Original Article
Altered long noncoding RNA profiles in rat
ischemia-reperfusion injury after IONPs administration
Yinghao Pei1, Haidong Zhang1, Bo Chen2,3, Linfeng Dai1, Xing Wang1, Xiquan Zhang3,4, Fei Xiong2,3, Hongmei Gu4, Ning Gu2,3
1Department of Intensive Care Unit, Jiangsu Provincial Hospital of Traditional Chinese Medicine, The Affiliated
Hospital of Nanjing University of Traditional Chinese Medicine, Nanjing, PR China; 2State Key Laboratory of
Bioelectronics, Jiangsu Key Laboratory for Biomaterials and Devices, School of Biological Science and Medical Engineering, Southeast University, Nanjing, PR China; 3Collaborative Innovation Center of Suzhou Nano-Science
and Technology, Suzhou, PR China; 4Research Institute of Chia Tai Tianqing Pharmaceutical Group Limited by
Share Ltd., Nanjing, PR China
Received May 18, 2017; Accepted August 1, 2017; Epub August 15, 2017; Published August 30, 2017
Abstract:Background and aim: Iron oxide nanoparticles (IONPs) can protect hearts from ischemia reperfusion (I/R) injuries. Long noncoding RNAs (LncRNAs) have been believed to play important roles in regulation of many
biologi-cal processes. The aim of our study was to investigate the lncRNA profile in I/R injury rat model following IONPs
ferumoxytol administration. Methods: Models of I/R were induced by 30 min of ischemia followed by 24 hours of reperfusion in male Sprague-Dawley rats. The rats were randomized into three groups: sham group, I/R group,
and ferumoxytol group. The profiles of lncRNAs and mRNAs were confirmed using Illumina RNA-Seq technology
combining quantitative real-time PCR analysis. Results: Ferumoxytol leads to a significant reduction in the area of
myocardial infarction. Iron deposition was observed around ischemic myocardial tissue by Prussian blue staining. Sequencing revealed the existence of a total of 19754 lncRNAs distributed in all chromosomes of the experimented rats. In ferumoxytol intra-myocardial injected heart, 433 lncRNAs were up-regulated, whereas 287 lncRNAs were down-regulated when compared with I/R injured hearts (fold-change >2.0). In I/R injury hearts, 419 lncRNAs were up-regulated, whereas 393 lncRNAs were down-regulated when compared with sham heart (fold-change >2.0). Top 10 lncRNAs were selected for RT-qPCR validation. Based on cis-acting target genes prediction and mRNA-sequenc-ing results, NONRNOT219022-Reg3b is likely to be the key lncRNA-mRNA pair. Conclusion: We identified a set of
lncRNAs aberrantly expressed in myocardial I/R injury rats with ferumoxytol, shedding lights for further research of the mechanism of feurmoxytol in myocardial protection and proposing a potential therapeutic target for I/R injuries.
Keywords: Ischemic reperfusion injury, heart, ferumoxytol, long noncoding RNA, profile
Introduction
Coronary artery diseases have become one of the major causes of mortality and morbidity worldwide [1]. As the most successful thera-peutic strategy, pharmacological or mechanical
restoration of coronary blood flow has been
established to be essential for preserving via-ble cardiomyocytes after myocardial infarction
[2]. Although reperfusion therapy is efficient in
alleviating myocardial injuries, reperfusion itself is a double-edged sword which may also
trigger a localized inflammatory response and
regional oxidative burst that leads to cell dam-age and even death. This pathophysiologic
pro-cess is defined as ischemia/reperfusion injury
(I/R injury) [3].
Global genomic and transcriptomic analyses
have identified that eukaryotic genomes are
transcrip-tional level, but also can regulate genes at the post-transcriptional and post-translational level [7]. It is now becoming evident that lncRNAs participate in a multitude of processes that
include chromatin modification, cell differentia -tion and prolifera-tion, transcrip-tion, transla-tion and other important biological processes [8]. In addition, lncRNAs possess close relationships in diverse types of human diseases, including cardiovascular diseases [9]. It has been dem-onstrated that drug administration can regulate levels of quite a few key lncRNAs to mediate pathophysiological process [10].
Iron oxide nanoparticles (IONPs) are generally considered as inert materials and have been
extensively used in the fields of cancer treat -ment and diagnosis [11, 12]. Recently, it has been reported that IONPs can mediate the cel-lular gap junctional crosstalk to improve
mes-enchymal stem cells efficacy for myocardial
infarction [13]. In the previous study, we found that IONPs can protect heart from ischemic reperfusion damage at the animal, tissue and cell level by maintaining the integrity of NPs but not surface charges or integrated molecules [14, 15].
