Functional roles of the rat nucleus accumbens: further investigations using microinjection, lesion and electrochemical techniques
Full text
Figure
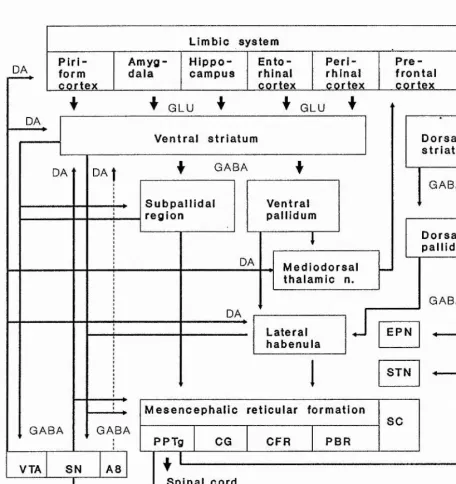
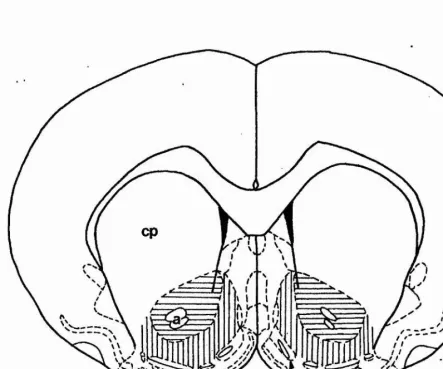

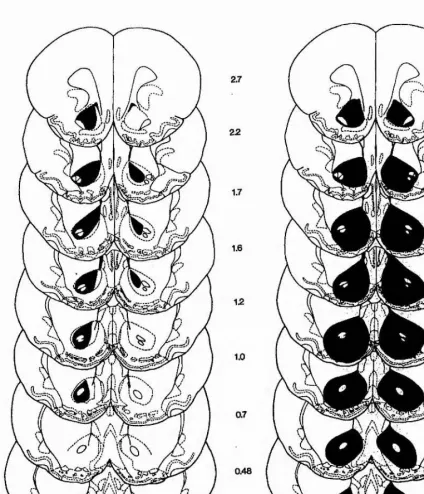
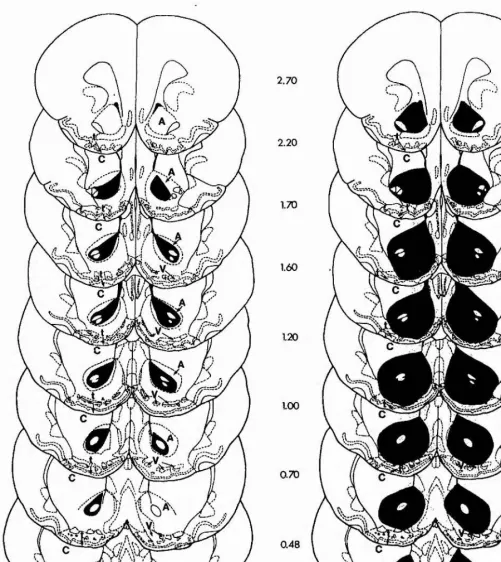



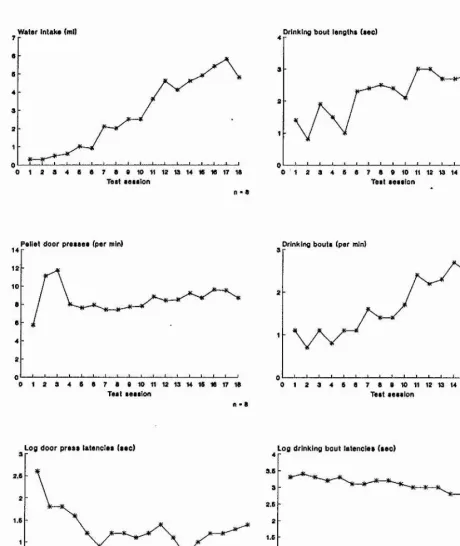
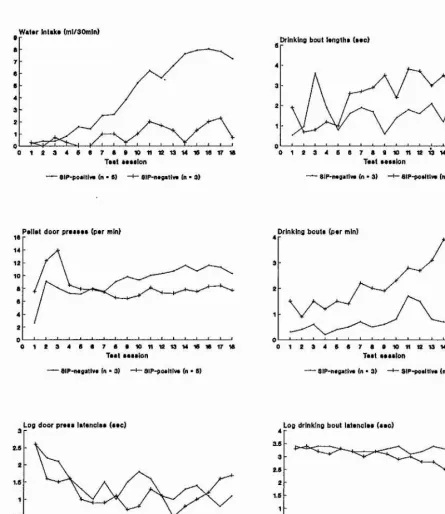
Outline
Related documents
hypertrophic cardiomyopathy or left ventricular hypertrophy based on race and gender in teenagers using screening echocardiography. Cardiovascular Ultrasound 2010
Integration as a service solutions are beginning to find ways of simplifying integration issue on a cloud-to-cloud platform while SaaS vendors are addressing in house
Other numerical techniques which were used to solve the Bratu’s problem include the shooting method [ 1 ], fi- nite element method [ 9 ], homotopy analysis method [ 13 ] and the
Keywords: Alzheimer ’ s disease (AD), Brain, Ankyrin 1 (ANK1), DNA methylation (5mC), DNA hydroxymethylation (5hmC), Entorhinal cortex (EC), Epigenetics, Epigenome-wide
The Court's interpretation of the requirements of both exceptions was ex- tremely narrow, though correct. As to the prior recorded testimony excep- tion, the Court properly
Keywords: data mining applications, support vector machines, decision trees, naive Bayes, thoracic surgery.. 1
Abnormal uterine bleeding in women belonging to different age groups show variable endometrial pattern on histopathological examination of samples obtained from
On the basis of the numbers available, no significant differences were detected between the groups treated with or without iliac crest bone graft with regard to the scores on
