L., X.Z., L.T., J.Z., H.W. and H.J. performed experiments and analyzed the data; W.L. and Z.Y. wrote the manuscript; W.L., X.Z., L.T., J.Z., H.W., H.J. and Z.Y. discussed and reviewed the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China [31130037, 91019002 and 31071282 to Z.Y.] and the National Key Basic Research Program of China [2011CB943904 and 2012CB966602 to Z.Y.]. The Friedrich Miescher Institute for Biomedical Research is part of the Novartis Research Foundation.
Supplementary material
Supplementary material available online at
http://dev.biologists.org/lookup/suppl/doi:10.1242/dev.119016/-/DC1
References
Ai, D., Fu, X., Wang, J., Lu, M.-F., Chen, L., Baldini, A., Klein, W. H. and Martin, J. F.(2007). Canonical Wnt signaling functions in second heart field to promote right ventricular growth.Proc. Natl. Acad. Sci. USA104, 9319-9324.
Bai, Y., Wang, J., Morikawa, Y., Bonilla-Claudio, M., Klysik, E. and Martin, J. F.
(2013). Bmp signaling represses Vegfa to promote outflow tract cushion
development.Development140, 3395-3402.
Black, B. L.(2007). Transcriptional pathways in second heart field development.
Semin. Cell Dev. Biol.18, 67-76.
Brand, T.(2003). Heart development: molecular insights into cardiac specification
and early morphogenesis.Dev. Biol.258, 1-19.
Bruneau, B. G.(2008). The developmental genetics of congenital heart disease.
Nature451, 943-948.
Cai, C.-L., Liang, X., Shi, Y., Chu, P.-H., Pfaff, S. L., Chen, J. and Evans, S.
(2003). Isl1 identifies a cardiac progenitor population that proliferates prior to
differentiation and contributes a majority of cells to the heart. Dev. Cell5,
877-889.
Cantley, L. C. (2002). The phosphoinositide 3-kinase pathway. Science 296, 1655-1657.
Chang, Z., Zhang, Q., Feng, Q., Xu, J., Teng, T., Luan, Q., Shan, C., Hu, Y., Hemmings, B. A., Gao, X. et al.(2010). Deletion of Akt1 causes heart defects
and abnormal cardiomyocyte proliferation.Dev. Biol.347, 384-391.
Cully, M., You, H., Levine, A. J. and Mak, T. W.(2006). Beyond PTEN mutations:
the PI3K pathway as an integrator of multiple inputs during tumorigenesis.Nat.
Rev. Cancer6, 184-192.
Di, R., Wu, X., Chang, Z., Zhao, X., Feng, Q., Lu, S., Luan, Q., Hemmings, B. A., Li, X. and Yang, Z.(2012). S6K inhibition renders cardiac protection against
myocardial infarction through PDK1 phosphorylation of Akt.Biochem. J.441,
199-207.
Dodou, E., Verzi, M. P., Anderson, J. P., Xu, S.-M. and Black, B. L.(2004). Mef2c is a direct transcriptional target of ISL1 and GATA factors in the anterior heart field
during mouse embryonic development.Development131, 3931-3942.
Engleka, K. A., Manderfield, L. J., Brust, R. D., Li, L., Cohen, A., Dymecki, S. M. and Epstein, J. A.(2012). Islet1 derivatives in the heart are of both neural crest and second heart field origin.Circ. Res.110, 922-926.
Evans-Anderson, H. J., Alfieri, C. M. and Yutzey, K. E.(2008). Regulation of cardiomyocyte proliferation and myocardial growth during development by FOXO transcription factors.Circ. Res.102, 686-694.
Feng, Q., Di, R., Tao, F., Chang, Z., Lu, S., Fan, W., Shan, C., Li, X. and Yang, Z.
(2010). PDK1 regulates vascular remodeling and promotes
epithelial-mesenchymal transition in cardiac development.Mol. Cell. Biol.30, 3711-3721.
Groszer, M., Erickson, R., Scripture-Adams, D. D., Lesche, R., Trumpp, A., Zack, J. A., Kornblum, H. I., Liu, X. and Wu, H.(2001). Negative regulation of neural stem/progenitor cell proliferation by the Pten tumor suppressor gene in
vivo.Science294, 2186-2189.
Harvey, R. P.(2002). Organogenesis: patterning the vertebrate heart.Nat. Rev. Genet.3, 544-556.
He, X. C., Zhang, J., Tong, W.-G., Tawfik, O., Ross, J., Scoville, D. H., Tian, Q., Zeng, X., He, X., Wiedemann, L. M. et al.(2004). BMP signaling inhibits intestinal
stem cell self-renewal through suppression of Wnt–β-catenin signaling. Nat.
