_________________________________________________________ Journal of Experimental Biology and Agricultural Sciences KEYWORDS
Asian rice gall midge
Biotypes
Gene specific markers
Novel gene
ABSTRACT
Asian rice gall midge (Orseolia oryzae) is one of the major pests of rice which is causing significant economic loss to the crop. So far 11 gall midge resistance genes have been characterized from different rice varieties and seven biotypes of the pest have been identified based on their reaction to these genes. Cultivation of varieties containing single resistance gene has resulted in frequent breakdown of resistance due to emergence of virulent biotypes of the insect. Pyramiding of two or more effective genes in a single cultivar may lead to durable gall midge resistance. In present study about 100 gall midge resistant rice genotypes were screened under field condition against gall midge biotype GMB4M at Warangal and GMB1 in greenhouse at DRR. Furthermore, these entries were genotyped by using five gene specific markers which can detect the presence of Gm1, gm3, Gm4, Gm8 or Gm11 genes with high degree of success. Based on both phenotypic and genotypic analysis, these genotypes were gr ouped into five phenotypic sets. In Set 1, 17 genotypes showed nil damage at both the locations and amplified one or multiple gene specific alleles. In Set 2, 31 genotypes showed low level of damage (< 20%) at DRR and or Warangal, of which two breeding lines and a land race are likely carry new gene(s). In set 3, 18 genotypes displayed resistance against GMB1 but were susceptible against GMB4M. Set 4 had 6 genotypes that were found resistant against GMB4M but recorded up to 50% plant damage in greenhouse against GMB1. Set 5 had 22 genotypes that were susceptible against both the biotypes and not carried R genes.
Dutta S Suvendhu
1,2,#, Divya D
1,#, Durga Rani Ch V
2, Dayakar Reddy T
3, Visalakshmi V
4, Cheralu
C
5, Ibohal Singh Kh
6and Bentur J S
1,7,*1
Directorate of Rice Research, Rajendranagar, Hyderabad-500030, AP, India 2
Institute of Biotechnology and 3Department of Genetics and Plant Breeding, Acharya NG Ranga Agricultural University, Hyderabad-500030.AP, India 4
Agricultural Research Station, Ragolu-532484, AP, India 5
Regional Agricultural Research Station, Warangal-506007, AP, India 6
Central Agricultural University, Imphal-795004, Manipur, India 7
Present address: Agri Biotech Foundation, Rajendranagar, Hyderabad 500030, India
Received – July 25, 2014; Revision – August 12, 2014, Accepted – August 23, 2014 Available Online – August 30, 2014
CHARACTERIZATION OF GALL MIDGE RESISTANT RICE GENOTYPES USING
RESISTANCE GENE SPECIFIC MARKERS
E-mail: [email protected] (Bentur J S)
Peer review under responsibility of Journal of Experimental Biology and Agricultural Sciences.
* Corresponding author (# Both the authors as contributed equally)
Journal of Experimental Biology and Agricultural Sciences, August - 2014; Volume – 2(4)
Journal of Experimental Biology and Agricultural Sciences
http://www.jebas.org
ISSN No. 2320 – 8694
1 Introduction
The Asian gall midge, Orseolia oryzae (Wood-Mason) is one of the serious insect pests of rice responsible for causing an estimated annual yield loss worth more than US$700 million (Herdt, 1991). In India gall midge damage causes an average annual yield loss of about 477 thousand tons of grain or 0.8% of the total production amounting to US$ 80 million (Bentur et al., 2003). Breeding of resistant rice varieties has proved to be a viable and ecologically acceptable approach for management of this pest. So far 11 gall midge resistance genes and seven biotypes of gall midge have been identified. Cultivation of varieties containing a single resistance gene has resulted in frequent breakdown of resistance due to emergence of virulent biotypes of the insect across many regions in India (Bentur et al., 1987; Nair & Devi, 1994;Vijaya Lakshmi et al., 2006). It is thus imperative to find novel genes and deploy them effectively. Pyramiding of two or more effective genes in a single cultivar may lead to durable gall midge resistance. Use of molecular markers for development of gene pyramids in desired combination is being now followed. Molecular breeding using DNA markers for selection of resistant plants for gene pyramiding is now a proven technology (Sundram et al., 2008). These markers have also been used in fine mapping, identification of candidate gene and cloning of the target gene (Sharma et al., 2005). R gene specific markers have also been used for allele mining to identify better alleles at the