So far, a few iron oxide nanomedicine have been used in the related research. While on the other hand, previous IONPs existed a host of problems, e.g. safety in vivo applications. Consequently, we chose ferumoxytol instead of ordinary IONPs in our research. Ferumoxytol is a kind of iron oxide nanomedicine, approved by USA Food and Drug Administration for
treat-ment of iron deficiency in clinic on 30 June
2009, and also widely used as magnetic reso-nance imaging agent in clinical trials. In this work, we explored the expression differences of lncRNA in I/R injury rat model following feru-moxytol administration. We found that intra-myocardial injection of ferumoxytol could pro-tect rat heart from I/R injury. At the same time, after high-throughput RNA-sequencing detec-tion with real-time PCR (RT-qPCR) validadetec-tion, we found that NONRNOT219022 might be the key lncRNA in the cardio-protection of feru-moxytol. GO and KEGG analyses were indica-tive of biological functions related to NADP+ -dependent isocitrate dehydrogenase (IDP) activity and citrate cycle (TCA cycle). The results may provide a crucial foundation for future pharmacology research on the roles of lncRNAs in ferumoxytol cardio-protection.
Materials and methods
Experimental animals and reagents
Adult male Sprague-Dawley rats weighing 280-320 g were purchased from the Animal Center of Jinling Hospital (Nanjing, China) and
main-tained for at least 1 week in a specific patho -gen-free animal facility to allow adjustment to the environment. The animals were housed in groups of eight and maintained in controlled conditions of temperature (22 ± 2°C), humidity (60-70%) and under a 12-h-light-dark cycle (lights on 07:00 AM). Food and water were available ad libitum. All procedures were per-formed in accordance with the guidelines of the National Research Council’s Guide for the Humane Care and Use of Laboratory Animals. Ferumoxytol were synthesized according to our previous work [16]. All procedures for the care and use of animals were performed in accor-dance with the guidelines of the National Research Council’s Guide for the Humane Care and Use of Laboratory Animals.
In vivo myocardial I/R model
Rats were anesthetized by an intraperitoneal injection of 3 ml/kg 10% chloral hydrate. The rats were intubated and mechanically ventilat-ed with room air using a rodent respirator (DV-2000, Shanghai Jia Peng Technology Co., Ltd., China). A limb lead II electrocardiogram (ECG) was recorded. An oscilloscope electrocardio-gram monitor was used to display the electro-cardiogram continuously throughout the proce-dure. A left thoracotomy was performed to expose the hearts. Ligation of the left anterior descending (LAD) artery was performed 2-3 mm from its origin between the pulmonary artery conus and the left atrium using a 6-0 silk prolene suture. A small vinyl tube was placed on top of the vessel to form a snare for revers-ible coronary occlusion. Myocardial cyanosis and ECG evidence of injury were used to
con-firm successful occlusion of the LAD artery. The
heart was subjected to regional ischemia for 30 min, followed by coronary reperfusion through release of the slipknot.
Experimental groups
occlu-sion and followed by 24-h reperfuocclu-sion, 30 μL
dextran was injected around the pale areas
(upper, center and lower with 10 μL in each
area) 5 min before reperfusion; 3) I/R+ feru-moxytol group: 0.5 mg/kg feruferu-moxytol diluted
in 30 μL dextran was injected around the pale areas (upper, center and lower with 10 μL in
each area) 5 min before reperfusion.
Measurement the size of myocardial infarction
Infarction size (n=4 in each group) was evalu-ated by triphenyl tetrazolium chloride (TTC)
staining as described previously [17]. Briefly,
the heart was isolated, frozen and sectioned perpendicular to the long axis (1.5-mm thick) up to the area of ligation. The slices were incu-bated in 1% TTC solution at 37°C for 20 min to visualize the infarcted area and transferred to 10% neutral buffer formalin overnight at room temperature to stabilize the staining. The area stained red by TTC represented ischemic but viable tissue. The infarcted myocardium was not stained by TTC and was paler than the TTC-stained area. The rates of infarct area (IA) were determined by computerized planimetry using Image Pro Plus software (Media Cybernetics, Rockville, MD, USA).