Genet.36, 1117-1121.
He, X. C., Yin, T., Grindley, J. C., Tian, Q., Sato, T., Tao, W. A., Dirisina, R., Porter-Westpfahl, K. S., Hembree, M., Johnson, T. et al. (2007). PTEN-deficient intestinal stem cells initiate intestinal polyposis.Nat. Genet.39, 189-198.
Hu, T., Yamagishi, H., Maeda, J., McAnally, J., Yamagishi, C. and Srivastava, D.
(2004). Tbx1 regulates fibroblast growth factors in the anterior heart field through a
reinforcing autoregulatory loop involving forkhead transcription factors.
Development131, 5491-5502.
Huelsken, J., Vogel, R., Erdmann, B., Cotsarelis, G. and Birchmeier, W.(2001). beta-Catenin controls hair follicle morphogenesis and stem cell differentiation in the skin.Cell105, 533-545.
Ilagan, R., Abu-Issa, R., Brown, D., Yang, Y.-P., Jiao, K., Schwartz, R. J., Klingensmith, J. and Meyers, E. N.(2006). Fgf8 is required for anterior heart
field development.Development133, 2435-2445.
Kelly, R. G., Brown, N. A. and Buckingham, M. E.(2001). The arterial pole of the
mouse heart forms from Fgf10-expressing cells in pharyngeal mesoderm.Dev.
Cell1, 435-440.
Klaus, A. and Birchmeier, W.(2009). Developmental signaling in myocardial
progenitor cells: a comprehensive view of Bmp-and Wnt/β-catenin signaling.
Pediatr. Cardiol.30, 609-616.
Klaus, A., Saga, Y., Taketo, M. M., Tzahor, E. and Birchmeier, W.(2007). Distinct roles of Wnt/β-catenin and Bmp signaling during early cardiogenesis.Proc. Natl. Acad. Sci. USA104, 18531-18536.
Klaus, A., Müller, M., Schulz, H., Saga, Y., Martin, J. F. and Birchmeier, W.
(2012). Wnt/β-catenin and Bmp signals control distinct sets of transcription
factors in cardiac progenitor cells. Proc. Natl. Acad. Sci. USA 109,
10921-10926.
Kwon, C., Arnold, J., Hsiao, E. C., Taketo, M. M., Conklin, B. R. and Srivastava,
D.(2007). Canonical Wnt signaling is a positive regulator of mammalian cardiac
progenitors.Proc. Natl. Acad. Sci. USA104, 10894-10899.
Li, L. and Clevers, H.(2010). Coexistence of quiescent and active adult stem cells
in mammals.Science327, 542-545.
Lin, Q., Schwarz, J., Bucana, C. and Olson, E. N.(1997). Control of mouse cardiac
morphogenesis and myogenesis by transcription factor MEF2C.Science276,
1404-1407.
Liu, W., Selever, J., Wang, D., Lu, M.-F., Moses, K. A., Schwartz, R. J. and Martin, J. F.(2004). Bmp4 signaling is required for outflow-tract septation and
branchial-arch artery remodeling.Proc. Natl. Acad. Sci. USA101, 4489-4494.
McCulley, D. J., Kang, J.-O., Martin, J. F. and Black, B. L.(2008). BMP4 is required in the anterior heart field and its derivatives for endocardial cushion
remodeling, outflow tract septation, and semilunar valve development.Dev. Dyn.
237, 3200-3209.
Mercola, M., Ruiz-Lozano, P. and Schneider, M. D.(2011). Cardiac muscle
regeneration: lessons from development.Genes Dev.25, 299-309.
Olson, E. N.(2006). Gene regulatory networks in the evolution and development of
the heart.Science313, 1922-1927.
Olson, E. N. and Schneider, M. D.(2003). Sizing up the heart: development redux
in disease.Genes Dev.17, 1937-1956.
Oudit, G. Y. and Penninger, J. M.(2009). Cardiac regulation by phosphoinositide
3-kinases and PTEN.Cardiovasc. Res.82, 250-260.
Park, E. J., Ogden, L. A., Talbot, A., Evans, S., Cai, C.-L., Black, B. L., Frank, D. U. and Moon, A. M.(2006). Required, tissue-specific roles for Fgf8 in outflow
tract formation and remodeling.Development133, 2419-2433.
Pearce, L. R., Komander, D. and Alessi, D. R.(2010). The nuts and bolts of AGC protein kinases.Nat. Rev. Mol. Cell Biol.11, 9-22.
