Copyright2001 by the Genetics Society of America
Minimum Requirements for the Function of Eukaryotic
Translation Initiation Factor 2
F. Les Erickson,
1Joseph Nika,
1Scott Rippel and Ernest M. Hannig
Department of Molecular and Cell Biology, University of Texas at Dallas, Richardson, Texas 75083-0688 Manuscript received May 24, 2000
Accepted for publication February 9, 2001
ABSTRACT
Eukaryotic translation initiation factor 2 (eIF2) is a G protein heterotrimer required for GTP-dependent delivery of initiator tRNA to the ribosome. eIF2B, the nucleotide exchange factor for eIF2, is a heteropen-tamer that, in yeast, is encoded by four essential genes and one nonessential gene. We found that increased levels of wild-type eIF2, in the presence of sufficient levels of initiator tRNA, overcome the requirement for eIF2B in vivo. Consistent with bypassing eIF2B, these conditions also suppress the lethal effect of overexpressing the mammalian tumor suppressor PKR, an eIF2␣kinase. The effects described are further enhanced in the presence of a mutation in the G protein (␥) subunit of eIF2,gcd11-K250R, which mimics the function of eIF2Bin vitro.Interestingly, the same conditions that bypass eIF2B also overcome the requirement for the normally essential eIF2␣structural gene (SUI2). Our results suggest that the eIF2␥ complex is capable of carrying out the essential function(s) of eIF2 in the absence of eIF2␣and eIF2B and are consistent with the idea that the latter function primarily to regulate the level of eIF2·GTP·Met-tRNAiMetternary complexesin vivo.
I
N the current model for eukaryotic translation initia- eIF2 heterotrimer are encoded by the single-copy essen-tion (Hinnebusch and Liebman 1991; Merrick tial genes SUI2 (␣), SUI3 (), and GCD11 (␥). The 1992;Pain1996), initiator methionyl-tRNA is delivered primary structures of the eIF2 subunits are conserved to the 40S ribosomal subunit in the form of a eukaryotic between yeast and mammals (Ernstet al.1987;Dona-translation initiation factor 2 (eIF2)·GTP·Met-tRNAiMet hueet al. 1988;Pathaket al. 1988;Ciganet al. 1989; ternary complex. The resulting 43S complex, which also Ericksonet al.1997). The␥-subunit of eIF2 is a member includes eIF3 and eIF1A, binds at or near the 5⬘ end of the GTP-binding (G) protein superfamily and is of capped eukaryotic messenger RNAs in a process that highly similar to eubacterial elongation factor EF1A appears to involve interactions between eIF3 and pro- (formerly EF-Tu;Hanniget al.1993;Gasparet al.1994). teins bound at the mRNA 5⬘ cap (Sachs et al.1997). Functional similarity between EF1A and eIF2␥proteins Once bound, the ribosome traverses the mRNA in a 5⬘- is further suggested by genetic and biochemical data to-3⬘direction and locates the AUG codon representing that indicate the ␥-subunit plays a significant role in the translational start site. Recognition of the start site binding nucleotide and tRNA ligands (Erickson and is accompanied by GTP hydrolysis, which releases Met- Hannig1996). The guanine nucleotide exchange fac-tRNAiMetto the ribosomal peptidyl site and converts eIF2 tor for eIF2, eIF2B, is a heteropentamer that, in yeast, to an eIF2·GDP binary complex. The eIF2·GDP com- is encoded by four essential genes (GCD1,GCD2,GCD6, plex must be converted to the GTP form to rebind Met- and GCD7) and one nonessential gene (GCN3) that tRNAiMetand to participate in another cycle of transla- are also conserved in mammals (Bushmanet al.1993a; tion initiation. Mammalian eIF2·GDP is extremely stable Ciganet al. 1993;Koonin1995; Priceet al. 1996a,b).
in vitroin the presence of physiological concentrations The exchange reaction is regulated indirectly via phos-of magnesium and requires the guanine nucleotide ex- phorylation of the ␣-subunit of eIF2, which converts change factor eIF2B to promote rapid conversion of eIF2 into a competitive inhibitor of the exchange reac-eIF2·GDP to eIF2·GTP. Although the latter complex is tion (Rowlandset al.1988;Deveret al.1995;Kimball less stable, GTP binding is stabilized upon binding Met- et al. 1998). In vivo, this leads to reduced global rates
tRNAiMet. of translation initiation and, in some cases, gene-specific
In the yeast Saccharomyces cerevisiae, subunits of the enhancement of translation (Hershey 1991;
Hinne-busch 1993; Rhoads 1993). A mammalian eIF2␣
ki-nase, PKR, has been proposed to function as a tumor Corresponding author:Ernest M. Hannig, Department of Molecular suppressor and underlies the importance of regulating and Cell Biology, University of Texas at Dallas, Mailstop FO3.1, P.O.
the exchange reaction for normal homeostasis in higher Box 830688, Richardson, TX 75083-0688.
eukaryotic organisms (Koromilaset al.1992;Meurset
E-mail: [email protected]
1These authors contributed equally to this work. al.1993;Barberet al.1995a,b;Donzeet al.1995).
Klenow enzyme and ligation withHindIII linkers. This
frag-We previously described a mutation, gcd11-K250R,
ment was inserted into HindIII-cleaved pSB32 to create
that conferred phenotypes consistent with reduced eIF2
Ep1033. The GCD7 fragment was obtained from pJB100
function, i.e., reduced growth rates and increased ex- (Bushmanet al.1993a) by first converting the EcoRI site to pression ofGCN4(EricksonandHannig1996). This anEagI/NotI site using oligonucleotide linkers as described
above. A 2.1-kbNotI fragment containingGCD7was removed
mutation alters the lysine residue within the NKXD
nu-from the modified pJB100, ligated toEagI-cleaved YEp24 to
cleotide-binding motif of eIF2␥ that is conserved in G
create Ep673, and then subcloned as a 2.1-kbEagI fragment
proteins (Deveret al.1987;Bourneet al.1991). Both
intoEagI-cleaved Ep1033. The resulting plasmid, Ep1037, was
phenotypes were suppressed by increased dosage of the cleaved with AatII (in vector sequences), flush-ended, and yeast initiator tRNA gene (IMT). In vitro,gcd11-K250R converted to anAscI site using oligonucleotide linkers to create
Ep1066.
led to increased dissociation rates for both eIF2·GDP
The pSB32/GCD1/GCD2/GCD6/GCD7plasmid Ep1127 was
and eIF2·GTP, while the binding of Met-tRNAiMet to
constructed by inserting an AscI fragment containingGCD1
eIF2·GTP complexes stabilized GTP binding for both
andGCD2intoAscI-cleaved Ep1066. TheGCD1fragment was
the wild-type and ␥K250R forms of eIF2 (Erickson and obtained as a 2.4-kbp BamHI fragment from YCp50-Sc4014
Hannig1996). Together, these data suggested that the (Hill and Struhl 1988) that was initially subcloned into
BamHI-cleaved pBluescript (Promega, Madison, WI) to create ␥K250Ralteration might also promote more rapid
forma-Ep1082. TheGCD2fragment was obtained from pY26 (Dever
tion of ternary complexes and thereby reduce the
re-et al.1995) as a 2.6-kbp EagI fragment following conversion
quirement for eIF2Bin vivo.In this article, we confirm
of the ClaI site to an EagI site as above. This fragment was
these predictions. Furthermore, sufficient levels of wild- subcloned intoEagI-cleaved Ep1082. VectorXhoI andSacI sites type eIF2 also reduce the requirement for eIF2Bin vivo, were sequentially converted toAscI sites by linker tailing to
create Ep1120. A 5.0-kbAscI fragment from Ep1120 containing
though much less efficiently than in the presence of
GCD1andGCD2was then subcloned intoAscI-cleaved Ep1066
gcd11-K250R. This suggests that increasing the rate of
to create Ep1127.
