1
CANCER: A TURBULENCE PROBLEM
Abicumaran Uthamacumaran Concordia University
7141 Sherbrooke St W, Montreal, Quebec H4B 1R6
ABSTRACT
As we transition towards an era of Computational Medicine, our mathematical models of cancer dynamics must be revised. As such, recent evidence supports the perspective that
cancer-microenvironment interactions consist of turbulent flows and strange attractor dynamics. Cancer pattern formation, invasion-growth dynamics, protein folding kinetics, stem cell fate bifurcations and metastatic invasion are discussed within the context of hydrodynamical turbulence. Cancer is presented as a three-dimensional Navier-Stokes equations global regularity, smoothness and existence problem.
POSTULATE: Cancer stem cells are strange attractors on the Waddington energy landscape governed by turbulence dynamics.
EMERGENCE
Cancers are complex systems. The Hanahan-Weinberg ‘hallmarks’ are an inadequate representation of these dynamical systems. Complex systems are systems in which the interactions of its elements and parts cannot be deduced due to interdependence. Some general characteristics of complex systems include nonlinearity, emergence, computational irreducibility, unpredictability,
nonequilibrium statistical mechanics and intractability. In simple terms, the concerted whole cannot be defined by the sum of its interacting parts. For example, the flow of exosomes between cancer cells are emergent characteristics of tumor ecosystems. It is impossible to understand exosomes without complex systems approaches. Exosomes are heterogeneous nano-scaled packets of information forming complex long-range communication networks. Along with ctDNA (circulating tumor DNA), exosomes are emerging targets for early tumor detection from liquid biopsies of patients (1, 2). Human embryonic stem cells-derived exosomes have shown capability of reprogramming malignant cancer phenotypes to benign-like fates, indicating exosomes are potent phenotype reprogramming machineries (3). However, the identity of cancer stem cells remains ambiguous. It is debated whether cancer stem cells conform a small subset of the tumor hierarchy or whether all cancer cells are potentially stem cells (i.e., have the potency of phenotypic plasticity and unlimited replication). In attempt to reconcile such problems, precision oncology is transitioning towards the use of artificial intelligence and machine learning algorithms in guiding clinical decision-making (4). Machine intelligence is emerging as a powerful tool in deciphering cancer ecosystems.
and Weinberg. Recently, neuron–glioma interactions were demonstrated to be highly complex, bidirectional networks. High-grade gliomas integrate into the electrical networks of healthy neurons wherein depolarizing potassium currents promote glioma progression (8). These few examples indicate the cancer- microenvironment is a dynamical system and cannot be reduced to specific properties. The hallmarks of cancer such as unlimited replication, immortality, growth invasion, etc. are merely some of the many emergent properties exhibited by the phenotypic plasticity of cancer stem cells. This
irreducible complexity further postulates the plausibility of nonlocal cross-talks between cancer stem cells and their niche.
Our current statistical inference methods in modelling cancer Waddington landscapes (network state-space) mainly comprise of graph theory, classical information theory, Bayesian networks, cluster analysis and machine learning algorithms (9-11). Due to the stochastic nature of molecules, the transitions between the attractors of the landscape are defined by random walks on a network (i.e., Brownian motion) (12). That is, the fluctuations around the epigenetic barriers of the landscapes are governed by diffusion equations (13-15). Gene regulatory networks are extensively modeled as Boolean networks where each network element computes a Boolean function based on the values of inputs to that element (16). The on/off gene promoters can be represented as binary occupancies of 0 or 1. The length of the attractors formed by the networks depend on the connectivity parameter K denoting phase-transitions in the network defined as:
𝐾
𝐶=
1 2𝑝𝑐(1−𝑝𝑐),
where 𝑝𝑐denotes the critical probability (17). A Boolean network can exhibit chaotic behavior when a critical value of the average number of connections of nodes 𝐾𝑐 is reached. In the unstable regime, the
Hamming distance between two initially close attractor states on average grows exponentially in time, a signature of chaos. Typically, values of 𝐾𝐶 > 2 can result in chaotic network dynamics which is often equated with unstable, aperiodic behaviors.
A stochastic model of gene express dynamics leads to diffusion-like differential equations known as the chemical Master equation that describe small fluctuations around a stable attractor of the system. The Master equation describes the probability transitions from one node (gene) to another within
topological directed graphs (i.e., a state vector). However, the Master equation generally becomes non-integrable and intractable once the number of chemical species in the system reaches more than three (18). Therefore, rather than solving the Master equation for the chemical kinetics, the Gillespie
algorithm, Monte Carlo method or the piecewise deterministic Markov processes (PDMP) are often used to simulate the network dynamics (19, 20).
It is often assumed the mRNA transcription and its protein synthesis-degradation dynamics settles to steady-state probability distributions. The Fokker-Planck equations are variants of the Liouville-Master equation used to characterize the dynamics of signaling molecules (21). Given a first order reaction rate k, let P be the probability that n proteins exist at time t. Then, the probability distribution of a single protein’s production/degradation kinetics is given by the Fokker-Planck equation:
𝜕𝑃𝑛
𝜕𝑡 = 𝑘[𝑃𝑛−1− 𝑃𝑛] − 𝑑[𝑛𝑃𝑛− (𝑛 + 1)𝑃𝑛+1].
accounted in current paradigms.
Consider a simple gene-pair Boolean circuit as follows. In vector form, the minimal set of equations representing this Boolean network is 𝑑𝑥𝑑𝑡= 𝐹(𝑥) = [𝐹1(𝑥1, 𝑥2), 𝐹2(𝑥1, 𝑥2)], where 𝑥1, 𝑥2 represent the cellular expression or activation levels of the two lineage-determining transcription factors, and 𝑎1,𝑎2, 𝑏1, 𝑏2, 𝑘1, 𝑘2 are positive parameters that denote the strength of the kinetics (binding/unbinding interactions). 𝐹1 and 𝐹2 are the driving forces of the system in a general 1D setting. Then,
𝑑𝑥1
𝑑𝑡 =
𝑎1𝑥1𝑛 𝑆𝑛+𝑥 1 𝑛+ 𝑏1𝑆𝑛
𝑆𝑛+𝑥 2
𝑛− 𝑘1𝑥1= 𝐹1(𝑥1, 𝑥2), and 𝑑𝑥2
𝑑𝑡 =
𝑎2𝑥2𝑛 𝑆𝑛+𝑥
2𝑛+ 𝑏2𝑆𝑛 𝑆𝑛+𝑥
1𝑛− 𝑘2𝑥2= 𝐹2(𝑥1, 𝑥2), where the first expression represents the self-activation (often assumed to be a sigmoidal transfer function of strength a) and the second term denotes the mutual inhibition given a basal expression of strength b. The third term is the first-order inactivation given in terms of the degradation rates k. The chemical kinetics of the Gene Regulatory Networks (GRN) are often assumed to be Hill equations, Michaelis-Menten kinetics or differential rate laws determined by experimental data. However, in practice, inferring kinetic equations is an NP-complete problem and distinguishing noise from data remains a challenge. The dynamics of a gene regulatory network in phase-space is known as an epigenetic (Waddington) landscape. In the resulting landscape, the vertical axis represents the potential U which is related to the probability density P as given by U = - ln P, while the horizontal axis represents the expression level of genes (14). The distribution of possible paths followed by a random walk on a Gene Regulatory Network (GRN) can be in three forms: finite, stretched exponential or power law (12). The latter two are used to describe the strange attractor dynamics of weather turbulence (i.e., Lorenz attractor) and protein folding (12, 23, 24). The Lorenz attractor, a strange attractor, is a toy model of deterministic chaos (i.e., sensitive dependence on initial conditions) (22). Strange attractors are also proposed as the solutions to the unsolved problem of fluid turbulence. The review paper intends to interlink the greatest unsolved problem in mathematical physics, fluid turbulence, to cancer dynamics.