Measurements of LDH and CK-MB
After 24 hours of reperfusion, blood samples were collected from the right ventricle and cen-trifuged at 3000× g for 10 min to isolate serum. Myocardial cellular damage was evaluated by measuring lactate dehydrogenase (LDH) and creatinine kinase MB (CK-MB) activity in plas-ma using commercially available assay kits (Sigma-Aldrich).
of iron deposition. A digital microscope with a multi-functional image analysis system (Leica DM 750 + DFC 450C, Leica Microsystems, Wetzlar, Germany) was used to analyze each
section at 400× magnification for determining
the positive Prussian blue staining. RNA extraction and purification
Myocardial tissue of rats (n=8 in each group) were collected after 24 h reperfusion for RNA sequencing. Total RNA was isolated using
RNAiso Plus (TAKARA, Japan). Strand-specific
libraries were prepared using the TruSeq® Stranded Total RNA Sample Preparation kit (Illumina, USA) following the manufacturer’s
instructions. Briefly, ribosomal RNA was
re-moved from total RNA using Ribo-Zero rRNA
removal beads. Following purification, the
mRNA is fragmented into small pieces using divalent cations under 94°C for 8 min. The
cleaved RNA fragments are copied into first
strand cDNA using reverse transcriptase and random primers. This is followed by second strand cDNA synthesis using DNA Polymerase I and RNase H. These cDNA fragments then go through an end repair process, the addition of a single ‘A’ base, and then ligation of the
adapt-ers. The products are then purified and enriched with PCR to create the final cDNA library. Purified libraries were quantified by Qubit® 2.0 Fluorometer (Life Technologies, USA) and vali-dated by Agilent 2100 bioanalyzer (Agilent
Technologies, USA) to confirm the insert size
[image:3.612.86.410.84.244.2]and calculate the mole concentration. Cluster was generated by cBot with the library diluted to 10 pM and then were sequenced on the Illumina HiSeq 2500 (Illumina, USA). The library Table 1. RT-qPCR Primers for 10 lncRNAs and U6
Gene Forward primer Reverse primer
NONRATT214498 5’-AGGGAGACCGAGGGTTTGA-3’ 5’-AGGCCAGGTGGACCTTGTG-3’ NONRATT226056 5’-TCTGAGCACGCATTCCC-3’ 5’-ACAACCGACTGGCTGAAC-3’ NONRATT219023 5’-GCACCTCTTTACTTCATCC-3’ 5’-AAAGGGAAACAAGAATACC-3’ NONRATT211868 5’-CAAGGTGCTGAAGCAAGTGC-3’ 5’-CGCTCGAAGATGTCGTTCAC-3’ NONRATT202015 5’-GAAAGTCTCCCTCATCTACG-3’ 5’-CATTCCTCCTGAACCAAG-3’ NONRATT219951 5’-CTCCTTCTTGGGTCTACG-3’ 5’-TCACCACAGCGATAACAG-3’ NONRATT228656 5’-CCTGGCCTATGCCTCACA-3’ 5’-CCAACGGTAGTGCCTGCT-3’ NONRATT208813 5’-ACAAACACGGCTGTAGATT-3’ 5’-GGTTGAGTATGGGAAGGA-3’ NONRATT219022 5’-AGCCACCTTTCTTCCATTT-3’ 5’-CTACATTCCCTCCCAACC-3’ NONRATT200186 5’-AGCATTGTCCTGGAAGTCG-3’ 5’-TCTGAACCAAGAGCACCCT-3’
U6 5’-CTCGCTTCGGCAGCACA-3’ 5’-AACGCTTCACGAATTTGCGT-3’
Histological observa-tions
Myocardial tissues (n= 4 in each group) were
fixed in 4% paraformal -dehyde solution for 24 hours and
embed-ded in paraffin. Using a microtome, 5
μm-thi-ck serial sections were
cut from the paraffin
construction and sequencing was performed at Shanghai Biotechnology Corporation.