dissociation of eIF2·GDP complexes is an essential
func-Additional plasmids containing the four essential eIF2B
sub-tion provided by eIF2B. Interestingly, the␣-subunit of unit genes, with or without GCD11
His8, were constructed as
eIF2, which appears to play an important role in eIF2/ follows. Ep1174 (pBluescript withXhoI andSacI sites altered eIF2B interactions (Vazquez de Aldana and Hinne- toAscI) was cleaved withBamHI andHindIII and ligated with a 2.1-kbBamHI/HindIII fragment containingGCD11His8.The busch1994;Pavittet al.1997, 1998;Nikaet al.2001),
resulting plasmid (Ep1246) was digested withBamHI and
li-is no longer required for viability under conditions that
gated with the 2.4-kbpBamHIGCD1fragment from
YCP50-lead to bypass of eIF2B. Our combined data suggest Sc4014. The resulting plasmid was cleaved withEagI and li-that the eIF2␥complex is capable of carrying out all gated with the 2.6-kbpEagIGCD2fragment to create Ep1247. essential eIF2 functions, including essential interactions A 7.1-kb AscI fragment, containing GCD1, GCD2, and GCD11His8, was ligated toAscI-cleaved Ep1067 and Ep1066 to
with additional components of the eukaryotic
transla-create Ep1250 (pRS316/GCD1/GCD2/GCD6/GCD7/GCD
tional machinery. We propose that eIF2B and the
11His8) and Ep1262 (pSB32/GCD1/GCD2/GCD6/GCD7/
␣-subunit of eIF2 comprise an elaborate regulatory
sys-GCD11His8), respectively. The pRS316/GCD1/GCD2/GCD6/
tem for modulating levels of ternary complex that, al- GCD7plasmid Ep1125 was constructed by first inserting the though not essential per sefor growth, plays a critical 2.1-kbEagIGCD7fragment from Ep673 intoEagI-cleaved pJB5 to create Ep1042. The 5.1-kbp GCD1/GCD2 AscI fragment
role in maintaining homeostasis and viability in
wild-from Ep1120 was then inserted at the SalI site in Ep1042,
type cells.
which had been modified usingAscI oligonucleotide linkers, to create Ep1125.
Plasmids used in the⌬sui2 suppression experiments con-MATERIALS AND METHODS
tained the 2.1-kbpHindIII/SnaBIGCD11fragment (where the SnaBI site was altered to aBamHI site by linker tailing;Hannig
Plasmids:pSB32 (LEU2), YCp50 (URA3;RoseandBroach
et al.1993) and/or a 2.5-kbp BamHI fragment from pD14-6 1991), and pRS316 (URA3;Sikorski andHieter1989) are
containing SUI2 (a gift from T. Dever, National Institutes yeast-Escherichia coli shuttle vectors containing yeast
centro-of Health). Construction centro-of YEp13/GCD11His8 (Ep832) and
mere sequences for maintenance in low copy number in yeast.
YEp13/gcd11-K250RHis8 (Ep921) were described previously
YEp24 (URA3), YEp13 (LEU2;RoseandBroach1991), and
(Erickson and Hannig 1996). YEp13/GCD11His8/SUI3
YEplac112 (TRP1; Gietz andSugino 1988) are yeast-E. coli
(Ep1064) and YEp13/gcd11-K250RHis8/SUI3 (Ep1065) were
shuttle vectors maintained in high copy in yeast due to the
constructed by removing the 2.5-kbpBamHISUI2 fragment presence of the 2 origin of replication. Construction of
from Ep847 and Ep922, respectively. YEp13/SUI2/SUI3/GCD11His8 (Ep847), YEp13/SUI2/SUI3/
Strain construction: The parent strain for EY878 (MAT␣ gcd11His8-K50R(Ep922), and YEplac112/IMT(Ep1013) were
leu2-3, 112 trp1-⌬63 ura3-52 gcd1::hisG gcd2::hisG gcd6⌬ described previously (Ericksonand Hannig1996). Unless
gcd7::hisG gcn3::hisG⬍Ep1125⬎) is EY809 (MAT␣leu2-3, 112 otherwise noted, theGCD11alleles used in this study contain
trp1-⌬63 ura3-52 gcd6⌬gcd7::hisG⬍Ep1042 (pRS316[URA3]/ a C-terminal octyl-histidine tag. The tag does not appear to
GCD6/GCD7⬎)). Thegcd7::hisGallele (from pJB110; Bush-alter the function of wild-type eIF2 in vivo or affect ligand
manet al.1993a) removes 66% (residues 75–325) of the 381-bindingin vitro(EricksonandHannig1996). The
galactose-amino-acidGCD7open reading frame (ORF). Thegcd6⌬allele induciblePKRconstruct (in YCp50) was a generous gift from
was from pJB96 (Bushmanet al.1993a), and removes 87% T. Dever (National Institutes of Health, Bethesda, MD).
(residues 93–713) of the 713-amino-acidGCD6ORF. The re-Ep1037 is a pSB32/GCD6/GCD7plasmid. A 3.3-kbSpeI
frag-mainder of the eIF2B subunit genes were deleted in EY809 ment from pJB6 (Bushman et al. 1993a) containing GCD6
LEU2plasmid Ep1127 that containsGCD1,GCD2,GCD6, and GCD7.Thegcd1⌬::hisG -URA3-hisGallele (Ep1191) was derived from Ep175 (a URA3 disruption version of Ep174;Hannig and Hinnebusch 1988) by replacing the disrupting URA3 fragment with the hisG -URA3-hisG cassette from pNKY51 (Alani et al. 1987). This removes 52% (residues 1–299) of the 578-amino-acid GCD1 ORF. The gcd2⌬::hisG -URA3-hisG allele (Ep1145) removes 93% (residues 26–632 on aPvuII/ EcoRI restriction fragment) of the 651-amino-acidGCD2ORF (Paddon et al.1989). The gcn3⌬::hisG -URA3-hisG allele was derived from Ep308 (Hannig et al.1990) by replacing the disrupting LEU2fragment with hisG -URA3-hisGas above to create Ep545. This construct removes the entire 305-amino-acidGCN3ORF. DNA fragments used for gene disruptions were obtained following digestion of the corresponding plas-mids withBamHI (GCD1),NotI (GCD2), orBglI/PvuII (GCN3)
Figure 1.—Effect of the␥K250R alteration on formation and purification by agarose gel electrophoresis. Gene
disrup-of ternary complexes by eIF2. The amount disrup-of [3H]Met-tRNA iMet tions were confirmed by Southern blot analysis of Ura⫹
trans-bound by eIF-2 was determined by nitrocellulose filter binding formants, using appropriate probes to distinguish
chromo-(EricksonandHannig1996) and reported as a percentage somal and plasmid-borne (in Ep1127) alleles (data not
of maximal Met-tRNAiMet bound. 䉭, wild-type eIF2; 䊐, shown). Chromosomal disruptants were plated on
5-fluoro-eIF2␥K250R;䉱, wild-type eIF2 and eIF2B;䊏, eIF2␥K250Rand eIF2B. orotic acid (5-FOA) medium to select for recombination
be-The data shown are the average of two experiments wherein tween the directhisGrepeats, an event that evicts theURA3
data points differ by⬍15% at each time point. GTP-indepen-gene, leaving behind a single copy of thehisGrepeat.GCD2,
dent [3H]Met-tRNA
iMetbinding was minimal (⬍0.1 pmol in GCD1, and GCN3 were disrupted sequentially in the EY809
all cases) and was not subtracted. Reactions were repeated in background. The URA3 plasmid Ep1125 was then used to
the absence of added GDP to determine maximal [3 H]Met-replace Ep1127 to create EY878. The absence of each essential tRNA
iMetbinding. The figure 100% corresponds to the follow-eIF2B subunit gene in EY878 was also demonstrated geneti- ing amount of [3H]Met-tRNA
iMetbound in each 5-l aliquot: cally by the inability of plasmids containing only three of the eIF2, 1.2 pmol; eIF2
␥K250R, 1.0 pmol; eIF2⫹eIF2B, 1.2 pmol; four essential eIF2B subunit genes, in all possible combina- eIF2
␥K250R⫹eIF2B, 1.1 pmol. tions, to complement in the absence of Ep1125.
