The specificity of B-cell response in multiple sclerosis
Full text
Figure

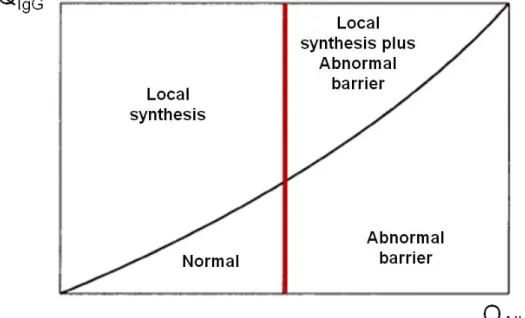
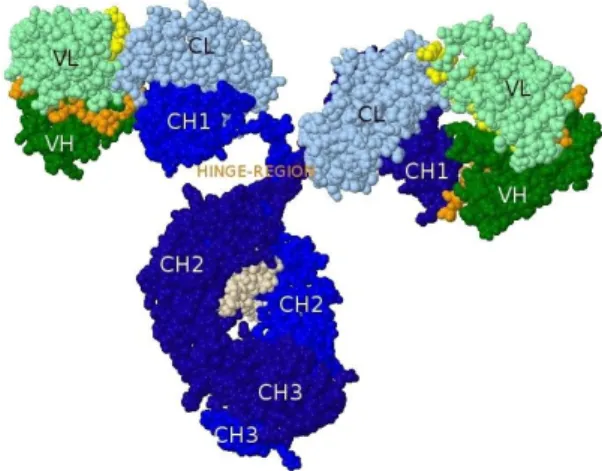
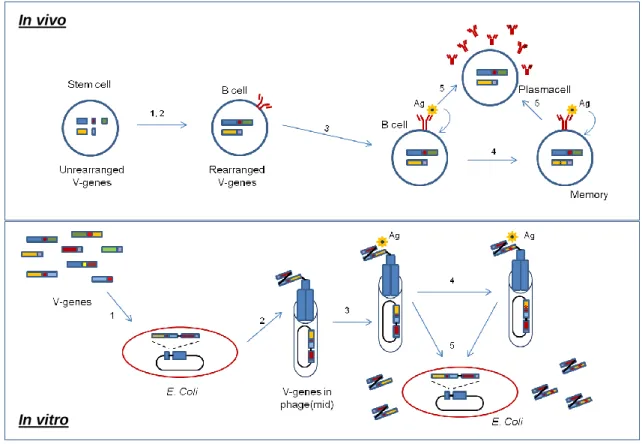
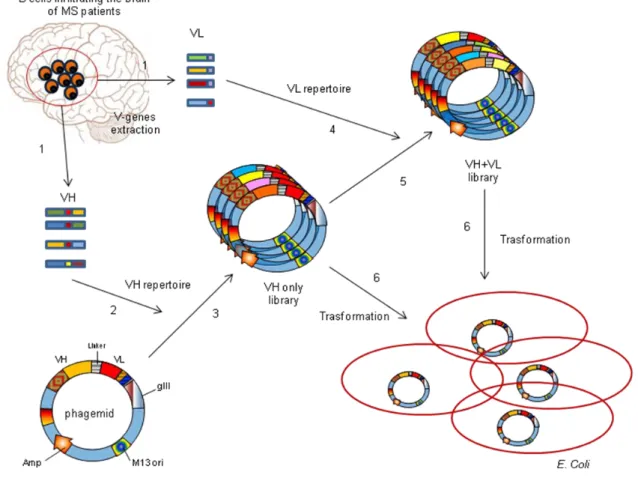
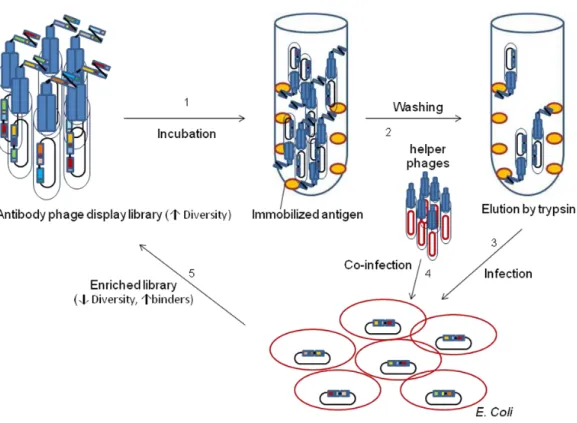
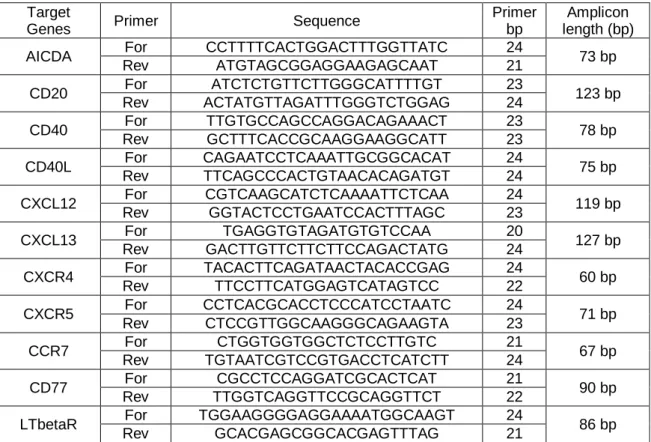
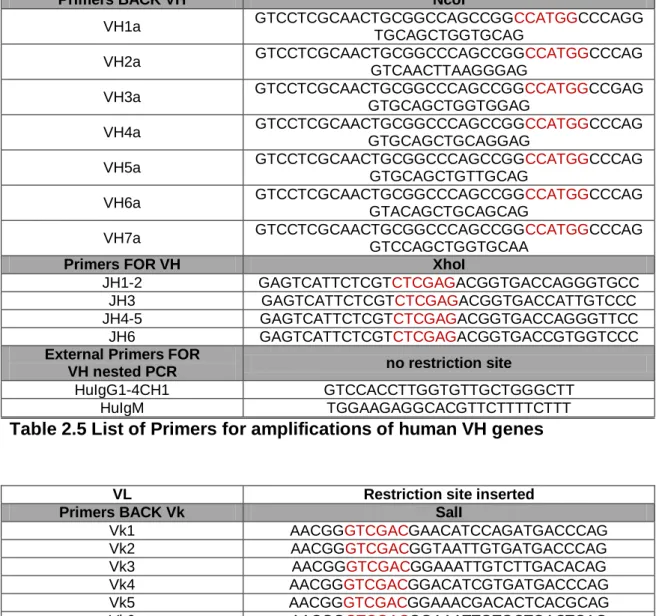
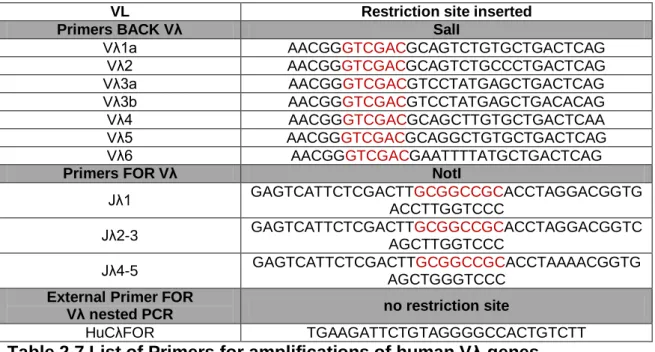
Related documents
19% serve a county. Fourteen per cent of the centers provide service for adjoining states in addition to the states in which they are located; usually these adjoining states have
Through my involvement with Homeless Pride and the Duncan Plaza encampment over the next six months, I met people who continue to be among my closest friends and colleagues in
Methods: After refining the training protocol in 2 chronic patients, 8 sub-acute stroke patients were randomised to receive additional therapy with the Finger Trainer for 20 min
MAP: Milk Allergy in Primary Care; CMA: cow’s milk allergy; UK: United Kingdom; US: United States; EAACI: European Academy of Allergy, Asthma and Clinical Immunology; NICE:
Also, both diabetic groups there were a positive immunoreactivity of the photoreceptor inner segment, and this was also seen among control ani- mals treated with a
1 In [4] authors presents a Priority based dynamic load balancing .It works better for heterogeneous cloud computing systems and is for balancing non-preemptive