Data analysis for gene expression
Sequencing raw reads were preprocessed by
filtering out rRNA reads, sequencing adapters,
short-fragment reads and other low-quality reads. Tophat v2.0.9 [18] was used to map the cleaned reads to the Rat rn6 reference genome with two mismatches. After genome mapping,
Cufflinks v2.1.1 [19] was run with a reference
annotation to generate FPKM values for known gene models. Differentially expressed genes
were identified using Cuffdiff [19]. The p-value
significance threshold in multiple tests was set
by the false discovery rate (FDR) [20, 21]. lncRNAs identification and expression analysis
Cufflinks was used to assemble reads into tran
-scripts. Putative lncRNAs were defined as novel
transcripts set through the following filters: length ≥200 bp; number of exons ≥2; ORF≤300
bp; have no or weak protein coding ability (CPC
score <0 [22] & CNCI score <0 [23] & no signifi -cant similarity with Pfam database [24]). Finally, to generate a unique set of lncRNAs, we used Cuff compare to integrate the RNA-seq derived lncRNAs with the known lncRNAs previously annotated by NONCODE v4. Differentially ex- pressed lncRNAs were selected for target pre-diction. The genes transcribed within a 10 kb window upstream or downstream of lncRNAs were considered as cis-acting target genes. The trans-acting target genes were predicted using RNAplex software [25].
RT-qPCR
LncRNA expression was quantified by RT-qPCR
with SYBR Green (Life Technologies, CarIsbad,
CA) as the fluorescent reporter. Total RNA (1 μg)
[image:4.612.93.311.71.455.2]was used to synthesize the first strand of cDNA
using M-MLV Reverse Transcriptase (Life Technologies). The RT-qPCR was performed as previously described. Primers for 10 lncRNAs and U6 are listed in Table 1. The relative levels of target lncRNAs were calculated using the
ΔΔCT method, and the data were normalized
with U6.
Statistical analysis
For lncRNA expression by RNA-Seq, signifi -cance levels were indicated as P<0.05 and greater than two-folds of change. Data obtained from RT-qPCR are presented as the means ± standard error (SE) and analyzed using a stu-dent t-test in SPSS version 20.0 software (SPSS, Chicago, IL, USA).
Results
Myocardial infarction size
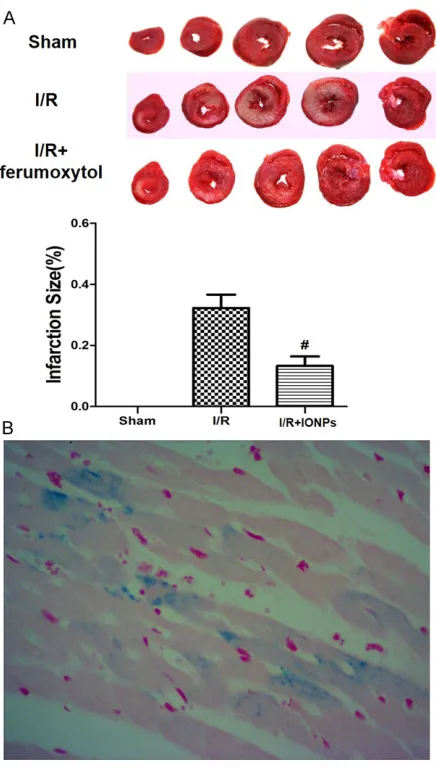
Representative images of the TTC staining for rats in each group were shown in Figure 1A.
by BoHao Bio-tech (Shanghai, China) using sequencing technology. In total, we acquired 194.7 million readings of 24.3 gigabase (Gb) of mRNA sequence. Sequencing revealed the existence of a total of 19754 lncRNAs that were ubiquitously distributed in all chromosomes including sex chromosomes of rats (Figure 2 and Table S1). A lncRNA that FPKM>1 and at least a 2-fold up-regulation or 0.5-fold down-regulation can be thought as a candidate lncRNA which may receive further validation. In ferumoxytol intra-myocardial injected heart, 433 lncRNAs were up-regulated, whereas 287 lncRNAs were down-regulated when compared with I/R injury heart (Table 2). In I/R injury heart, 419 lncRNAs were up-regulated, where-as 393 lncRNAs were down-regulated when compared with sham heart (Table 2).
Validation of key lncRNA
To validate the result of sequencing, the top 10 lncRNAs with both increased and decreased expression were selected for RT-qPCR (Table Figure 2. The genomic map of total lncRNAs in chromosomes of rats. The distribution of lncRNAs in chromosomes of sham group, I/R group and I/R+ ferumoxytol are marked in green, red and yellow, respectively. Sequencing revealed the existence of a total of 19754 lncRNAs in our study. The numbers and symbols represent chromosome numbers.
Table 2. Changes in lncRNAs
Fold change 2 5 10 Total
IONPs vs. I/R injury Up-regulated 376 51 6 433 Down-regulated 240 44 3 287 I/R injury vs. Sham Up-regulated 368 45 6 419 Down-regulated 336 55 2 393
Compared with rats in I/R group, feru-moxytol administration led to a sig-
nificant reduction in the area of myo -cardial infarction induced by I/R injury (13.1 ± 3.7% vs. 33.6 ± 4.3%, P<0.05). Prussian blue staining was applied to visualize the iron deposition around ischemia myocardial tissue as shown in Figure 1B.