The similarity between forecasting weather patterns and the reaction-diffusion dynamics of the immune systems have been documented (25). There is a striking similarity between fluid turbulence and pattern formation in biosystems. For example, turbulent flows of cytoskeletal fluids and fluids of Min proteins (MinC, MinD and MinE) from E. coli lysates forming cascades of patterns on a petri dish are suggesting the adoption of fluid dynamics in morphogenesis (26, 27). Tumorigenesis is merely a subset of such pattern formation systems. Current approaches to reconstructing tumor developmental landscapes hold the naïve view that cancer phenotypes are stable or fixed-point attractors on the network state space. However, experimental findings and mathematical modelling indicate cancer stem cells are strange
attractors.
By convention, the protein folding free-energy landscape is best defined as a stochastic process with spin glass formulation (28). However, the power law decay observed in protein folding kinetics is a signature of turbulence observed in experimental fluid dynamics (23, 29). Interestingly, many biological networks of protein-protein interactions and DNA-protein interactions form power law distributions (i.e., scale-free networks) (30). Furthermore, power law decay is a characteristic for many edges of chaos phenomena. The transition from quantum to classical chaos in dynamical systems forms a power-law decay as well (30). Power law decays may also help explain the extended quantum coherence and quantum transport observed in phenomena associated to the newly emerging field of quantum biology, such as the photosynthetic energy transfer of chromophores at room temperature (31).
thermal fluctuations. However, the emerging field of ‘quantum biology’ asserts quantum phase transitions can explain complex systems that are not fully defined by classical physics. There are numerous unorthodox interpretations of quantum mechanics such as Madelung’s hydrodynamics and de Broglie-Bohm pilot wave theory that both support macroscopic quantum effects and capture the principles of complex systems. These interpretations are also analogous to fluid dynamics and hence, related to the Navier-Stokes equations. Bohm’s concepts of wholeness, and the process of
enfoldment/unfoldment (i.e., implicate and explicate order) are embedded within complexity science (32). Furthermore, Bohm’s interpretation is driven by information; the active information (Quantum) potential guiding the trajectories of particles demonstrates nonlinearity, a doctrine of complex systems theory (32).
Many nanoscale and mesoscopic systems in soft matter physics are exhibiting macroscopic quantum effects as emergent phenomena. For example, enzymatic tunneling and the associated changes in protein conformations may involve quantum tunneling. With quantum tunneling, the transition-state energy barriers are bypassed (33). The quantum coherence of collective vibrational modes in proteins can withstand thermal fluctuations. Classical rate laws become inadequate descriptions of such chemical kinetics. Moreover, quantum effects such as proton tunneling between base pairs has been established as a promoter of mutations (34). Although fundamentally everything is quantum-mechanical, quantum biology asserts that macroscopic quantum effects such as nonlocality and quantum coherence for extended time scales exist in many-body (bio)systems indicating a scaling-problem alike the Navier-Stokes problem of turbulence. Moreover, whether criticality is observed in a system cannot be predicted, often until experimentally tested.
SYMMETRY-BREAKING
As mentioned, there is a striking similarity between fluid turbulence and chemical pattern formation in cellular systems. Ruelle described non-periodic cases of chemical oscillations exhibiting chaotic dynamics (29). Such turbulent chemical oscillations may be the problem underlying cancer pattern formation. Experimental turbulence exhibits three properties: (a) A continuous frequency spectrum, (b) Intermittency, i.e., the transition between laminar and chaotic flow regimes, and (c) Nonlinear resonances and nested multi-fractal structures that describe its complex bifurcations (29). All three properties can be accounted as ‘strange attractor’ dynamics (24, 29).
Turing in 1952 was the first to define morphogenesis using a set of nonlinear partial-differential
equations characterizing the activator-inhibitor dynamics of morphogen gradients (35). Tumorigenesis is a subset of morphogenesis exploiting this property to attain non-equilibrium patterning. Prigogine et al. proposed that Turing bifurcations from diffusion-driven instabilities in a nonequilibrium chemical system can result in the spontaneous emergence of dissipative structures (36). The general reaction- diffusion equation for tissue patterning is given by:
𝑐𝑡 = 𝐹(𝑐, 𝑥, 𝑦, 𝑧, 𝑡) + 𝐷∆𝑐,
where D is the diagonal matrix of diffusion coefficients, (𝑥, 𝑦, 𝑧) are the three-dimensional Cartesian coordinates of space, t is time, F determines the local reactions of chemical species and c is the concentration density. The Turing pattern formation for two chemical species (u and v) is given by:
𝜕𝑢
𝜕𝑡 = 𝐹(𝑢, 𝑣) − 𝑑𝑢 + 𝐷𝑢∆𝑢 𝜕𝑣
The equations state that the rate of change of u and v is the result of Production - (degradation+ diffusion) terms (i.e., F and G describe the production, d determines degradation, and D denotes the diffusion coefficients). The diffusion coefficient is assumed in most biological contexts to be given by the Einstein-Stokes relation for a spherical Brownian particle with Langevin drag forces: 𝐷 = 𝜇𝑘𝐵𝑇, where 𝜇 is the mobility, 𝑘𝐵 is the Boltzmann constant and T is the absolute temperature.