same locus (Ramkumar et al., 2011). However, use of markers in gene discovery is relatively new (Himabindu, 2009). Several mapping studies have been undertaken using molecular markers to find the physical position of the gall midge resistance genes on the rice genome. Gm1 is mapped on chromosome 9 between the SSR markers RM23956 and RM23941 (Sundaram, 2007). Gm2 is mapped on chromosome 4 between the SSR markers RM17473 and RM17503 (Sundaram, 2007) while gm3 on chromosome 4 between the SSR makers RM17480 and gm3SSR4 (Sama, 2011). Gm4 is localized on chromosome 8 between the SSR markers RM22551 and RM22562 (Himabindu, 2009). Gm5 is tagged using RAPD markers OPR-19, OPR-09, OPB-14, OPQ-05 and OPE-01 but not mapped (Katiyar et al., 2000). Gm6 is reported on chromosome 4 between the AFLP markers RG476 and RG214 (Katiyar et al., 2001). Gm7 has been mapped on chromosome 4 using AFLP marker SA598 (Sardesai et al., 2002). Gm8 has been recently fine mapped on chromosome 8 with RM22685 and RM22709 (Sama et al., 2012) and Gm11 on chromosome 12 between the SSR markers RM28574 and RM28706 (Himabindu et al., 2010). Based on fine mapping of the R gene, candidate genes have been identified through in silico analysis of the rice genome within the marker delimited region (Yasala et al., 2012). A LRR gene is suspected to be the candidate gene at Gm4 locus on Chromosome 8 (Himabindu, 2009). Based on the sequence analysis of a polymorphic fragment of the gene amplified by a designed primer pair, a functional marker has been developed that showed no segregation with the trait in the mapping population. Likewise, the marker gm3del3 designed for the candidate gene NB-ARC
for gm3 gene located on the chromosome 4 (Sama, 2011; Sama et al., 2014) completely co-segregated with the trait in the mapping population of 300 F10 RILs. Yasala et al. (2012)
identified the Proline Rich Protein encoding gene as the candidate gene for Gm8 located on chromosome 8 and designed markers targeting the gene. All the three candidate genes have been functionally validated through Real-Time reverse transcription PCR (Divya et al., 2013). These three gene based markers have shown high degree of confidence in detecting the presence of gene in alternative mapping populations and are being used in MAS for gene introgression. In the present paper, these gene based markers for detecting the presence of the respective genes in a pool of about 100 rice genotypes has been used, most of which have been identified to be resistant against gall midge biotype GMB4 and identified new sources of reported genes and genotypes possibly carrying new resistance genes against the Asian rice gall midge.
2 Materials and methods
2.1 Plant material
A set of 77 gall midge resistant genotypes was collected from ARS, Ragolu, Andhra Pradesh (AP), India. These donor lines have been showing resistance with nil damage through successive years of testing and seed multiplication under natural pest infestation at this pest endemic location. Along with these, 20 breeding lines nominated under national gall midge testing program and three lines of the susceptible check TN1 were also included.
2.2 Phenotyping
The test entries were screened against gall midge biotype GMB1 in greenhouse at DRR, Hyderabad as per the standard protocols (Vijaya Lakshmi et al., 2006). Plants of test entries along with those of the standard susceptible check TN1 were raised in lines in plastic trays (60 x 30 x 30 cm) filled with puddled soil enriched with fertilizers. When plants were 10-15 days old, the tray was covered with a nylon mesh and 25 female + 10 male adult gall flies were released on covered crop for two consecutive days for infestation. Trays were transferred to a high humidity chamber (>90%RH) for the next two days for egg hatching and maggot establishment. Later, the trays were moved to ambient conditions and maintained till the gall appeared after around 20 days after the adult release. Entries were scored for reaction in terms of percent plant damage (n ≥10). Two replications were maintained. Test was considered valid if the susceptible check showed more than 90% plant damage. Plants without galls were dissected to observe dead maggots and/or for presence of tissue necrosis as manifestation of hypersensitive reaction (HR) to confirm these as resistant. Test entries with nil damage and up to 20% plant damage were considered as resistant while others were grouped as susceptible.
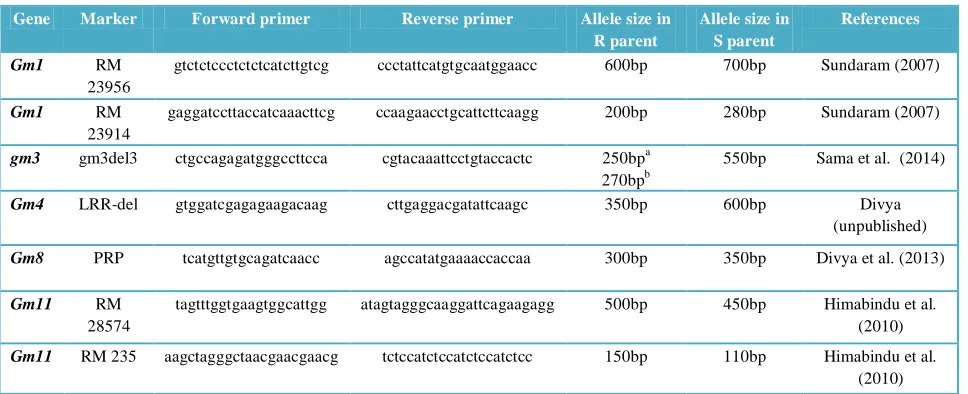
Table 1 Details of gene specific markers used in characterizing rice genotypes.