Serum levels of CK-MB and LDH were used as myocardial injury markers in our study. Both the plasma CK-MB and
LDH levels were significantly increased
in rats of the I/R group compared with those in the sham group. Consistently, ferumoxytol treatment reduced the levels of CK-MB and LDH as compar- ed with those in the rats of the I/R group (CK-MB: 2898.3 ± 722.1 U/L vs. 4256.4 ± 532.6 U/L, LDH: 1322.6 ± 264.3 U/L vs. 2346.3 ± 313.6 U/L, respectively; both P<0.05. Figure 1C). Determination of lncRNA profile
LncRNA profile in the myocardial tissue
of rats in each group was determined Table 3. Top 10 lncRNA with differential expression
lncRNA Fold change in sequencing Fold change in RT-qPCR IONPs vs.
I/R injury I/R injury vs. Sham IONPs I/R injuryvs. I/R injury vs. Sham
NONRNOT214498 0.05 4.00 0.10 18.64
NONRNOT226056 0.07 16.50 0.59 1.87
NONRNOT202015 0.07 2.55 0.78 1.40
NONRNOT219951 0.10 7.44 0.32 1.63
NONRNOT228656 0.12 2.67 0.59 1.87
NONRNOT208813 10.56 0.66 2.02 0.62
NONRNOT200186 11.06 0.86 2.78 0.61
NONRNOT219022 11.81 0.80 10.79 0.42
NONRNOT219023 14.19 0.29 0.46 2.51
3). The results suggested two lncRNAs (NO- NRNOT214498 and NONRATT219022) that were strongly consistent with those of sequenc-ing (Table 3). Based on cis-acting target genes prediction and mRNA-sequencing results, NONRNOT219022-Reg3b might be the key lncRNA-mRNA pair (Table 4). RT-qPCR inferred that expression levels of Reg3b in I/R heart were 0.11 and 0.34-fold down-regulated when compared with ferumoxytol heart and sham heart, respectively.
Gene ontology and KEGG analysis
Among the GO ontology analyses, the top 3 terms included the following: IDP activity, 2-oxo-glutarate metabolic process and voltage-gated potassium channel activity (Figure 3A). Based on the KEGG pathway analysis, the most 3 enriched pathways were TCA cycle, glutathione metabolism and vascular smooth muscle con-traction (Figure 3B).
Discussion
Ischemia reperfusion is a complex process with an army of factors, such as calcium overload and altered energy metabolism, which
contrib-ute to the final damage to the myocardium.
Numerous interventions and agents have been
identified as prophylaxis for cardiac I/R injuries,
such as erythropoietin [26], omega-3 polyun-saturated fatty acids [27], and adipokine nesfa-tin-1 [28]. However, translation of these strate-gies and agents to the clinical setting has been far from satisfying. In the previously study, we found IONPs could protect hearts from isch-emia reperfusion injury in vivo as well as in vitro
[14].
NPs are submicron-sized polymeric particles with a therapeutic agent absorbed onto their surface or within the matrix [29]. Direct intra-muscular injection bone marrow cells [30] or growth factor [31] into the border zones of the
infarct has been extensively investigated for reducing ischemia reperfusion injury. In this
work, we reported for the first time that
intra-myocardial injection of IONPs (ferumoxytol) were safe and feasible. Prussian blue staining was applied to visualize the iron deposition around ischemia myocardial tissue.
At the same time, we explored the lncRNA
profile in the ischemia/perfusion injury rat
model following ferumoxytol treatment. High-throughput RNA-sequencing detection revealed a total of 19754 lncRNAs in rat hearts. Four-hundred and thirty-three lncRNAs were elevat-ed, and 287 lncRNAs were reduced in feru-moxytol heart compared with I/R injury heart. In I/R injury heart, 419 lncRNAs were up-regu-lated, whereas 393 lncRNAs were down-regu-lated when compared with sham heart. Ten of the dysregulated lncRNAs with most meaning-ful change in expression fold were further vali-dated by RT-qPCR assays. Based on target genes prediction and sequencing-PCR valida-tion, we found that NONRNOT219022-Reg3b might be one of the key lncRNA-mRNA pairs in the cardiac-protection of ferumoxytol. Reg3b was originally discovered in a rat pancreatitis model [32] and was later found to promote wound repair [33] and neuronal regeneration [34]. Reg3b has been indicated to be an essen-tial regulator of macrophage recruitment and expansion to the damaged ischemia myocardi-um [35]. Resident macrophages contribute to local homeostatic processed by clearing dying cardiomyocytes and excess neutrophil, which lead to matrix degradation reduction and colla-gen deposition promotion. Loss of Reg3b can lead to a large decrease in the number of mac-rophages in the ischemic heart accompanied
by increased ventricular dilatation and insuffi -cient removal of neutrophils [35].