Each tumor is analogous to an ecosystem governed by predator-prey dynamics of the different cell types. The Lotka-Volterra equations are also well-known to produce chaotic behavior reminiscent of fluid turbulence (37, 38). The ecological predator-prey dynamics well-fit the power law decays observed in experimental fluid dynamics and Kolmogorov’s theory of turbulence (39). The Lotka-Volterra predator-prey dynamics are simple 1st order differential equations describing population density changes in time according to their growth rates given by:
𝑑𝑥
𝑑𝑡 = 𝑎𝑥 − 𝛽𝑥𝑦 𝑑𝑦
𝑑𝑡 = 𝛿𝑥𝑦 − 𝛾𝑦
(x and y are coordinates of two species, t= time and the remainders are growth/decay parameters).
Smale in 1976 showed that when the Lotka-Volterra model is applied to model the concentrations of N≥ 5 chemical species at certain conditions, asymptotic behaviour occurs as seen in experimental fluid dynamics (40). As such, complex Hopf-Turing bifurcations in a many-body chemical system form strange
attractors. Cancer being a complex, heterogeneous niche of dynamical phenotypes, the applicability of
strange attractors must be self- evident.
A simple reaction-diffusion model that incorporates the competitive predator-prey dynamics and period-doubling bifurcations is the Fisher-Kolmogorov-Petrovsky-Piskunov (FKPP) equation. It was initially used to describe the spatial spread of an advantageous allele in terms of its nonlinear travelling wave solutions (41, 42). The FKPP equation for c density of chemical species is given by:
𝜕𝑐
𝜕𝑡+ ∇ ∙ (𝑉𝑐) = 𝛾𝑐 (1 − 𝑐
𝐾) + ∇ ∙ (𝑢∇𝑁)
𝛾
is the growth rate of the population, K denotes the carrying capacity, V denotes a spatial parameter, t is time, u is the velocity field, and ∇ is the gradient. The NSE exhibit very complex, multi-nested bifurcations as well, where the competition is between the nonlinear velocity and viscosity terms.Current mathematical models assume the chemical oscillations of such morphogens behave as quasi-periodic oscillators. However, genetic oscillators can exhibit nonequilibrium behaviors when sudden outbursts of gene expression are encountered (i.e., intermittency). In support of this argument, using the finite element method, Halatek and Frey showed that the mass-conserved reaction-diffusion system of an E. coli pattern formation complex - the Min system - must undergo an initial phase of turbulent flows characterized by a Kolmogorov-Richardson-like energy cascade (43-45). Such dynamics
as cell division and morphogenesis. Simulations of the Min system showed that at the critical transition time (bifurcation), the propagating wavefront triggers an energy cascade of destabilized local equilibria entering a turbulent state whereby all state dynamics of the kymogram are spatially uncorrelated (45). The Min protein complexes are key regulators of cell polarity and asymmetric stem cell division. The mammalian orthologue of these proteins is known as PAR complexes. The PAR 3/6-aPKC complexes are determinants of stem-cell fate choice. They interplay the self-renewal and cleavage of cancer stem cells through feedback loops between the mitotic spindle network and cytoskeletal-ECM (Extracellular Matrix) remodeling pathways (46, 47). Further investigations on what other pathways pertaining to the cancer stem cell niche exhibit turbulent chemical oscillations, especially at the onset of pattern
formation and cell division are required to understand cancer stemness (48-50). It may be self-evident to postulate that a greater level of turbulent flows must occur in the chemical oscillations of stem cell symmetry-breaking protein complexes such as developmental morphogens, PAR proteins and cytoskeletal proteins (e.g. microtubules, actin, etc.) accounting for the chaotic patterning of cancer tissues.
Furthermore, it was recently confirmed the chaotic dynamics in the transcription factor NF-κB, a well-described transcriptional regulator in cancer networks, can affect downstream protein production (51). The nonlinear genetic oscillator NF-κB, when periodically driven by sufficiently large TNF (tumor necrosis factor) amplitudes, will exhibit deterministic chaos. That is, small initial changes in a cell (e.g., a mutation of a cancer driving gene) can lead to huge unexpected downstream cascades (i.e., butterfly effect). As another example, RNA-Seq data of the PI3K-mTOR axis was used as a toy-model to exhibit chaotic dynamics in breast cancer cells (52). Such findings contrast the widely held stochastic gene expression model with that of deterministic chaos.
On an additional note, cytokinesis defects form aneuploidy and genomic instability in cancers. The tetraploid cells generated through cytokinesis defects serve as intermediate states on the route to genetic diversification and heterogeneity (53). Furthermore, a Tubulin Code hypothesis is proposed to explain the computational complexity of cancer cell division (54). Surprisingly, cancer cells show upregulated MAPs (microtubule associated proteins) and an absence of PTMs (post-translational modifications) on HeLa cancer cells (54). The implications of these findings in the phenotypic plasticity and immortalized (in vitro) regenerative potential of HeLa cells remain unanswered.
FRACTALS AND CHAOS
In this section, tumors are painted as chaotic systems forming multifractal structures. As Prigogine states, chaos is a form of order in which the temporal sequence is complex (36). Chaos and multifractality are also signatures of fluid turbulence. Metastatic invasion and focal adhesion
dynamics in cancer cells are tightly regulated by fluid dynamics (55, 56). Non-laminar shear stress and turbulence may increase the adhesive ability of cancer cells in metastatic invasion through cross-talk between actin-cytoskeletal reorganization and polarity complexes. For example, higher streams of vascular flow induce aggressive phenotypes in ovarian cancer models by promoting EMT (epithelial-mesenchymal transition) (57, 58).
stage, and response to therapy (61). Numerical results show anomalous fluctuations in the fractal dimension at interfaces of tumor-host networks (62). The increased fractal characteristics of chromatin structure has been proposed as a diagnostic indicator of tumor self-organization (63). Moreover, multi-fractality is a characteristic signature of fluid turbulence (64). The notion of higher fractal dimensions is intertwined to increased turbulent flows. It is well-established that increased turbulent fluid circulation is an indicator of pathological conditions at systemic scales (65, 66).
Using a mass conservation equation and focal adhesion energy, recent 3D-modelling of glioblastoma shows vascular-endothelial interactions result in tumor relapse and chemoresistance. A simplified Navier-Stokes equation (i.e., Darcy’s law) was used to model GBM (Glioblastoma Multiforme)
patterning, assuming a mass-averaged velocity of solid components (tumor cells) and a focal adhesion energy modelled as a double-well potential (67). Darcy’s law can be derived from the 3D Navier-Stokes equations for a shallow flow between two plates. Electric cell impedance recordings were performed in rat’s prostate cancers. The time-series Fourier analysis of cancer micro-motions were assessed by Takens’ theorem (time-delay embedding) to detect patterns distinguishable from a random signal. The attractor reconstruction showed positive Lyapunov exponents in phase portraits (i.e., a signature of chaos) (68). Moreover, Posadas et al. demonstrated the Verhulst logistic map is a simple chaotic oscillator that encapsulates the period-doubling bifurcations observed in the cancer cells (68). Strange attractor solutions resembling the Lorenz attractor emerged in the reaction-diffusion invasion model of melanoma with oxygen-nutrients dependence as a critical order parameter.