Disruption ofGCD11in EY878 utilized a derivative of EY878 in which Ep1262(LEU2) replaced Ep1125(URA3).GCD11was
RESULTS then disrupted using thegcd11::hisG-URA3-hisG allele from
Ep523 as described (Dorriset al.1995), followed by growth Yeast eIF2 exhibits an intrinsic nucleotide exchange on 5-FOA. TheURA3plasmid Ep1250 was used to replace the activity:We measured nucleotide exchange activity by LEU2plasmid Ep1262 to create EY923.
testing the ability of GDP-bound yeast eIF2 to form
EY740 (MATa leu2-3, -112 ura3-52 trp1-⌬63 gcd11::hisG
eIF2·GTP·Met-tRNAiMetternary complexesin vitro (Fig-GAL2⫹⬍Ep293; YCp50/GCD11⬎) was obtained as a meiotic
segregant from a cross between aGAL2⫹derivative of H1515 ure 1). In mammalian systems, this reaction requires
(Martonet al.1993) and EY585 (MAT␣leu2-3, -112 ura3-52 eIF2B to dissociate the eIF2·GDP complex (reviewed in gcd11::hisG GAL2⫹⬍Ep293⬎). Thegcd11::hisGallele lacks the Merrick1992). In contrast to the mammalian system, entireGCD11open reading frame (Hanniget al.1993). EY835
wild-type yeast eIF2·GDP readily formed ternary
com-(MAT␣leu2-3, -112 ura3-52 trp1-⌬63 sui2⌬gcd11::hisG GAL2⫹
plexes in the absence of eIF2B, with 40% of maximal ⬍Ep1130; Ycp50/GCD11/SUI2⬎) was obtained from a cross
complex formation seen after 16 min. We attribute this
between EY740 and EY779 (MAT␣ leu2-3, -112 ura3-52
trp1-⌬63 sui2⌬ ⬍pSB32/SUI2⬎). The sui2⌬ allele removes the effect to an appreciable intrinsic GDP off-rate for yeast
N-terminal two-thirds of theSUI2ORF (Deveret al.1992). eIF2 that is not seen for the mammalian factor (Ahmad EY779 is a derivative of H1816 (Vazquez de Aldana and et al.1985; Erickson andHannig1996). Addition of Hinnebusch1994).
catalytic amounts of yeast eIF2B further increased ter-Growth rate determination:Doubling times at 30⬚were
de-nary complex formation by wild-type eIF2 toⵑ64% of
termined for log-phase cultures grown in minimal (SD) media
maximum over the same time period. In the absence
supplemented as necessary (Shermanet al.1986).
eIF2B assay:Reactions (30l) contained 20 mmTris-HCl of eIF2B, eIF2␥K250Rdisplayed an apparent rate of ternary (pH 7.5), 100 mmKCl, 5 mm MgCl2, 0.1 mm Na2EDTA, 1 complex formation (at early time points) that was mmdithiothreitol, 5% glycerol, 1 mg/ml creatine kinase (as greater than that for wild-type eIF2 alone and similar carrier), 10 m GDP, 100 m GTP, and 300 nm [3
H]Met-to the eIF2B-promoted reaction. The possibility that the
tRNAiMet(65 kcpm/pmol)3. eIF2 preparations (3g, ⬎80%
eIF2␥K250Rpreparation was contaminated with eIF2B is pure;Ericksonand Hannig1996) were prebound to GDP
for 10 min at 23⬚ in the absence or presence of 1 g yeast unlikely, since the addition of eIF2B to the eIF2␥K250R eIF2B (Nikaet al.2001) and then transferred to a 10⬚water reaction did not further enhance ternary complex for-bath for an additional 5 min. Under these conditions, both mation (i.e., reaction rates at early time points are simi-wild-type eIF2 and eIF2␥K250R are maximally bound to GDP lar). The ability of the␥K250Ralteration to mimic eIF2B (Erickson and Hannig 1996). GTP and [3H]Met-tRNA
iMet
activityin vitrosuggested thatgcd11-K250Rstrains might
were then added, and 5-l aliquots were withdrawn and
fil-demonstrate a reduced requirement for eIF2B function
tered through nitrocellulose as described previously
the absence of the high-copy IMT plasmid, all strains failed to grow upon induction of PKR expression on galactose media (data not shown). However, in the pres-ence of increased IMTgene dosage,gcd11-K250R sup-pressed the PKR-mediated growth defect in the pres-ence of elevated levels of eIF2␣ and eIF2(Figure 2), suggesting that suppression is mediated by the eIF2 complex. Under these same conditions wild-type eIF2 was a much weaker suppressor, as would be predicted on the basis of its slower intrinsic off-rate for GDP. eIF2␥Y142H, which demonstrates a wild-type GDP off-rate
in vitro(Erickson andHannig1996), suppresses at a level similar to wild-type eIF2. The effects of overexpres-sion of eIF2 are likely not due to inhibition of phosphor-ylation of the ␣-subunit by PKR, as these conditions have been shown to increase the level of phosphorylated
Figure 2.—Suppression of lethality associated with PKR
expression in yeast. Three plasmids were introduced into yeast eIF2 in the cell (Deveret al.1995). Our results suggest
strain EY740 (MATa ura3-52 trp1-⌬63 leu2-3 leu2-112 that elevated levels of eIF2 and initiator tRNA reduce gcd11::hisG GAL2⫹): (i) a high-copy LEU2plasmid (YEp13)
the requirement for eIF2B at a level proportional to the
containing genes encoding the ␣(SUI2), (SUI3), and
intrinsic nucleotide off-rate for yeast eIF2. ␥(GCD11) subunits of eIF-2, or a low-copy LEU2 plasmid
Bypass of the essential function of eIF2B:If the only
(pSB32) encoding the␥-subunit only; (ii) a high-copyTRP1
plasmid (YEplac112;GietzandSugino1988) containing the essential function provided by eIF2B is to promote the yeastIMTgene; and (iii) a low-copyURA3plasmid (YCp50; rapid dissociation of eIF2·GDP complexes, we reasoned RoseandBroach1991) with (PKR) or without (vector) the
thatgcd11-K250Rmight suppress a deletion of some or
mammalian PKR gene expressed from the inducible yeast
all of the four essential eIF2B subunit genes. To test
GAL1 promoter. Strains representing independent isolates
this hypothesis, we constructed a yeast strain lacking the
were grown to saturation in liquid minimal media containing 2% dextrose (Sherman et al. 1986), diluted, andⵑ104cells
chromosomal GCD1, -2, -6, and -7 genes that encode
were spotted onto minimal media plates containing either 2% the essential eIF2B subunits (Ciganet al.1991, 1993), dextrose or 2% galactose and were then incubated for 2 days
as well as the nonessential eIF2B subunit gene GCN3
at 30⬚.