Zou et al. reported that NONRATT021972 ex-
[image:7.612.90.520.85.175.2]pression was significantly higher in the serum Table 4. Chromosomal location and gene ontology of two key LncRNA and their linked genes
LncRNA Chromosomal location Cis-target gene Trans-target gene
NONRNOT214498 Chr2:216941790-216950409 Col11a1 -NONRATT219022 Chr4:109468916-109470508 Reg3a, LOC100361595,
AC115202.1, Reg3b Zfp84Myo18a AABR07007905.1 AABR07027902.1 Asic1 Ctnna3
of myocardial infarction (MI) patients and stel-late ganglia of rat model [36]. Treatment with NONRATT021972 siRNA in the rat model of MI could inhibit the up-regulation of P2X7 recep-tors in the stellate ganglia and ameliorate myo-cardial ischemic damage [36]. In this research,
we did not found significant fluctuation of
NONRATT021972 after I/R operation or feru-moxytol administration. This might because
that lncRNA profile of our work was evaluated
from heart tissue in a I/R model but not stellate ganglia in a MI model.
GO terms describe the molecular actions of gene products, suggesting biological pro-cessed, components, or structures in which individual genes and proteins are known to be involved. The enrichment of GO terms calculat-ed bascalculat-ed on the p value can be suggestive of the feasible functions of the gene involved. Our data revealed the biological process related to the energy metabolism and ion channel, such as IDP activity, 2-oxoglutarate metabolic pro-cess and voltage-gated potassium channel activity. KEGG pathway analysis is widely used to gain insight into the underlying biology of dif-ferentially expressed genes and proteins. According to our data, TCA cycle, glutathione metabolism and vascular smooth muscle
con-traction were the most significant pathway
putatively affected by the differentially ex- pressed lncRNAs. As a member of isocitrate dehydrogenases that catalyze oxidative
decar-boxylation of isocitrate into α-ketoglutarate, IDP is responsible for fluctuation of NAPD+/ NADPH in cytoplasm. NADPH is an essential cofactor in the metabolism of glutathione (GSH) and thioredoxin, which plays a key role in the antioxidant system. It has been demonstrated that IDP is reduced under condition of I/R and changes of IDP activity may have a regulatory
significance for the functioning of the myocar -dial antioxidative system [37, 38].
To the best of our knowledge, this is the first research to demonstrate the lncRNA profile
changes and related importance in myocardial I/R injury after ferumoxytol intra-myocardial administration. The results of our exploration
may provide a foundation for future research on the roles of lncRNA in ferumoxytol cardio-protection. How this regulation process oper-ates at the epigenetic level needs to be further explored. Further research on the biological functions, molecular mechanisms and signal-ing pathways of the lncRNA are deserved to elu-cidate pharmacological mechanism of feru-moxytol, a potential therapeutic targets of novelty for I/R injuries.
Acknowledgements
This work was supported by grants from the National Natural Science Foundation of China (NSFC) for Key Project of International Coo- peration (61420106012).
Disclosure of conflict of interest
None.
Address correspondence to: Xing Wang, Department of Intensive Care Unit, Jiangsu Provincial Hospital
of Traditional Chinese Medicine, The Affiliated
Hospital of Nanjing University of Traditional Chinese Medicine, 155 Han Zhong Road, Nanjing 210029, PR China. E-mail: [email protected]; Ning Gu, State Key Laboratory of Bioelectronics, Jiangsu Key Laboratory of Biomaterials and Devices, School of Biological Science and Medical Engineering, Southeast University, 2 Sipailou, Nanjing 210096, PR China. E-mail: [email protected]
References
[1] Heusch G, Libby P, Gersh B, Yellon D, Böhm M, Lopaschuk G and Opie L. Cardiovascular re-modelling in coronary artery disease and heart failure. Lancet 2014; 383: 1933-1943. [2] Ibrahim AW, Riddell TC and Devireddy CM.