A chaotic tumor growth model was shown by Itik and Banks adapted from an earlier model proposed by de Pillis and Radunskaya, describing the competitive interactions among three cell populations: host cells, effector immune cells, and tumor cells (69). It only differed from the earlier model by a non-constant influx of effector immune cells. Chaotic attractors emerged in the reaction-diffusion modelling of tumor growth depending on changes in the control bifurcation parameters, which vary amidst different phenotypes of a heterogeneous ecosystem (e.g. oxygen concentration, glucose level, tumor volume, diffusion from surface and growth parameters) (70). Lyapunov exponents and fractal dimension were calculated for the tumor patterning. While the Lorenz attractor has a fractal dimension of ≈2.06 (assessed by the Box-counting algorithm), the cancer models showed a fractal dimension of ≈2.03 indicating a chaotic attractor with Shilnikov-like bifurcations (69, 70). Similar studies by Lettelier et al. demonstrated the solutions to the competitive cancer growth model are topologically equivalent to the spiral Rössler attractor, a strange attractor (71). Moreover, tumor growth rates are assumed to fit logistic equations or Gompertz functions, as deemed appropriate with experimental fitting. With a set of parameters defining the growth and death rates of each cell type chaotic attractors emerged along with fixed points (stable equilibria) in the bifurcation analysis (71). The tumor-immune competitive networks described by a coupled set of three ordinary differential equations showed period-doubling bifurcations on the route to chaos with time-delay analysis, wherein strange attractor solutions emerged as
indicators of tumor relapse. The results were recently reconfirmed for a cancer model with time-delay as the bifurcation parameter. It exhibited periodic oscillations as well as chaotic behavior (strange attractors), which are indicators of long-term tumor relapse (72). These findings are some of the many emerging data suggesting cancer cells are aberrant, unsteady attractors in the network state-space governed by nonlinear dynamics (73).
TURBULENCE
solutions to turbulence is well-established (75). By convention, cells are considered highly viscous structures with macromolecular crowding and hence the motion of proteins is governed by diffusion equations. However, reaction-diffusion systems can transit to intermittent puffs of turbulent flows (i.e., chemical turbulence). Microscopic fluctuations and sudden outbursts in turbulence denote
intermittency, a statistical property observed in the chemical oscillations of pattern-formation systems (29). If one takes the concentrations of a chemical system, as in the case of gene expression activity, to be defined as the state vector X defined in time-series 𝑋1(𝑡), 𝑋2(𝑡), … , 𝑋𝑚(𝑡) at time t, the gene expression dynamics can be mapped in similarity to the time evolution of a viscous fluid by the NSE equations: 𝑑𝑡𝑑 𝑋(𝑡) = 𝑋𝜇(𝑡), where 𝜇 is the Reynolds parameter or diffusion coefficient (44).
There are asymptotic measures to characterize the time evolution of complex fluid flows from the average statistical properties of turbulent eddies and vortices. The frequency spectra analysis from the square-amplitude of the frequency modes of the fluid motion can detect strange attractor presence, when broad bands are observed that are not regularly spaced. This is analogous to the approach used by Posadas et al. to detect strange attractors in tumor cell cultures (68). Our current understanding of complex fluid patterns such as turbulence are derived from studying the statistical properties of experimental fluid dynamics, such as the power law decays in Kolmogorov’s model. All data of experimental fluid turbulence show that the solutions of the NSE tend asymptotically to a set with complex structures and multi-fractal character, namely, strange attractors (24, 29). Topological graph theory, the current framework of reconstructing gene regulatory networks, is also emerging as a powerful tool to investigate the spatial structure of turbulent flows as complex network structures (76, 77).
As discussed, recent data suggests the roles of chemical turbulence in the emergence of ordered structures during cellular pattern formation and protein folding. Chemical turbulence at the onset of protein-mediated cell patterning was recently confirmed experimentally (27). Following turbulence transition, coherent patterns emerged through diffusively coupled local equilibria. The chemical
turbulences were shown to occur in very rapid bursts followed by laminar-like flows (i.e., intermittency). Cell division is orchestrated by intracellular protein patterning, mainly from EMT
(epithelial-mesenchymal transition) pathways, cytoskeletal filaments and cell polarity complexes. Abnormal cell division is the primary signature of cancer. The recent theory by Halatek and Frey challenged the dogma (45). Using the finite elements method (FEM), a computational algorithm used to approximate complex fluid flows, simulations predicted chemical turbulence at the onset of the pattern-forming instabilities within cells. Chemical turbulence was qualitatively used to characterize the spatio-temporal chaos observed at the onset of pattern formation. Cytosolic diffusion constants 𝐷𝑐were in the order of 60 𝜇𝑚2𝑠−1, where the MinD-ATP/ADP in bulk were given by the Turing reaction-diffusion equations:
𝜕𝑡𝑢𝐷(𝑧, 𝑡) = 𝐷𝑐∇𝑧2𝑢𝐷− 𝐹𝑢𝐷, 𝜕𝑡𝑢𝑇(𝑧, 𝑡) = 𝐷𝑐∇𝑧2𝑢𝑇+ 𝐹𝑢𝐷,
With
𝐹
𝑢𝐷denoting the local-reaction term, and 𝑢𝐷 and 𝑢𝑇 defining the cytosolic densities of MinD-ATP/ADP conformations; the MinD-ATP was assumed to bind to the membrane via nonlinear coupling rate constants. The resultant kymograms demonstrated turbulent flow patterns at low MinE/MinD
ratios. As mentioned, low levels of gene expressions or protein fluctuations are filtered out in the single
The PAR cell polarity complex is the mammalian equivalence of the Min proteins complex which coordinate cancer cell division. According to these findings, chemical turbulence in pattern formation is used as a synonym for spatio-temporal chaos, i.e., a broad distribution in the power spectrum and a low spatial correlation length reminiscent of the Kolmogorov spectrum. The term was adapted from the work by Nobel laureate Gerhard Ertl on reactions of heterogeneous catalysis (79). According to this work, during chemical turbulence, both the amplitude and the phase of local concentration oscillations are strongly fluctuating, creating spiral waves as seen in the findings of Denk et al. concerning Min proteins pattern formation (27). Turbulent chemical oscillations can give rise to both patchy multi-fractal structures and ordered patterns in cellular reaction-diffusion systems as seen in the multi-fractal self-organization of cancer cells (80, 81).