(HannigandHinnebusch1988). The resulting strain
(EY878) is viable because it contains a low-copyURA3
plasmid harboring the four essential eIF2B subunit PKR-mediated growth inhibition is suppressed by
in-creased eIF2 gene dosage:We reasoned that yeast cells genes. To test the requirement for eIF2B ingcd11-K250R
strains, we constructedLEU2plasmids containing either less dependent upon eIF2B might show decreased
sensi-tivity to overexpression of the mammalian eIF2␣kinase gcd11-K250R or GCD11in combination with SUI2and
SUI3.These plasmids were introduced into EY878 that PKR, which confers a severe slow-growth phenotype in
wild-type yeast (Deveret al.1993). This phenotype ap- also harbored either a high-copy IMT plasmid or the empty vector. Transformants were then plated on 5-FOA pears to result from an increase in phosphorylated
eIF2␣(at residue serine 51), which converts eIF2 to a medium to examine the ability of the eIF2 plasmids to suppress the complete loss of essential eIF2B genes. competitive inhibitor of the exchange reaction (
Row-landset al.1988;Chonget al.1992;Deveret al.1995). 5-FOA selects for Ura⫺ cells that have lost the URA3
plasmid (Boekeet al.1987), which, in the case of EY878, Because eIF2B is typically present at reduced levels
rela-tive to eIF2 (PriceandProud1994), the resulting func- contains the only copies of the essential eIF2B subunit genes. As shown in Figure 3, the gcd11-K250R/SUI2/ tional sequestration of eIF2B leads to decreased levels
of ternary complex and thus reduced growth rates. We SUI3 combination was an effective suppressor in the eIF2B quintuple-deletion strain. Suppression was de-constructed yeast strains harboring a high-copy plasmid
containingGCD11orgcd11-K250Rin combination with pendent upon increasedIMTgene dosage and, in addi-tion, required high-copy expression of all three eIF2
SUI2 andSUI3, or a low-copy plasmid containing the
GCD11allele alone. Co-overexpression of the three eIF2 subunit genes (Figure 3; data not shown). Western blot analysis confirmed the absence of detectable eIF2B sub-subunit genes has been shown to increase the level of
this initiation factor complex at least fivefold in vivo units in the suppressed strain (data not shown). Overex-pression of wild-type eIF2 also suppressed the complete
(Hanniget al.1993; Deveret al.1995;Erickson and
Hannig1996). The chromosomalGCD11allele was de- absence of eIF2B in anIMT-dependent manner, albeit
at a reduced level, consistent with the intrinsic nucleo-leted in these strains, which, in addition, harbored a
plasmid containing a galactose-inducible PKR construct, tide off-rate seen for wild-type eIF2·GDP complexes. Enhanced dissociation of eIF2·GDP resulting from the as well as a high-copyIMTplasmid (or the empty vector).
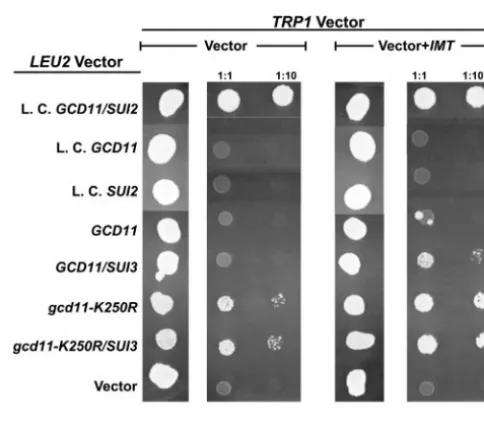
Figure 3.—Suppression of the lethal effect associated with deletion of the five eIF2B sub-unit genes. Low-copy (L.C.; pSB32) or high-copy (H.C.; YEp13) LEU2 plasmids indi-cated to the left of the figure were introduced into the eIF2B quintuple deletion strain EY878, which also harbored either YEplac112 [TRP1] (Vec-tor) or YEplac112/IMT (Vec-tor ⫹ IMT). Plasmid trans-formants were selected, grown in liquid media containing ura-cil, and 5l of a 1:10 dilution was spotted onto minimal media containing uracil⫹5-FOA (5-FOA) or uracil alone (SD⫹URA). Each spot corresponds to an independent isolate. This photograph was taken following incubation at 30⬚for 5 days. Low-copy LEU2plasmids harboring any combination of three of the four essential eIF2B subunit genes failed to complement in EY878 (data not shown).
more efficient bypass of the nucleotide exchange func- ␣-subunit to eIF2 function is not essential for ligand binding or the interaction of eIF2 with additional com-tion of eIF2B. Our results lend strong support to the
notion that the rapid dissociation of eIF2·GDP com- ponents of the translational apparatus.
Comparative requirements for bypass of essential plexes is an essential eIF2B function.
Bypass of the essential function ofSUI2 (eIF2␣):In eIF2B and eIF2␣functions:Examination of results pre-sented in Figures 3 and 4 reveals a difference in the cells that no longer require eIF2B, it is possible that
certain eIF2 subunit(s) with which eIF2B interacts are requirements for suppression in the eIF2B deletion not required. We chose the ␣-subunit of eIF2 to test
this idea, on the basis of previous genetic evidence that suggested a direct interaction between the␣-subunit of eIF2 and eIF2B (Vazquez de Aldanaet al.1993;Pavitt
et al.1997, 1998). We constructed a⌬gcd11⌬sui2strain (EY835) that harbored aURA3/GCD11/SUI2 plasmid and used the plasmid shuffle technique described above to examine the ability ofGCD11andgcd11-K250R con-structs (LEU2) to suppress the⌬sui2mutation by confer-ring viability in the absence of the residentURA3 plas-mid. To demonstrate the absence of chromosomalSUI2
andGCD11in this strain, a low-copyLEU2plasmid con-taining bothSUI2andGCD11, but not plasmids harbor-ing either gene alone, supported the viability of EY835 in the absence of theURA3/GCD11/SUI2plasmid (Fig-ure 4). High-copy plasmids containinggcd11-K250R, ei-ther alone or in combination withSUI3 (eIF2), sup-pressed the⌬sui2mutation, increasing doubling times 1.5- to 2.2-fold compared with controls (Figure 5, bot-tom panel). Suppression bygcd11-K250Rwas
indepen-dent of the presence of a multi-copyIMTplasmid, al- Figure4.—The␣-subunit of eIF2 is dispensable in
gcd11-K250Rstrains.LEU2plasmids harboring alleles listed to the
though suppression was more efficient with the IMT
left were introduced into yeast strain EY835 (MAT␣leu2-3, -112
plasmid (20–40% decrease in doubling times). This
re-trp1-⌬63 ura3-52 sui2⌬ gcd11::hisG ⬍YCp50/SUI2/GCD11⬎)
sult suggests that the two-subunit form (␥) of eIF2
containing either YEplac112 (Vector) or YEplac112/IMT
is functional in this strain, but does not rule out the (Vector⫹IMT). Transformants were grown in minimal media additional possibility that the ␥-subunit alone is func- containing uracil for 2 days at 30⬚, plated as described in
Figure 2 (either undiluted or at a 1:10 dilution) on minimal
tional. A low-copy plasmid containinggcd11-K250Ralso
media (SD) containing uracil and 5-FOA (5-FOA) or uracil
suppressed ⌬sui2, albeit less efficiently (data not
alone (SD⫹URA), and grown for 4 days at 30⬚. Results are
shown). Overexpression of wild-typeGCD11weakly
sup-representative of analysis of a larger number of independent
pressed ⌬sui2 (5-fold increase in doubling time) and transformants (data not shown). L.C., plasmids based on the suppression required co-overexpression of SUI3 and low-copy vector pSB32; the remainder of the plasmids listed
to the left are based upon the high-copy vector YEp13.