Acute myocardial infarction. Crit Care Clin 2014; 30: 341-364.
[3] Ibáñez B, Heusch G, Ovize M and Van de Werf F. Evolving therapies for myocardial ischemia/ reperfusion injury. J Am Coll Cardiol 2015; 65: 1454-1471.
[4] Panwar B, Arora A and Raghava GP. Prediction
and classification of ncRNAs using structural
[5] Greco S, Gorospe M and Martelli F. Noncoding RNA in age-related cardiovascular diseases. J Mol Cell Cardiol 2015; 83: 142-155.
[6] Ponting CP, Oliver PL and Reik W. Evolution and functions of long noncoding RNAs. Cell 2009; 136: 629-641.
[7] Wang K and Chang H. Molecular mechanisms of long noncoding RNAs. Mol Cell 2011; 43: 904-914.
[8] Prensner JR and Chinnaiyan AM. The emer-gence of lncRNAs in cancer biology. Cancer Discov 2011; 1: 391-407.
[9] Wang N, Luo YH, Deng JC, Feng J and Wang X. LncRNAs and cardiovascular diseases. Sheng Li Ke Xue Jin Zhan 2014; 45: 172-176. [10] Wu T, Chen C, Yang L, Zhang M, Zhang X, Jia J,
Wang J, Fu Z, Cui X, Ji C, Guo X, Tong M, Chen
R, Hong Q and Chi X. Distinct lncRNA expres
-sion profiles in the prefrontal cortex of SD rats
after exposure to methylphenidate. Biomed Pharmacother 2015; 70: 239-247.
[11] Xiong F, Zhu ZY, Xiong C, Hua XQ, Shan XH,
Zhang Y and Gu N. Preparation, characteriza-tion of 2-deoxy-D-glucose funccharacteriza-tionalized dimer-captosuccinic acid-coated maghemite nano- particles for targeting tumor cells. Pharm Res 2012; 29: 1087-1097.
[12] Hirsch LR, Stafford RJ, Bankson JA, Sershen SR, Rivera B, Price RE, Hazle JD, Halas NJ and West JL. Nanoshell-mediated near-infrared thermal therapy of tumors under magnetic resonance guidance. Proc Natl Acad Sci U S A 2003; 100: 13549-13554.
[13] Han J, Kim B, Shin JY, Ryu S, Noh M, Woo J, Park JS, Lee Y, Lee N, Hyeon T, Choi D and Kim BS. Iron oxide nanoparticle-mediated develop-ment of cellular gap junction crosstalk to im-prove mesenchymal stem cells’ therapeutic
efficacy for myocardial infarction. ACS Nano
2015; 9: 2805-2819.
[14] Xiong F, Wang H, Feng Y, Li Y, Hua X, Pang X, Zhang S, Song L, Zhang Y and Gu N. Cardiopro-tective activity of iron oxide nanoparticles. Sci Rep 2015; 5: 8579.
[15] Zhang S, Chen X, Gu C, Zhang Y, Xu J, Bian Z, Yang D and Gu N. The effect of iron oxide mag-netic nanoparticles on smooth muscle cells. Nanoscale Res Lett 2009; 4: 70-77.
[16] Chen B, Li Y, Zhang X, Liu F, Liu Y, Ji M, Xiong F
and Gu N. An efficient synthesis of ferumoxytol induced by alternating-current magnetic field.
Materials Letters 2016; 170: 93-96.
[17] Pei YH, Chen J, Xie L, Cai XM, Yang RH, Wang X and Gong JB. Hydroxytyrosol protects against myocardial ischemia/reperfusion injury th- rough a PI3K/Akt-dependent mechanism.
Me-diators Inflamm 2016; 2016: 1232103.
[18] Trapnell C, Pachter L and Salzberg SL. TopHat: discovering splice junctions with RNA-Seq. Bio-informatics 2009; 25: 1105-1111.
[19] Trapnell C, Williams BA, Pertea G, Mortazavi A, Kwan G, van Baren MJ, Salzberg SL, Wold BJ and Pachter L. Transcript assembly and
quan-tification by RNA-Seq reveals unannotated
transcripts and isoform switching during cell differentiation. Nat Biotechnol 2010; 28: 511-515.