Ito et al. experimentally demonstrated turbulent flows in the environment can increase the level of iPSC production in hematopoietic stem cells (82). Turbulent flow generating turbines and microPIV (particle imaging velocimetry) techniques were used to increase the production of platelet-forming stem cells (82). As such, we can further speculate that cancer stem cells may be exploiting turbulent flows in their surroundings through the various mechanisms discussed, such as angiogenesis/nutrient supply, pattern formation and metastasis, to confer their microenvironment-specific adaptive
advantages, especially in the context of exosomes secretion.
The Kolmogorov-Richardson cascade describes experimentally observed isotropic turbulence of an incompressible fluid, where large-scale flow structures (eddies and vortices) decay into smaller fractal structures. As mentioned, we are currently adherent to the dogma of visualizing protein folding as a stochastic process. However, the simulated protein folding transitions of the SH3-domain (Src Homology 3) protein was shown to obey the vortex dynamics and hydrodynamical equations of Kolmogorov’s statistical theory of turbulence (23, 83). The spatial flow distributions of the probability fluxes were determined to be self-similar (Kolmogorov-Richardson cascades) with a fractal dimension that decreases toward the native state, indicating that the flow becomes more turbulent at more stable protein
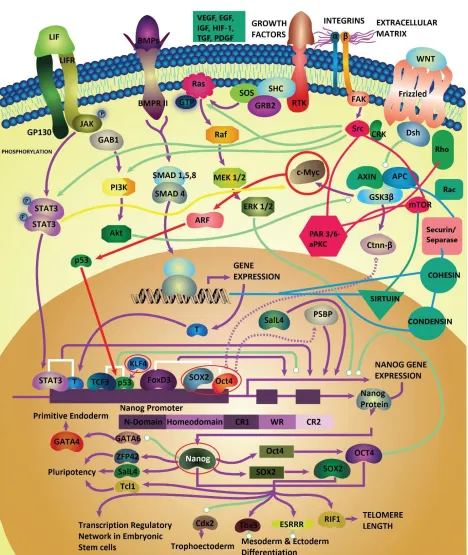
conformations (23, 83). Interestingly, most focal adhesion complexes and tyrosine receptor kinases mediating the initiation and progression of cancer metastasis depend on the structural conformation of SH3 domains (e.g., Src/FAK/Crk/Cas pathways) (Figure 1) (83). Likewise, the folding dynamics of the villin subdomain HP-35 protein in a FRET (Forster resonance energy transfer) experiment was shown to obey the 𝛽-model of turbulence with many orders (scales) of turbulent flow transitions for the eddies in the 3D conformational space (84, 85).
In (86) turbulence was treated as a heat flow problem using the nonequilibrium statistical mechanics of moving fluid particles in three-dimensional lattice boxes. The macroscopic fluid transitions were characterized by Boltzmann-Gibbs distributions. Such a description well applies to the metastable cell states of the Waddington attractor landscape. The critical Reynolds number was shown to be in the order of ~60-100 for Taylor-Couette and Rayleigh-Benard convection systems, which is relatively feasible at biologically relevant scales (86, 87). Recall that the Reynolds number is an ill-defined parameter. For instance, whether one consider the diameter or radius of a pipe will alter the number by 2-fold.
demonstrating that the turbulent energy cascades of the Navier-Stokes equations may help explain collective cell migration and pattern formation within cells (93-95). The cytoskeletal protein
microtubules and actin are being recognized as candidates for these active turbulence models, wherein the swarming and flocking behaviors of proteins can phase-transition from the stochastic Brownian motion to the turbulent regime of the Navier-Stokes equations (95, 96). These findings suggest the role of the Navier-stokes equations in protein-mediated patterning within cancer cells severely lacks experimental investigation although it is surfacing in systems science.
CONCLUSION
In summary, tumor pattern formation is intrinsically a nonequilibrium, nonlinear dynamical system (97). In nonequilibrium statistical mechanics, most systems show variations or patterns in structures. Some general characteristics of nonequilibrium systems include: spatiotemporal chaos, emergence of structures across many length and time-scales (e.g., multifractality), wavenumber/frequency selection, self-organization, pattern formation, transition between states (criticality), stability, boundary
dependence, transient, transport and control (sensitive dependence on order parameters and to
perturbation) (97). These characteristics are exhibited by both complex systems discussed herein: cancer ecosystems and fluid turbulence.
The Navier–Stokes equations have been used as fluid-based models of tumor growth wherein tumor cells can be considered as an incompressible fluid (98). Reaction-diffusion systems governing tumor morphogenesis can exhibit spatio-temporal chaos (i.e., chemical turbulence) (99). Cancer cells are proposed to be strange attractors on the cancer networks’ state-space (Waddington landscape), whereby turbulence governs their dynamics at all scales (processes).
Finding the master clique controlling cancer stemness remains an NP-hard problem in graph-theoretic network reconstruction. Finding fixed-point attractors is an NP-hard problem in computational complexity theory (100 – 104). The above-discussed sections demonstrate that cancer cells behave as strange attractors rather than fixed-points on the network state space. Then, the already challenging NP-hard problem becomes a more complex, intractable problem. A deeper understanding into the fluid dynamics of cancer cell fate transitions and pattern formation/morphogenesis are suggested to better steer these complexity problems.
There are no direct means of testing for the presence of criticality and phase transitions to chaotic behaviors in Gene Regulatory Networks. Finding an efficient algorithm for detecting strange attractors in the state-space of cancer networks remains difficult. However, there are some evidence from
experimental single-cell data and networks inference algorithms that have shown bits and pieces of signaling networks that may confer tumor cells to behave as strange attractors of the Waddington landscape. For example, the epigenetic landscape of GBMs (Glioblastoma Multiforme) shows
tremendous spatiotemporal heterogeneity. However, a core set of neurodevelopmental transcription factors (POU3F2, SOX2, SALL2, OLIG2) were identified to be essential for GBM propagation in brain cancer stem cells (105). Hence, criticality in GRNs must be experimentally tested by perturbing gene expression networks and monitoring single-cell dynamics, as is the case for detecting strange attractor dynamics in experimental turbulence. The turbulence equations are proposed as a solution for
navigating and reconstructing the complexity of cancer stemness networks.
pathway, Notch, VEGF, HIF-1, ECM (extracellular matrix)-remodeling pathways, etc. are shown to induce phenotypic plasticity and regulate the iPSC (induced pluripotency stem cell) network of Yamanaka factors (OCT4, SOX2, c-MYC, KLF4). Moreover, they form feedback loops with the ECM-stroma focal adhesion pathways and differentiation and cancer growth pathways such as
Src/mTOR pathway.