⌬gcd11strain (EY923) isogenic with EY878 and repeated the eIF2B bypass experiments. The results, shown in Figure 6, are essentially identical to those shown in Figure 3;i.e., bypass of the essential function of eIF2B requires overexpression of both eIF2 and initiator tRNA and is independent of the presence of a chromosomal
GCD11allele in the host strain. Again, eIF2␥K250Ris a more efficient suppressor, resulting in a 1.4-fold increase in doubling time (vs.the control) compared with a 3-fold increase for wild-type eIF2 (Figure 5, top).
DISCUSSION
Previous biochemical studies using mammalian
fac-Figure5.—Growth rates of suppressed strains lacking eIF2B tors indicated that eIF2 and eIF2B play critical roles in
or eIF2␣. Growth rates were determined in EY923 (eIF2B the initiation of eukaryotic protein synthesis (reviewed bypass; see Figure 6) or EY835 (eIF2␣ bypass; see Figure 4) inMerrick1992;PriceandProud1994;Pain1996). following growth on 5-FOA as described inresults.Low-copy
Genetic analyses in yeast provided evidence that both
(L.C.) or high-copy (H.C.) LEU2 plasmids present in each
factors are required for growth and viability (
Hinne-strain containing eIF2 or eIF2B subunit genes are indicated
to the left. All strains contained a high-copy TRP1vector or busch1997). In yeast, each of the three conserved eIF2
the same vector containingIMTas indicated. Doubling times subunits and four of the five conserved eIF2B subunits were determined in triplicate as described inmaterials and are encoded by essential genes. We previously described methods.
a mutation in the gene encoding the␥-subunit of eIF2,
gcd11-K250R, that increased the intrinsic rate of dissocia-tion of guanine nucleotides from binary complexes in
strain compared with the ⌬sui2 strain. In the former
instance, suppression in all cases required increased vitro(EricksonandHannig1996). Although eIF2␥K250R showed increased dissociation for both GDP and GTP
IMT gene dosage, whereas suppression of ⌬sui2 by
gcd11-K250R is independent of, though enhanced by, in vitro, GTP binding by both eIF2␥K250R and wild-type eIF2 could be stabilized by forming ternary complexes the presence of additional copies ofIMT.A trivial
expla-nation for this difference may be related to the presence with charged initiator tRNA. These results were consis-tent with in vivo experiments that demonstrated in-of the chromosomalGCD11allele in EY878 used in the
eIF2B bypass experiments (Figure 3). In this case, the creased IMT gene dosage suppressed both the slow growth and increased expression ofGCN4(i.e., the Gcd⫺ presence of wild-type eIF2 complexes may compete with
eIF2␥K250Rand thereby reduce the efficiency of suppres- phenotype) ingcd11-K250Rstrains. The latter are indic-ative of at least partial restoration of eIF2 function (
Hin-sion in these strains. To test this idea, we created a
nebusch1994). These combined results suggested to in gene dosage requirements forIMTraises the possibil-us that gcd11-K250R strains might exhibit a reduced ity that eIF2B provides a function, in addition to nucleo-dependence upon eIF2B, resulting in partial or com- tide exchange, that is substituted (in the eIF2B bypass plete bypass of the requirement for eIF2B dependent experiments) by elevated levels of initiator tRNA. It is upon (or enhanced by) increasedIMTgene dosage. The possible that catalyzed nucleotide exchange proceeds data presented here demonstrate that overexpression of through an eIF2·GTP·eIF2B intermediate that facili-either the wild-type or ␥K250R form of eIF2 suppresses tates the interaction of eIF2 with initiator tRNA, perhaps deletion of the four essential eIF2B subunit genes and by increasing the on-rate for tRNA relative to eIF2·GTP that bypass of essential eIF2B function(s) requires co- binary complexes.ManchesterandStasikowski(1990) overexpression of initiator tRNA. Consistent with a re- proposed a similar model based upon theoretical con-duced requirement for eIF2B function, strains grown siderations of association and dissociation rate constants under bypass conditions but containing all eIF2B sub- under physiological conditions and the reaction rates unit genes show a reduced sensitivity to the eIF2␣ ki- for protein synthesis initiation. If this is indeed the pre-nases PKR (Figure 2) and Gcn2p (Dever et al.1995). ferred pathway for ternary complex formationin vivo, The efficiency of suppression of both the PKR-induced increasing the level of initiator tRNA may overcome the growth phenotype and the absence of eIF2B correlated requirement for this eIF2B function via mass action. directly with the rate of dissociation of guanine nucleo- However, this model does not fully explain the require-tides determined previously with purified eIF2 prepara- ment for increased IMT gene dosage in gcd11-K250R tions;i.e.,gcd11-K250Rstrains were more efficient than strains in the absence of eIF2B. In the presence of eIF2B, strains harboring the wild-typeGCD11allele. Our data the viability ofgcd11-K250R ⌬sui2 strains does not re-imply that enhancing the rate of nucleotide dissociation quire additional copies of IMT, despite the fact that from eIF2 is an essential function of eIF2B. these conditions would be expected to bypass the eIF2B
Kinzy and Woolford (1995) demonstrated pre- nucleotide exchange function. In fact,
co-overexpres-viously that additional copies of theTEF2gene, encod- sion ofgcd11-K250R,SUI3, andIMTis sufficient to by-ing elongation factor eEF1A (previously eEF1␣), were pass eIF2Bin vivo(J. NikaandE. M. Hannig, unpub-sufficient to bypass the requirement for its exchange lished observations). Furthermore, we demonstrated factor, eEF1B, when provided on a low-copy-number recently that eIF2␣is required to promote efficient in-plasmid. The requirement for elevated levels of both
teraction between eIF2 and eIF2Bin vitro(Nikaet al.
wild-type eIF2 and initiator tRNA in bypassing eIF2B
2001). These combined observations suggest that eIF2B function suggests that eIF2 and/or initiator tRNA are
may contribute to the formation of eIF2␥K250R ternary normally maintained at limiting levels such that eIF2B
complexes in a manner that does not appear to require is essential for promoting levels of ternary complex
re-catalyzed nucleotide exchange and that is independent quired in rapidly growing cells. Such a mechanism
of (or less dependent upon) direct eIF2/eIF2B interac-would also allow for rapid and effective changes in the
tion. An alternative means through which eIF2B may level of ternary complexes by modulating eIF2B activity
facilitate ternary complex formation is by increasing in response to various stimuli and, as such, may play an
local concentrations of tRNA, perhaps through a chan-important role in regulating cell growth. Our results
neling type of mechanism. Such a mechanism may be make the prediction that cells harboring mutations
anal-direct or inanal-direct, would not require anal-direct interaction ogous togcd11-K250R may be less sensitive to growth
between eIF2 and eIF2B, and may be facilitated by a regulation mediated through protein kinases that
phos-ribosomal localization of at least a portion of the eIF2B phorylate the ␣-subunit of eIF2 (Samuel 1993; Wek
pool (Mattset al.1988;Ciganet al.1991;Chakrabarti
1994;Shiet al. 1998;Sood et al. 2000a,b). Additional
andMaitra1992; Ramaiahet al.1992;Bushmanet al.