Prall, O. W. J., Menon, M. K., Solloway, M. J., Watanabe, Y., Zaffran, S., Bajolle, F., Biben, C., McBride, J. J., Robertson, B. R., Chaulet, H. et al.(2007). An
Nkx2-5/Bmp2/Smad1 negative feedback loop controls heart progenitor
specification and proliferation.Cell128, 947-959.
Saga, Y., Miyagawa-Tomita, S., Takagi, A., Kitajima, S., Miyazaki, J. and Inoue,
T.(1999). MesP1 is expressed in the heart precursor cells and required for the
formation of a single heart tube.Development126, 3437-3447.
Srivastava, D.(2006). Making or breaking the heart: from lineage determination to
morphogenesis.Cell126, 1037-1048.
Sussman, M. A., Völkers, M., Fischer, K., Bailey, B., Cottage, C. T., Din, S., Gude, N., Avitabile, D., Alvarez, R., Sundararaman, B. et al.(2011). Myocardial
AKT: the omnipresent nexus.Physiol. Rev.91, 1023-1070.
Suzuki, A., de la Pompa, J. L., Stambolic, V., Elia, A. J., Sasaki, T., del Barco Barrantes, I., Ho, A., Wakeham, A., Itie, A., Khoo, W. et al.(1998). High cancer susceptibility and embryonic lethality associated with mutation of the PTEN tumor suppressor gene in mice.Curr. Biol.8, 1169-1178.
Suzuki, A., Yamaguchi, M. T., Ohteki, T., Sasaki, T., Kaisho, T., Kimura, Y., Yoshida, R., Wakeham, A., Higuchi, T. and Fukumoto, M.(2001). T cell-specific loss of Pten leads to defects in central and peripheral tolerance.Immunity14, 523-534.
van Wijk, B., Moorman, A. F. M. and van den Hoff, M. J. B.(2007). Role of bone morphogenetic proteins in cardiac differentiation.Cardiovasc. Res.74, 244-255.
Verzi, M. P., McCulley, D. J., De Val, S., Dodou, E. and Black, B. L.(2005). The right ventricle, outflow tract, and ventricular septum comprise a restricted
expression domain within the secondary/anterior heart field.Dev. Biol. 287,
134-145.
Vincent, S. D. and Buckingham, M. E.(2010). How to make a heart: the origin and regulation of cardiac progenitor cells.Curr. Top. Dev. Biol.90, 1-41.
Walsh, K.(2006). Akt signaling and growth of the heart.Circulation113, 2032-2034.
Wang, J., Greene, S. B., Bonilla-Claudio, M., Tao, Y., Zhang, J., Bai, Y., Huang, Z., Black, B. L., Wang, F. and Martin, J. F.(2010). Bmp signaling regulates myocardial differentiation from cardiac progenitors through a MicroRNA-mediated
mechanism.Dev. Cell19, 903-912.
Watanabe, Y., Miyagawa-Tomita, S., Vincent, S. D., Kelly, R. G., Moon, A. M. and Buckingham, M. E.(2010). Role of mesodermal FGF8 and FGF10 overlaps in the
DEVEL
O
development of the arterial pole of the heart and pharyngeal arch arteries.Circ.
Res.106, 495-503.
Watanabe, Y., Zaffran, S., Kuroiwa, A., Higuchi, H., Ogura, T., Harvey, R. P., Kelly, R. G. and Buckingham, M.(2012). Fibroblast growth factor 10 gene regulation in the second heart field by Tbx1, Nkx2-5, and Islet1 reveals a genetic switch for
down-regulation in the myocardium.Proc. Natl. Acad. Sci. USA109, 18273-18280.
Xie, L., Hoffmann, A. D., Burnicka-Turek, O., Friedland-Little, J. M., Zhang, K. and Moskowitz, I. P.(2012). Tbx5-hedgehog molecular networks are essential in the second heart field for atrial septation.Dev. Cell23, 280-291.
Xu, H., Morishima, M., Wylie, J. N., Schwartz, R. J., Bruneau, B. G., Lindsay, E. A. and Baldini, A.(2004). Tbx1 has a dual role in the morphogenesis of the
cardiac outflow tract.Development131, 3217-3227.
Yuasa, S. and Fukuda, K.(2008). Multiple roles for BMP signaling in cardiac
development.Drug Discov. Today5, 209-214.
Zhao, X., Lu, S., Nie, J., Hu, X., Luo, W., Wu, X., Liu, H., Feng, Q., Chang, Z., Liu, Y. et al. (2014). Phosphoinositide-dependent kinase 1 and mTORC2 synergistically maintain postnatal heart growth and heart function in mice.Mol. Cell. Biol.34, 1966-1975.