Figure 2: GBM landscape The essential drivers of GBM stemness are shown on a Waddington landscape, based on the findings of reference (105). In speculation, the identified circuitry may cause a normal cell (fixed point attractor) to behave as a cancer stem cell (strange attractor). The multi-colored landscape demonstrates tumor heterogeneity. The attractor X represents a cancer stem cell expressing the SOX2 gene, while the attractor state Y represents a cell fate expressing the SOX2-OCT7 regulatory network. The cell fate Z represents the GBM stem cell expressing all of the identified essential drivers of cancer stemness, and hence, at the highest peak of the mountain (i.e., highest stem cell potential). Note that the OCT7 gene is shown to exhibit a Rössler attractor-like dynamics, whereas the cell fate Z expressing all four factors displays a Lorenz-attractor-like strange attractor as speculated by the flow cytometry data in reference (105). These speculations are not experimentally supported due to the absence of dynamics (time-dependent information) in the datasets.
Data Availability Statement: Data sharing is not applicable to this article as no new data were created or analyzed in this study
REFERENCES
1. Shen SY, Singhania R, Fehringer G, Chakravarthy A, Roehrl MHA, Chadwick D, Zuzarte PC, Borgida A, Wang TT, Li T, Kis O, Zhao Z, Spreafico A, Medina TDS, Wang Y, Roulois
D, Ettayebi I, Chen Z, Chow S, Murphy T, Arruda A, O'Kane GM, Liu J, Mansour M, McPherson JD, O'Brien C, Leighl N, Bedard PL, Fleshner N, Liu G, Minden MD, Gallinger S, Goldenberg A, Pugh TJ, Hoffman MM, Bratman SV, Hung RJ, De Carvalho DD. Sensitive tumor detection and classification using plasma cell-free DNA methylomes. Nature 563, 579-583 (2018)
2. Halvaei, S., Daryani S, Eslami-S Z, Samadi T, Jafarbeik-Iravani N, Bakhshayesh TO, Majidzadeh-A K, Esmaeili R.Exosomes in cancer liquid biopsy: a focus on breast cancer. Mol Ther Nucl Ac. 1:131-141 (2018)
3. Zhou, S., Abdouh, M., Arena, V., Arena, M., & Arena, G. Reprogramming Malignant Cancer Cells toward a Benign Phenotype following Exposure to Human Embryonic Stem Cell Microenvironment. PloS One.12(1): e0169899 (2017)
4. Esteva A, Robicquet A, Ramsundar B, Kuleshov V, DePristo M, Chou K, Cui C, Corrado G, Thrun S, Dean J.A guide to Deep-learning Healthcare. Nat. Med. 25: 24–29 (2019)
5. Printz, C. Spontaneous Regression of Melanoma May Offer Insight into Cancer Immunology. JNCI. 93(14): 1047–1048 (2001)
MJ, Park JB. Pigment epithelium derived factor (PEDF) expression induced by EGFRvIII
promotes self-renewal and tumor progression of glioma stem cells. PLOS Biol. 14(1): e1002367 (2015)
7. Choi D, Montermini L, Kim DK, Meehan B, Roth FP, Rak J.The impact of oncogenic EGFRvIII on the proteome of extracellular vesicles released from glioblastoma cells. Mol Cell Proteomics. 17(10):1948-1964 (2018)
8. Venkatesh HS, Morishita W, Geraghty AC, Silverbush D, Gillespie SM, Arzt M, Tam LT, Espenel C, Ponnuswami A, Ni L, Woo PJ, Taylor KR, Agarwal A, Regev A, Brang D, Vogel H , Hervey-Jumper S, Bergles DE, Suvà ML, Malenka RC, Monje M. Electrical and synaptic integration of glioma into neural circuits. Nature, (7775): 539-545 (2019)
9. Herbach, Ulysse et al., Inferring gene regulatory networks from single-cell data: a mechanistic approach. BMC systems biology vol. 11,1 105. 21 (2017)
10. Kauffman, S. & Levin, S. Towards a general theory of adaptive walks on rugged landscapes. J. Theor. Biol. 128(1):11-45 (1987)
11. Lummertz da Rocha E, Rowe RG, Lundin V, Malleshaiah M, Jha DK, Rambo CR, Li H, North TE, Collins JJ, Daley GQ., Reconstruction of complex single-cell trajectories using CellRouter. Nature Comm. 9: 892 (2018)
12. Perkins, T.J., Foxall, E., Glass, L., & Edwards, R., A scaling law for random walks on networks. Nat. Comm. 5:5121 (2014)
13. Li Q, Wennborg A, Aurell E, Dekel E, Zou JZ, Xu Y, Huang S, Ernberg I.Dynamics inside the cancer cell attractor reveal cell heterogeneity, limits of stability, and escape. PNAS. 113(10):2672-7 (2016)
14. Wang, J., Xu, L., Wang, E. Potential landscape and flux framework of nonequilibrium
networks: robustness, dissipation, and coherence of biochemical oscillations. PNAS 105 (34), 12271–12276 (2008)
15. Mojtahedi M, Skupin A, Zhou J, Castaño IG, Leong-Quong RY, Chang H, Trachana K, Giuliani A, Huang S. Cell fate decision as a high dimensional critical state transition. PLoS Biol. 14(12): e2000640 (2016)
16. Kauffman, S. The origins of order. Oxford University Press, New York. (1993)
17. L Glass, C Hill, Ordered and disordered dynamics in random networks. Europhysics Letters. Volume 41(6): 599-604 (1998)
19. Lin, Y.T., and Nicolas E B. Efficient analysis of stochastic gene dynamics in the non-adiabatic regime using piecewise deterministic Markov processes. Journal of the Royal Society,
Interface vol. 15,138: 20170804 (2018)
20. Wang, J. Landscape and flux theory of non-equilibrium dynamical systems with application to biology. Advances in Physics. Vol 64(1): Pages 1-137 (2015)
21. Metzler, R. Barkai, E. and Klafter, J. Deriving fractional Fokker-Planck equations from a generalised master equation. Europhys. Lett., 46 (4), pp. 431-436 (1999)
22. Lorenz, E.N. Deterministic nonperiodic flow. J. Atm. Sci. 20:130-141 (1963)
23. Kalgin, I.V. & Chekmarev, S.F. Turbulent phenomena in protein folding. Phys Rev E. 83, 011920 (2011)
24. Ruelle, D. Strange Attractors. The Mathematical Intelligencer. 2 (3):126-137 (1979)
25. Dalgleish, A. The relevance of non-linear mathematics (chaos theory) to the treatment of cancer, the role of the immune response and the potential for vaccines. QJM: An International Journal of Medicine, Volume 92, Issue 6, June 1999 pp. 347–359