strategies developed to circumvent this regulatory
mech-1993b; Muelleret al.1998). However, we cannot rule anism, such as specific alterations in eIF2␣that render
out completely the involvement of at least some form it refractory to phosphorylation (Donzeet al.1995) or
of an eIF2·eIF2B intermediate. In this respect, it is inter-dominant negative mutations in PKR (Koromilaset al.
esting to note recent data demonstrating genetic as well 1992; Meurs et al. 1993; Barber et al. 1995a), have
as physical interaction betweenGCD11andLOS1(
Hell-been shown to promote tumor formation in mammals,
muthet al.1998;Grosshanset al.2000).LOS1encodes implying that PKR may function as a tumor suppressor
a member of the-importin family that plays a
nonessen-(Lengyel1993).
tial role in transport of tRNA across the yeast nuclear Conditions required to suppress a ⌬sui2 mutation
membrane. However, it is unclear whether this interac-differed somewhat from those required to suppress the
tion is important in the specific transport and/or local-deletion of essential eIF2B subunit genes. In the latter
ization of initiator tRNA. On the other hand, eEF1A case, increased IMT gene dosage was absolutely
re-(the functional homolog of prokaryotic EF1A) does ap-quired, whereas suppression of ⌬sui2 in gcd11-K250R
pear to be required for efficient nuclear export of cer-strains was more efficient in the presence of, but did
func-Bourne, H. R., D. A. SandersandF. McCormick,1991 The GTPase tion in coordinating translation and tRNA export in
superfamily: conserved structure and molecular mechanism.
Na-yeast (Grosshanset al.2000). ture349:117–127.
Our combined data predict that eIF2␥ carries out Brostrom, C. O., C. R. Prostko, R. J. KaufmanandM. A. Brostrom, 1996 Inhibition of translational initiation by activators of the all eIF2 functions required for translation initiation,
glucose-regulated stress protein and heat shock protein stress including interactions with ribosomes and other transla- response systems. Role of the interferon-inducible double-tional factors, start site recognition, nucleotide ex- stranded RNA-activated eukaryotic initiation factor 2␣kinase. J.
Biol. Chem.271:24995–25002. change (in the presence or absence of eIF2B), and
Bushman, J. L., A. I. Asuru, R. L. MattsandA. G. Hinnebusch,
formation of ternary complexes. This suggests a model 1993a Evidence thatGCD6andGCD7, translational regulators in which eIF2␥ comprises the eIF2 functional core, ofGCN4, are subunits of the guanine nucleotide exchange factor for eIF-2 inSaccharomyces cerevisiae.Mol. Cell. Biol.13:1920–1932. whereas the␣-subunit of eIF2 and the eIF2B
heteropen-Bushman, J. L., M. Foiani, A. M. Cigan, C. J. PaddonandA. G.
tamer form a regulatory core that modulates the level Hinnebusch,1993b Guanine nucleotide exchange factor for of eIF2 function by regulating nucleotide exchange and eukaryotic translation initiation factor 2 inSaccharomyces cerevisiae: interactions between the essential subunits GCD2, GCD6, and formation of ternary complexesin vivo.The availability
GCD7 and the regulatory subunit GCN3. Mol. Cell. Biol.13: of yeast strains lacking normally essential subunits of
4618–4631.
eIF2 and eIF2B should provide valuable tools for dis- Chakrabarti, A.,andU. Maitra,1992 Release and recycling of eukaryotic initiation factor 2 in the formation of an 80 S ribosomal secting the functions of individual polypeptides in these
polypeptide chain initiation complex. J. Biol. Chem.267:12964– multisubunit complexes. Such functions could include
12972.
roles in catalysis, as well as regulatory functions involved Chong, K. L., L. Feng, K. Schappert, E. Meurs, T. F. Donahueet al., 1992 Human p68 kinase exhibits growth suppression in in cellular responses to stress or other environmental
yeast and homology to the translational regulator GCN2. EMBO stimuli (WelshandProud1992;Engelberget al.1994;
J.11:1553–1562.
Kimball and Jefferson 1994; Brostrom et al. 1996; Cigan, A. M., E. K. Pabich, L. FengandT. F. Donahue,1989 Yeast translation initiation suppressorsui2encodes the␣subunit of
Gallieet al.1997;Quet al.1997;Scheperet al.1997).
eukaryotic initiation factor 2 and shares sequence identity with In this regard, results of previous studies have indicated
the human␣subunit. Proc. Natl. Acad. Sci. USA86:2784–2788. a role for Gcd6p in catalysis, whereas Gcd2p, Gcd7p, Cigan, A. M., M. Foiani, E. M. Hannig andA. G. Hinnebusch, and Gcn3p appear to form a regulatory subcomplex 1991 Complex formation by positive and negative translational
regulators ofGCN4.Mol. Cell. Biol.11:3217–3228.
(YangandHinnebusch1996;Fabianet al.1997;
Pav-Cigan, A. M., J. L. Bushman, T. R. BoalandA. G. Hinnebusch,1993
itt et al. 1997, 1998; Gomez and Pavitt 2000). Bio- A protein complex of translational regulators ofGCN4mRNA is chemical analysis of individual eIF2B polypeptides and the guanine nucleotide-exchange factor for translation initiation
factor 2 in yeast. Proc. Natl. Acad. Sci. USA90:5350–5354. subcomplexes that are devoid of contaminating
sub-Dever, T. E., M. J. GlyniasandW. C. Merrick,1987 GTP-binding units, purified from strains using the genetic back- domain: three consensus sequence elements with distinct spac-grounds developed in this article, in addition to further ing. Proc. Natl. Acad. Sci. USA84:1814–1818.
Dever, T. E., L. Feng, R. C. Wek, A. M. Cigan, T. F. Donahueet al., genetic analyses of these strains, will allow these
ques-1992 Phosphorylation of initiation factor 2␣by protein kinase tions to be addressed more directly. GCN2 mediates gene-specific translational control ofGCN4in
yeast. Cell68:585–596. We thank Juan Gonzalez and Larry Reitzer for thoughtful comments
Dever, T. E., J. J. Chen, G. N. Barber, A. M. Cigan, L. Fenget al., on the manuscript, Alan Hinnebusch forGCD6/GCD7plasmids and
1993 Mammalian eukaryotic initiation factor 2␣kinases func-Gcd6p/Gcd7p antibodies, Tom Dever for PKR plasmid constructs,
tionally substitute for GCN2 protein kinase in theGCN4 transla-and Weimin Yang for construction of the yeast strain used for
purifica-tional control mechanism of yeast. Proc. Natl. Acad. Sci. USA90:
tion of eIF2B. This work was supported by a grant from the American 4616–4620.
Cancer Society (RPG-97-061-01-NP). Dever, T. E., W. Yang, S. Astrom, A. S. BystromandA. G.
Hinne-busch,1995 Modulation of tRNA(iMet), eIF-2, and eIF-2B ex-pression shows thatGCN4translation is inversely coupled to the level of eIF-2.GTP.Met- tRNA(iMet) ternary complexes. Mol. Cell. LITERATURE CITED Biol.15:6351–6363.
Donahue, T. F., A. M. Cigan, E. K. PabichandB. C. Valavicius, Ahmad, M. F., N. Nasrin, M. K. Bagchi, I. ChakravartyandN. K.
1988 Mutations at a Zn(II) finger motif in the yeast eIF-2gene Gupta,1985 A comparative study of the characteristics of eIF-2
alter ribosomal start-site selection during the scanning process. and eIF-2-ancillary factor activities from yeastSaccharomyces
cerevis-Cell54:621–632.
iaeand rabbit reticulocytes. J. Biol. Chem.260:6960–6965.
