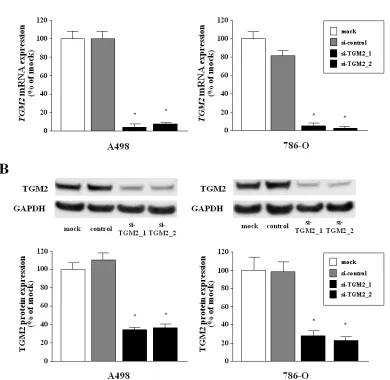
Tumor suppressive microRNA-1285 regulates novel molecular targets:Aberrant expression and functional significance in renal cell carcinoma
Full text
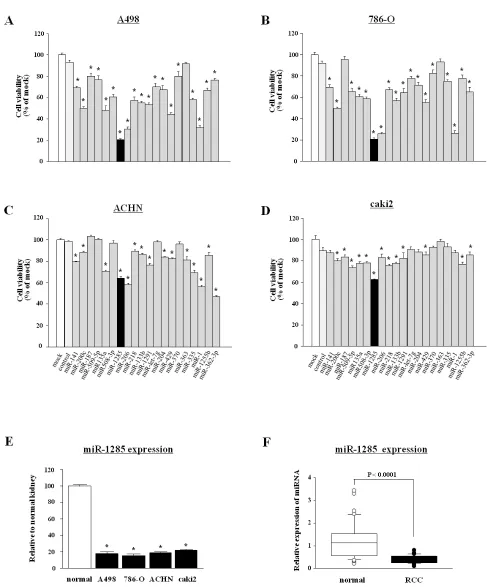
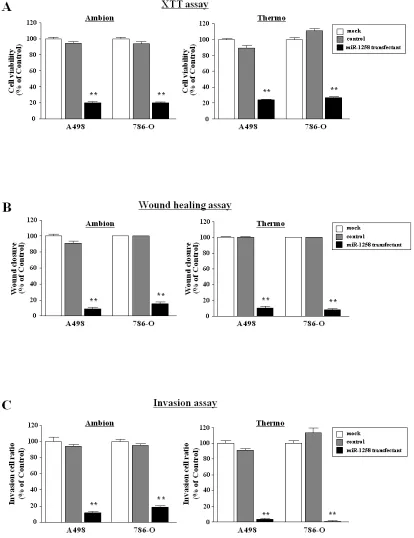
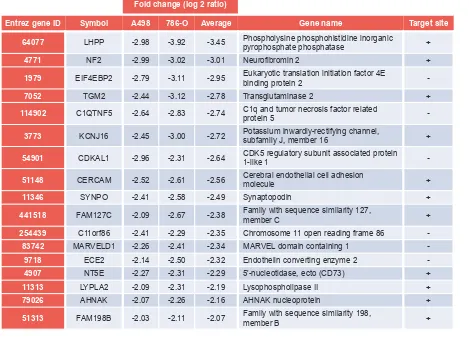
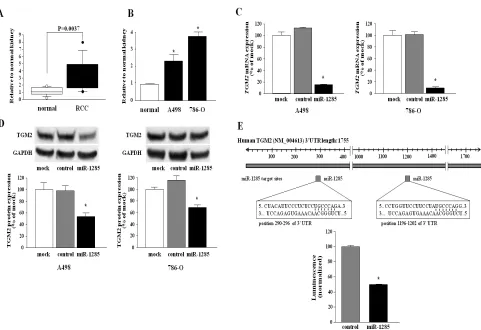
Figure



Related documents
This subsection evaluates success of SMT-based attacks with random cube PCs instead of random PCs. To do so, 50 instances of randomly selected cube plaintexts for each size 2 2 to 2
Assuming that the target region is FL24, this requires 2014 data available in USAGE-TERM for: the value of capital in FL24; the value of public expenditures in FL24’s city (FL24, FL23
This analysis aimed to study the cost and burden of neuropsychiatric disorders among refugees from Syria and Iraq requiring exceptional, United Nations-funded care in a country of
Financial costs associated with community-based targeting in Burkina Faso amounted to USD 5,73 and the economic costs to USD 11,83 per ultra-poor person identified.. Economic
Multidrug-resistant and extended-spectrum beta-lactamase (ESBL)- producing Salmonella enterica (serotypes Typhi and Paratyphi A) from blood isolates in Nepal: surveillance
The CCPA sets forth one exception to this broad rule against cable operators' exercise of editorial control over the content of PEG channels, namely that "a
In form, novel is comparable to nationalism because it is not an epic, where the representation of Reality is still wholesome and “beautiful”, but it pretends to
At 8 weeks, the tendons of the semitendinosus, gracilis, sartorius and semimembranosus muscles were straight and inserted into the initial shaft-like proximal end of the tibia on