26. Giomi, L. (2015). Geometry and topology of turbulence in active nematics. Phys. Rev. X 5, 031003.
27. Denk J, Kretschmer S, Halatek J, Hartl C, Schwille P, Frey E. MinE conformational switching confers robustness on self-organized Min protein patterns. PNAS, 115 (18) 4553-4558. (2018)
28. Bryngelson, J D. & Wolynes, P G. Spin glasses and the statistical mechanics of protein folding. PNAS, 84 (21) 7524-7528 (1987)
29. Ruelle, D. Turbulence, strange attractors and chaos. World Scientific Ser A. 16. (1995)
30. Vattay, G., Kauffman, S., & Niiranen,S. Quantum biology on the Edge of Quantum Chaos. PLoS One. 9(3): e89017 (2014)
31. Lambert, N., Chen, Y.N., Cheng, Y.C., Li, C.M., Chen, G.Y., and Nori, F., Quantum Biology. Nature Phys. 9:10-18 (2012)
32. Bohm, D., 1980. Wholeness and the Implicate Order. Routledge, London.
33. Marias, A. et al., The future of quantum biology. J of the Royal Soc. Interface (2018)
34. Löwdin, P.-O., 1963. Proton tunneling in DNA and its biological implications. Rev. Mod. Phys. 35 (3), 725–732.
35. Turing, A.M. The chemical basis of morphogenesis. Philos. Trans. R. Soc. Lond. B: Biol. Sci. 237 (641): 37–72 (1952)
37. Kozlov, V. and Vakulenko, S., On chaos in Lotka-Volterra systems: An analytical approach. Nonlinearity. 26(8) (2013)
38. Wang, F. and Zeng, G. Chaos in a Lotka-Volterra predator-prey system with periodically impulsive ratio-harvesting the prey and time-delays. Chaos, Solitons and Fractals. 32(4): 1499-1512 (2007)
39. Shih, H-S., Hsieh, T-L., and Goldenfeld, N., Ecological collapse and the emergence of travelling waves at the onset of shear turbulence. Nat. Phys. Vol 12: 245-248 (2016)
40. Smale, S. On the differential equations of species in competition. J. Math. Biol. 3(1): 5-7 (1976)
41. Fisher, R.A. The wave of Advance of Advantageous Genes. Annals of Eugenics. 7(4):353-369 (1937)
42. Kolmogorov, A., Petrovskii, I. & Piskunov, N. Study of the diffusion equation with growth of the quantity of matter and its application to a biological problem. Bull. State Univ. Mos., 1-25. (1937)
43. Halatek, J., Brauns, F. & Frey, E. Self-organization principles of intracellular pattern formation. arXiv:1802.07169 [physics.bio-ph] (2018)
44. Dang, Y., Grundel, D.A.J., Youk, H. Cellular Dialogues: Cell-Cell Communication through Diffusible Molecules Yields Dynamic Spatial Patterns. Cell Systems, 10, 82–98 (2019)
45. Halatek, J. & Frey, E. Rethinking pattern formation in reaction–diffusion systems. Nat. Phys. 14: 507-514 (2018)
46. Kim E. J. Y., Korotkevich E. & Hiiragi,T. Coordination of cell polarity, mechanics and fate in tissue self-organization. Trends Cell Biol. 28(7) :541-550 (2018)
47. Li, L., Okura, M., and Imamoto, A., Focal adhesions require catalytic activity of Src family kinases to mediate integrin- matrix adhesion. Molecular and cellular biology. 22(4): 1203-17 (2002)
48. Battle, E., & Clevers, H., Cancer stem cells revisited. Nat Med. 23(10):1124-1134 (2017)
49. Scadden, D.T. The stem cell niche as an entity of action. Nature. 441(7097):1075-9 (2006)
50. Graf, T. & Envers T., Forcing Cells to change Lineage. Nature 462: 587–594 (2009)
51. Heltberg, M.L., Krishna, S., and Jensen, M.H., On chaotic dynamics in transcription factors and the associated effects in differential gene regulation. Nat. Commun. 10(71) (2019)
53. Lens, S.M.A. & Medema, R.H. Cytokinesis defects and cancer. Nat. Rev. Cancer. 19: 32– 45 (2019)
54. Janke, C. The tubulin code: Molecular components, readout mechanisms, and functions. JCB. 206 (4): 461-472 (2014)
55. Huang, Q., Hu, X., He, W., Zhao,Y., Hao, S., Wu, Q., Li, S.,, Zhang, S., and Shi, M., Fluid shear stress and tumor metastasis. American journal of cancer research. 8,5 763-777 (2018)
56. Goetz, J., Metastases go with the flow. Science 362 (6418):999-1000 (2018)
57. Rizvi I1, Gurkan UA, Tasoglu S, Alagic N, Celli JP, Mensah LB, Mai Z, Demirci U, Hasan T.
Flow induces epithelial-mesenchymal transition, cellular heterogeneity and biomarker modulation in 3D ovarian cancer nodules. PNAS, 110 (22) E1974-E1983 (2013)