Donze, O., R. Jagus, A. E. Koromilas, J. W. HersheyandN. Sonen-Alani, E., L. CaoandN. Kleckner,1987 A method for gene
disrup-berg, 1995 Abrogation of translation initiation factor eIF-2 tion that allows repeated use ofURA3selection in the
construc-phosphorylation causes malignant transformation of NIH 3T3 tion of multiply disrupted yeast strains. Genetics116:541–545.
cells. EMBO J.14:3828–3834. Barber, G. N., R. Jagus, E. F. Meurs, A. G. HovanessianandM. G.
Dorris, D. R., F. L. EricksonandE. M. Hannig,1995 Mutations in Katze,1995a Molecular mechanisms responsible for malignant
GCD11, the structural gene for eIF-2␥in yeast, alter translational transformation by regulatory and catalytic domain variants of the
regulation ofGCN4and the selection of the start site for protein interferon-induced enzyme RNA-dependent protein kinase. J.
synthesis. EMBO J.14:2239–2249. Biol. Chem.270:17423–17428.
Engelberg, D., C. Klein, H. Martinetto, K. StruhlandM. Karin, Barber, G. N., M. Wambach, S. Thompson, R. Jagus andM. G.
1994 The UV response involving the Ras signaling pathway and Katze,1995b Mutants of the RNA-dependent protein kinase
AP-1 transcription factors is conserved between yeast and mam-(PKR) lacking double-stranded RNA binding domain I can act as
mals. Cell77:381–390. transdominant inhibitors and induce malignant transformation.
Erickson, F. L.,andE. M. Hannig,1996 Ligand interactions with Mol. Cell. Biol.15:3138–3146.
eukaryotic translation initiation factor 2: role of the␥-subunit. Boeke, J. D., J. Trueheart, G. NatsoulisandG. R. Fink,1987
5-EMBO J.15:6311–6320. Fluoroorotic acid as a selective agent in yeast molecular genetics.
1997 Functional analysis of homologs of translation initiation the lethality of aTEF5null allele encoding elongation factor 1. factor 2␥in yeast. Mol. Gen. Genet.253:711–719. Genetics141:481–489.
Ernst, H., R. F. DuncanandJ. W. Hershey,1987 Cloning and Koonin, E. V.,1995 Multidomain organization of eukaryotic gua-sequencing of complementary DNAs encoding the␣-subunit of nine nucleotide exchange translation initiation factor eIF-2B sub-translational initiation factor eIF-2. Characterization of the pro- units revealed by analysis of conserved sequence motifs. Protein tein and its messenger RNA. J. Biol. Chem.262:1206–1212. Sci.4:1608–1617.
Fabian, J. R., S. R. Kimball, N. K. HeinzingerandL. S. Jefferson, Koromilas, A. E., S. Roy, G. N. Barber, M. G. KatzeandN. Sonen-1997 Subunit assembly and guanine nucleotide exchange activ- berg,1992 Malignant transformation by a mutant of the IFN-ity of eukaryotic initiation factor-2B expressed in Sf 9 cells. J. Biol. inducible dsRNA-dependent protein kinase. Science257:1685–
Chem.272:12359–12365. 1689.
Gallie, D. R., H. Le, C. Caldwell, R. L. Tanguay, N. X. Hoang Lengyel, P.,1993 Tumor-suppressor genes: news about the
inter-et al., 1997 The phosphorylation state of translation initiation feron connection. Proc. Natl. Acad. Sci. USA90:5893–5895. factors is regulated developmentally and following heat shock in Manchester, K. L.,andP. Stasikowski,1990 Kinetic parameters wheat. J. Biol. Chem.272:1046–1053. governing the formation of eIF-2.methionyl-tRNAi complexes in Gaspar, N. J., T. G. Kinzy, B. J. Scherer, M. Humbelin, J. W. Hershey protein synthesis. Biochem. Int.20:257–265.
et al., 1994 Translation initiation factor eIF-2. Cloning and ex- Marton, M. J., D. CrouchandA. G. Hinnebusch,1993 GCN1, a pression of the human cDNA encoding the ␥-subunit. J. Biol. translational activator ofGCN4inSaccharomyces cerevisiae, is re-Chem.269:3415–3422. quired for phosphorylation of eukaryotic translation initiation Gietz, R. D.,andA. Sugino,1988 New yeast-Escherichia coli shuttle factor 2 by protein kinase GCN2. Mol. Cell. Biol.13:3541–3556.
vectors constructed with in vitro mutagenized yeast genes lacking Matts, R. L., N. S. Thomas, R. HurstandI. M. London,1988 Cor-six-base pair restriction sites. Gene74:527–534. relation between the distribution of the reversing factor and Gomez, E.,andG. D. Pavitt,2000 Identification of domains and eukaryotic initiation factor 2 in heme-deficient or double-residues within theε-subunit of eukaryotic translation initiation stranded RNA-inhibited reticulocyte lysates. FEBS Lett.236:179– factor 2B (eIF2Bepsilon) required for guanine nucleotide ex- 184.
change reveals a novel activation function promoted by eIF2B Merrick, W. C.,1992 Mechanism and regulation of eukaryotic pro-complex formation. Mol. Cell. Biol.20:3965–3976. tein synthesis. Microbiol. Rev.56:291–315.
Grosshans, H., E. HurtandG. Simos,2000 An aminoacylation- Meurs, E. F., J. Galabru, G. N. Barber, M. G. KatzeandA. G. dependent nuclear tRNA export pathway in yeast. Genes Dev. Hovanessian, 1993 Tumor suppressor function of the
inter-14:830–840. feron-induced double-stranded RNA-activated protein kinase.
Hannig, E. M.,andA. G. Hinnebusch,1988 Molecular analysis of Proc. Natl. Acad. Sci. USA90:232–236.
GCN3, a translational activator ofGCN4: evidence for posttransla- Mueller, P. P., P. Grueter, A. G. HinnebuschandH. Trachsel, tional control of GCN3regulatory function. Mol. Cell. Biol.8: 1998 A ribosomal protein is required for translational
regula-4808–4820. tion ofGCN4mRNA. Evidence for involvement of the ribosome
Hannig, E. M., N. P. Williams, R. C. WekandA. G. Hinnebusch, in eIF2 recycling. J. Biol. Chem.273:32870–32877.
1990 The translational activator GCN3functions downstream Nika, J., S. RippelandE. M. Hannig,2001 Biochemical analysis of fromGCN1andGCN2in the regulatory pathway that couples the eIF2␥complex reveals a structural function for eIF2␣in
GCN4expression to amino acid availability inSaccharomyces cerevis- catalyzed nucleotide exchange. J. Biol. Chem.276:1051–1056. iae.Genetics126:549–562. Paddon, C. J., E. M. HannigandA. G. Hinnebusch,1989 Amino Hannig, E. M., A. M. Cigan, B. A. FreemanandT. G. Kinzy,1993 acid sequence similarity between GCN3 and GCD2, positive and
GCD11, a negative regulator ofGCN4expression, encodes the␥ negative translational regulators ofGCN4: evidence for antago-subunit of eIF-2 in Saccharomyces cerevisiae.Mol. Cell. Biol. 13: nism by competition. Genetics122:551–559.
506–520.
Pain, V. M.,1996 Initiation of protein synthesis in eukaryotic cells. Hellmuth, K., D. M. Lau, F. R. Bischoff, M. Kunzler, E. C. Hurt Eur. J. Biochem.236:747–771.
et al., 1998 Yeast Los1p has properties of an exportin-like
nucleo-Pathak, V. K., P. J. Nielsen, H. TrachselandJ. W. Hershey,1988 cytoplasmic transport factor for tRNA. Mol. Cell. Biol.18:6374– Structure of thesubunit of translational initiation factor eIF-2. 6386.