[20] Benjamini Y and Hochberg Y. Controlling the false discovery rate-a practical and powerful approach to multiple testing. Journal of the Royal Statistical Society 1995; 57: 289-300. [21] Benjamini Y and Yekutieli D. The control of the
false discovery rate in multiple testing under dependency. Ann Stat 2001; 29: 1165-1188. [22] Kong L, Zhang Y, Ye ZQ, Liu XQ, Zhao SQ, Wei L
and Gao G. CPC: assess the protein-coding po-tential of transcripts using sequence features and support vector machine. Nucleic Acids Res 2007; 35: W345-9.
[23] Sun L, Luo H, Bu D, Zhao G, Yu K, Zhang C, Liu Y, Chen R and Zhao Y. Utilizing sequence intrin-sic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res 2013; 41: e166.
[24] Sun L, Zhang Z, Bailey TL, Perkins AC, Tallack MR, Xu Z and Liu H. Prediction of novel long non-coding RNAs based on RNA-Seq data of mouse Klf1 knockout study. BMC Bioinformat-ics 2012; 13: 331.
[25] Tafer H and Hofacker IL. RNAplex: a fast tool for RNA-RNA interaction search. Bioinformat-ics 2008; 24: 2657-2663.
[26] Tsompos C, Panoulis C, Toutouzas K, Zografos G and Papalois A. The effect of erythropoietin on progesterone levels during ischemia reper-fusion injury in rats. Actual Osteol 2014; 6: 65-70.
[27] Castillo RL, Arias C and Farías JG. Omega 3 chronic supplementation attenuates myocar-dial ischaemia-reperfusion injury through rein-forcement of antioxidant defense system in rats. Cell Biochem Funct 2014; 32: 274-281. [28] Angelone T, Filice E, Pasqua T, Amodio N,
Gal-luccio M, Montesanti G, Quintieri AM and Cerra
MC. Nesfatin-1 as a novel cardiac peptide:
identification, functional characterization, and
protection against ischemia/reperfusion inju-ry. Cell Mol Life Sci 2013; 70: 495-509. [29] Douglas SJ, Davis SS and Illum L.
Nanoparti-cles in drug delivery. Crit Rev Ther Drug Carrier Syst 1987; 3: 233-261.
[31] Nelson DM, Hashizume R, Yoshizumi T, Blakney AK, Ma Z and Wagner WR. Intramyocardial in-jection of a synthetic hydrogel with delivery of bFGF and IGF1 in a rat model of ischemic car-diomyopathy. Biomacromolecules 2014; 15: 1-11.
[32] Iovanna J, Orelle B, Keim V and Dagorn JC. Messenger RNA sequence and expression of rat pancreatitis-associated protein, a lectin-re-lated protein overexpressed during acute ex-perimental pancreatitis. J Biol Chem 1991; 266: 24664-24669.
[33] Lai Y, Li D, Li C, Muehleisen B, Radek KA, Park
HJ, Jiang Z, Li Z, Lei H, Quan Y, Zhang T, Wu Y,
Kotol P, Morizane S, Hata TR, Iwatsuki K, Tang C and Gallo RL. The antimicrobial protein RE-G3A regulates keratinocyte proliferation and differentiation after skin injury. Immunity 2012; 37: 74-84.
[34] Nishimune H, Vasseur S, Wiese S, Birling MC, Holtmann B, Sendtner M, Iovanna JL and Hen-derson CE. Reg-2 is a motoneuron neurotroph-ic factor and a signalling intermediate in the CNTF survival pathway. Nat Cell Biol 2000; 2: 906-914.
[35] Lorchner H, Poling J, Gajawada P, Hou Y, Polyakova V, Kostin S, Adrian-Segarra JM, Boettger T, Wietelmann A, Warnecke H, Richter M, Kubin T and Braun T. Myocardial healing re-quires Reg3beta-dependent accumulation of macrophages in the ischemic heart. Nat Med 2015; 21: 353-362.
[36] Zou L, Tu G, Xie W, Wen S, Xie Q, Liu S, Li G, Gao Y, Xu H, Wang S, Xue Y, Wu B, Lv Q, Ying M,
Zhang X and Liang S. LncRNA NONRATT021972 involved the pathophysiologic processes medi-ated by P2X7 receptors in stellate ganglia after myocardial ischemic injury. Purinergic Signal 2016; 12: 127-137.
[37] Kim J, Kim KY, Jang HS, Yoshida T, Tsuchiya K, Nitta K, Park JW, Bonventre JV and Park KM. Role of cytosolic NADP+-dependent isocitrate dehydrogenase in ischemia-reperfusion injury in mouse kidney. Am J Physiol Renal Physiol 2009; 296: F622-633.