58. Coffey, D.S. Self-organization, complexity and chaos: The new biology for medicine. Nat Med. 4(8): 882-885 (1998)
59. Baish JW, Jain RK. Cancer, angiogenesis and fractals. Nat Med. 4(9):984. (1998)
60. Baish JW, Jain RK. Fractals and cancer. Cancer Res. 60(14):3683-8 (2000)
61. Lennon FE et al., Lung cancer- a fractal viewpoint. Nat Rev Clin Oncol. 12(11): 664–675. (2015)
62. Brú, A., Casero, D., S.de Franciscis, Herrero, M.A. Fractal analysis and tumor growth.
Mathematical and computer modelling. Volume 47, Issues 5–6, March 2008, Pages 546-559
63. Metze, K., Adam, R., Florindo, J.B., The fractal dimension of chromatin- a potential molecular marker for carcinogenesis, tumor progression and prognosis. Expert Review of Molecular Diagnostics. 4(19):299-312 (2019)
64. Sreenivasan, K.R. and Meneveau, C. The multifractal nature of turbulent energy dissipation. J. Fluid Mech. Vol 224: 429-484 (1991)
65. Hojin, H., Ziegler, M., Welander, M., Bjarnegard, N., Carlhall, C-J., Lindenberger, M., Lanne, T., Ebbers, T., and Dyverfeldt, P., Age-Related Vascular Changes Affect Turbulence in Aortic Blood Flow. Frontiers in physiology vol. 9 36. (2018)
66. Stein PD, and Sabbah HN. Turbulent blood flow in the ascending aorta of humans with normal and diseased aortic valves. Circ Res. 39(1):58-65. (1979)
Endothelial Cells Mediate Resistance to Current Standard-of-Care Therapy. Cancer Res; 77(15) (2017)
68. Posadas EM, Criley SR, Coffey DS. Chaotic oscillations in cultured cells: rat prostate cancer. Cancer Res. (1996)
69. Itik, M. & Banks, S.P. Chaos in a three-dimensional cancer model. international Journal of bifurcation and chaos. 20(01):71 -79 (2010)
70. Ivancevic, T.T., Bottema, M.J., Jain, L.C., A theoretical model of chaotic attractor in tumor growth and metastasis. arXiv.0807.4272 (2008)
71. Letellier, C., Denis F, Aguirre LA., What can be learned from a chaotic cancer model? Journal Theor. Biol. 322:7-16 (2013)
72. Khajanchi, S., Perc, M., and Ghosh, D., The influence of time delay in a chaotic cancer model. Chaos 28, 103101 (2018)
73. Huang, S., Emberg, I., & Kauffman, S. Cancer attractors: a systems view of tumors from a gene network dynamics and developmental perspective. Seminars in cell & developmental biology. 20(7): 869-76. (2009)
74. Miles, J. (1984). Strange Attractors in Fluid Dynamics. Advances in Applied Mechanics Volume 24, 189–214.
75. Takens, F. (1981). Detecting strange attractors in turbulence. Dynamical Systems and Turbulence, Warwick 1980, 366–381
76. Bossomaier, T.R.J. and Green, D.J. Complex Systems. Cambridge Univ. Press. (2000)
77. Scarsoglio, S., Iacobello, G., Ridolfi, L. Complex networks unveiling spatial patterns in turbulence, Int. J. of Bifurcation and Chaos, 26 (13), 1650223, (2016)
78. Glock, P. et al., Design of biochemical pattern forming systems from minimal motifs. eLife. 8: e48646 (2019)
79. Kim, M. Bertram M, Pollmann M, von Oertzen A, Mikhailov AS, Rotermund HH, Ertl G. Controlling Chemical Turbulence by Global Delayed Feedback: Pattern Formation in Catalytic CO Oxidation on Pt (110). Science. Vol. 292, Issue 5520, pp. 1357-1360 (2001)
80. Ouyang, Q. and Swinney, H.L. Transition to chemical turbulence. Chaos 1, 411 (1991)
81. Mecke, K.R. Morphological characterization of patterns in reaction-diffusion systems. Phys. Rev. E 53, 4794 (1996)
K, Sawaguchi A, Harimoto KI, Nakagawa M, Yamamoto T, Handa M, Watanabe N, Nishi E, Arai F, Nishimura S, Eto K. Turbulence activates platelet biogenesis to enable clinical scale ex vivo production. Cell. 174(3): 636-648 (2018)
83. Andryuschenko, V.A. & Chekmarev, S.F. Turbulence in protein folding: vorticity, scaling and diffusion of probability flows. PloS One. 12(2): e0188659 (2017)
84. Andryushchenko, V.A. and Chekmarev, S.F. On hydrodynamic interpretation of folding of an
α-helical protein. Thermophysics and Aeromechanics. Volume 23, Issue 6, pp 941–944 (2016)
85. Chekmarev, S.F. Protein Folding Dynamics in the Space of Experimentally Measured Variables: Turbulence Phenomena. J App Mech. Techn. Phys. Vol. 59, Issue 5, pp 827–833 (2018)
86. Ruelle, D.P. Hydrodynamic turbulence as a problem in nonequilibrium statistical mechanics. PNAS 109 (50) 20344-20346 (2012)
87. Ruelle, D. A Theory of Hydrodynamic Turbulence Based on Non-equilibrium Statistical Mechanics. J. Stat. Phys. 169(6):1039-1044 (2017)
88. Bate, T. E., Jarvis, E.J., Varney, M.E., and Wu, K-T., Collective dynamics of microtubule-based 3D active fluids from single microtubules. Soft matter, 25. (2019)
89. James, M., Bos, W.J. T., and Wilczek, M. Turbulence and turbulent pattern formation in a minimal model for active fluids. Phys. Rev. Fluids, 3(6) (2018)
90. Dunkel, J., Heidenreich, S., Drescher, K., Wensink, H.H., Bär, M. & Goldstein, R.E. Fluid dynamics of bacterial turbulence. Phys Rev Lett. 110(22):228102 (2013)
91. Wensink HH, Dunkel J, Heidenreich S, Drescher K, Goldstein RE, Löwen H, Yeomans JM. Meso-scale turbulence in living fluids. PNAS, 109(36): 14308-13 (2012)
92. Martinez-Pratt et al. Selection mechanism at the onset of active turbulence. Nature Physics, 15:362-366. (2019)
93. Urzay, J., Doostmohammadi, A., & Yeomans, J.M., Multi-scale statistics of turbulence motorized by active matter. J. Fluid. Mech. 822: 762-773 (2017)
94. C. Blanch-Mercader and J. Casademunt, Hydrodynamic instabilities, waves and turbulence in spreading epithelia. Soft Matter, Vol 38. (2017)
95. Bratanov, V., Jenko, F, & Frey, E. “New class of turbulence in active fluids” PNAS. 112,49:
15048- 53. (2015)
97. Cross, M. and Greenside H., Pattern Formation and Dynamics in nonequilibrium systems. Cambridge University Press. (2009)
98. Vauchet, N. and Ztorska, E. Incompressible limit of the Navier—Stokes model with a growth term. Nonlinear Analysis, 163: 34-59 (2017)
99. Kuramoto Y. Chemical Turbulence. In: Chemical Oscillations, Waves, and Turbulence. Springer Series in Synergetics, vol 19. Springer, Berlin, Heidelberg (1984)
100.Akutsu, T., Kuhara, S., Maruyama, O., and Miyano, S., A system for identifying genetic networks from gene expression patterns produced by gene disruptions and overexpressions. Genome Informatics 9, 151-160 (1998)
101.Milano, M. and Roli, A., Solving the satisfiability problem through Boolean networks. LNCS Vol. 1792. Advances in AI: 72-83. (2000)
102.Barillot, E. et al., Computational systems biology of cancer. CRC Press, Taylor & Francis Group. (2013)
103.Songjian, L. et al. An exact algorithm for finding cancer driver somatic genome alterations: the weighted mutually exclusive maximum set cover problem. Algorithms for molecular biology: AMB vol. 11:11. (2016).
104.Hajkarim, M.C., Upfal, E. and Vandin, F. Differentially mutated subnetworks discovery. Algorithms for Molecular Biology volume 14, Article number: 10 (2019)