Cell54:633–639. Hershey, J. W., 1991 Translational control in mammalian cells.
Pavitt, G. D., W. YangandA. G. Hinnebusch,1997 Homologous Annu. Rev. Biochem.60:717–755.
segments in three subunits of the guanine nucleotide exchange Hill, D. E.,andK. Struhl, 1988 Molecular characterization of
factor eIF2B mediate translational regulation by phosphorylation
GCD1, a yeast gene required for general control of amino acid
of eIF2. Mol. Cell. Biol.17:1298–1313. biosynthesis and cell-cycle initiation. Nucleic Acids Res.16:9253–
Pavitt, G. D., K. V. Ramaiah, S. R. KimballandA. G. Hinnebusch, 9265.
1998 eIF2 independently binds two distinct eIF2B subcom-Hinnebusch, A. G.,1993 Gene-specific translational control of the
plexes that catalyze and regulate guanine-nucleotide exchange. yeastGCN4gene by phosphorylation of eukaryotic initiation
fac-Genes Dev.12:514–526. tor 2. Mol. Microbiol.10:215–223.
Price, N.,andC. Proud,1994 The guanine nucleotide-exchange Hinnebusch, A. G.,1994 Translational control ofGCN4: an in vivo
factor, eIF-2B. Biochimie76:748–760. barometer of initiation-factor activity. Trends Biochem. Sci.19:
Price, N. T., S. R. Kimball, L. S. JeffersonandC. G. Proud,1996a 409–414.
Cloning of cDNA for the␥-subunit of mammalian translation Hinnebusch, A. G.,1997 Translational regulation of yeastGCN4.
initiation factor 2B, the guanine nucleotide-exchange factor for A window on factors that control initiator-TRNA binding to the
eukaryotic initiation factor 2. Biochem. J.318:631–636. ribosome. J. Biol. Chem.272:21661–21664.
Price, N. T., H. Mellor, B. L. Craddock, K. M. Flowers, S. R. Hinnebusch, A. G.,andS. W. Liebman,1991 Protein synthesis and
Kimballet al., 1996b eIF2B, the guanine nucleotide-exchange translational control in Saccharomyces cerevisiae, pp. 627–735 in
factor for eukaryotic initiation factor 2. Sequence conservation
The Molecular Biology of the Yeast Saccharomyces: Genome Dynamics,
between the␣,and␦subunits of eIF2B from mammals and
Protein Synthesis, and Energetics, edited byJ. R. Broach, J. R.
Prin-yeast. Biochem. J.318:637–643. gleandE. W. Jones.Cold Spring Harbor Laboratory Press, Cold
Qu, S., S. E. Perlaky, E. L. Organ, D. CrawfordandD. R. Cavener, Spring Harbor, NY.
1997 Mutations at the Ser50 residue of translation factor eIF-Kimball, S. R.,andL. S. Jefferson,1994 Mechanisms of
transla-2␣dominantly affect developmental rate, body weight, and viabil-tional control in liver and skeletal muscle. Biochimie76:729–736.
ity ofDrosophila melanogaster.Gene Exp.6:349–360. Kimball, S. R., J. R. Fabian, G. D. Pavitt, A. G. HinnebuschandL. S.
Ramaiah, K. V., R. S. Dhindsa, J. J. Chen, I. M. LondonandD. Jefferson, 1998 Regulation of guanine nucleotide exchange
Levin,1992 Recycling and phosphorylation of eukaryotic initia-through phosphorylation of eukaryotic initiation factor eIF2␣.
tion factor 2 on 60S subunits of 80S initiation complexes and Role of the ␣- and ␦- subunits of eIF2B. J. Biol. Chem. 273:
polysomes. Proc. Natl. Acad. Sci. USA89:12063–12067. 12841–12845.
Rhoads, R. E.,1993 Regulation of eukaryotic protein synthesis by Kinzy, T. G.,andJ. L. Woolford, Jr.,1995 Increased expression
Rose, M. D.,andJ. R. Broach,1991 Cloning genes by complementa- homologues in humans,Drosophila melanogasterandCaenorhabditis elegansthat mediate translational control in response to endoplas-tion in yeast. Methods Enzymol.194:195–229.
Rowlands, A. G., R. PanniersandE. C. Henshaw,1988 The cata- mic reticulum stress. Biochem. J.346 Pt. 2:281–293.
Sood, R., A. C. Porter, D. A. Olsen, D. R. Cavener andR. C. lytic mechanism of guanine nucleotide exchange factor action
and competitive inhibition by phosphorylated eukaryotic initia- Wek,2000b A mammalian homologue of GCN2 protein kinase important for translational control by phosphorylation of eukary-tion factor 2. J. Biol. Chem.263:5526–5533.
Sachs, A. B., P. SarnowandM. W. Hentze,1997 Starting at the otic initiation factor-2␣. Genetics154:787–801.
Vazquez de Aldana, C. R.,andA. G. Hinnebusch,1994 Mutations beginning, middle, and end: translation initiation in eukaryotes.
Cell89:831–838. in the GCD7 subunit of yeast guanine nucleotide exchange factor eIF-2B overcome the inhibitory effects of phosphorylated eIF-2 Samuel, C. E.,1993 The eIF-2␣protein kinases, regulators of
transla-tion in eukaryotes from yeasts to humans. J. Biol. Chem.268: on translation initiation. Mol. Cell. Biol.14:3208–3222. Vazquez de Aldana, C. R., T. E. DeverandA. G. Hinnebusch,1993 7603–7606.
Scheper, G. C., J. Mulder, M. Kleijn, H. O. Voorma, A. A. Thomas Mutations in the␣subunit of eukaryotic translation initiation factor 2 (eIF-2␣) that overcome the inhibitory effect of eIF-2␣
et al., 1997 Inactivation of eIF2B and phosphorylation of PHAS-I
in heat-shocked rat hepatoma cells. J. Biol. Chem.272:26850– phosphorylation on translation initiation. Proc. Natl. Acad. Sci. USA90:7215–7219.
26856.
Sherman, F., G. R. Fink andJ. B. Hicks, 1986 Methods in Yeast Wek, R. C., 1994 eIF-2 kinases: regulators of general and
gene-Genetics.Cold Spring Harbor Laboratory, Cold Spring Harbor, specific translation initiation. Trends Biochem. Sci.19:491–496. NY. Welsh, G. I.,andC. G. Proud,1992 Regulation of protein synthesis Shi, Y., K. M. Vattem, R. Sood, J. An, J. Lianget al., 1998 Identifica- in Swiss 3T3 fibroblasts. Rapid activation of the guanine-nucleo-tion and characterizaguanine-nucleo-tion of pancreatic eukaryotic initiaguanine-nucleo-tion fac- tide-exchange factor by insulin and growth factors. Biochem. J. tor 2␣-subunit kinase, PEK, involved in translational control. 284:19–23.
Mol. Cell. Biol.18:7499–7509. Yang, W.,andA. G. Hinnebusch,1996 Identification of a regulatory Sikorski, R. S.,andP. Hieter,1989 A system of shuttle vectors and subcomplex in the guanine nucleotide exchange factor eIF2B yeast host strains designed for efficient manipulation of DNA in that mediates inhibition by phosphorylated eIF2. Mol. Cell. Biol.
Saccharomyces cerevisiae.Genetics122:19–27. 16:6603–6616.
Sood, R., A. C. Porter, K. Ma, L. A. Quilliam andR. C. Wek,