“R” and “S” parent indicates resistant and susceptible respectively
a
In RP2068-18-3-5 with gm3
b
In Phalguna with Gm2
Another set of the entries was screened against GMB4M at Agricultural Research Station, Warangal, Andhra Pradesh, under field conditions. Test entries were raised as seedlings and then transplanted with 15 x 20 cm spacing between plants and rows. Each entry was planted in two rows each with 20 plants. All the local agronomic practices recommended for rice cultivation were followed except application of any insecticide throughout the crop growth. Plants were scored on 30 and 45 days after transplanting based on percent plant damage. Test entries with nil damage and up to 20% plant damage were considered as resistant while others were grouped as susceptible.
2.3 Genotyping
Plant genomic DNA was extracted from leaf tissue of the test genotypes, gene differentials and the susceptible check TN1 through the method of Zheng et al. (1991). The purity and concentration of isolated genomic DNA were estimated by agarose gel electrophoresis with standard sample (0.8% agarose) and for PCR the protocol of Chen et al. (1997) was followed. Markers used for PCR amplification of genomic DNA were linked and flanking SSR markers RM23956 and RM23914 for Gm1 (Sundaram, 2007), RM28574 and RM28706 for Gm11 (Himabindu et al., 2010) and candidate gene based markers gm3del3 for gm3 (Sama et al., 2014), LRR-del for Gm4 (designed for this study) and PRP for Gm8 (designed for this study) (Table 1). The PCR products were
analyzed by electrophoresis using an agarose gel. The gels were visualized in a UV gel documentation system (Alpha innotech, Cell Biosciences Company USA).
3 Results
3.1 Phenotypic screening
In DRR greenhouse screening TN1 check showed an average plant damage of 99.09 ± 0.90 % (n = 11), while at Warangal it showed an average plant damage of 63.34 ± 14.23 % (n = 3). Among 97 genotypes evaluated, 17 (Set 1) recorded nil damage at both locations. Set 2 had 31 genotypes that showed marginal damage (≤20%) at DRR and Warangal. Set 3 had 18 entries that were resistant with nil damage at DRR but were susceptible (>20% DP) at Warangal in field screening. Set 4 had 6 entries displaying resistance with nil damage at Warangal but were susceptible (>20%DP) at DRR in greenhouse screening. Set 5 had 22 entries showing susceptibility reaction at both the locations.
3.2 Validation of gene specific markers
Markers used in the study were revalidated in the gene differentials. Results showed that one of the Gm1 gene flanking SSR marker RM23956 amplified a band size of 600bp in donor source Kavya and 700bp in susceptible source TN1.
Similarly the other flanking SSR marker RM23914 amplified a band size of 200bp in Kavya and of 280bp in TN1 (Table 1). The gm3 gene specific marker gm3del3 amplified a band of
250bp in RP2068-18-3-5 and 550bp in TN1, and also showed a third allele (270bp) specific to Phalguna carrying Gm2 gene as reported earlier (figure 1) (Sama, 2011).
Gene Marker Forward primer Reverse primer Allele size in
R parent
Allele size in S parent
References
Gm1 RM
23956
gtctctccctctctcatcttgtcg ccctattcatgtgcaatggaacc 600bp 700bp Sundaram (2007)
Gm1 RM
23914
gaggatccttaccatcaaacttcg ccaagaacctgcattcttcaagg 200bp 280bp Sundaram (2007)
gm3 gm3del3 ctgccagagatgggccttcca cgtacaaattcctgtaccactc 250bpa 270bpb
550bp Sama et al. (2014)
Gm4 LRR-del gtggatcgagagaagacaag cttgaggacgatattcaagc 350bp 600bp Divya
(unpublished)
Gm8 PRP tcatgttgtgcagatcaacc agccatatgaaaaccaccaa 300bp 350bp Divya et al. (2013)
Gm11 RM
28574
tagtttggtgaagtggcattgg atagtagggcaaggattcagaagagg 500bp 450bp Himabindu et al. (2010)
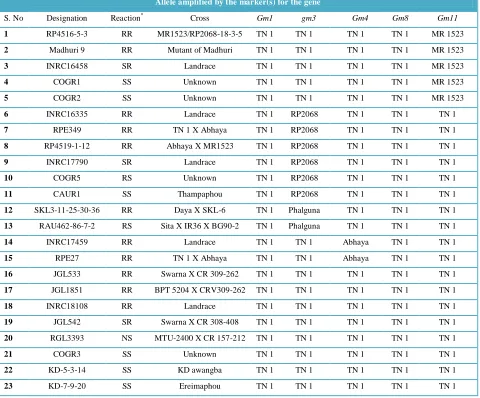
Table 2 Rice genotypes displaying single gene specific allele or none when amplified with R gene linked markers.
Allele amplified by the marker(s) for the gene
S. No Designation Reaction* Cross Gm1 gm3 Gm4 Gm8 Gm11
1 RP4516-5-3 RR MR1523/RP2068-18-3-5 TN 1 TN 1 TN 1 TN 1 MR 1523
2 Madhuri 9 RR Mutant of Madhuri TN 1 TN 1 TN 1 TN 1 MR 1523
3 INRC16458 SR Landrace TN 1 TN 1 TN 1 TN 1 MR 1523
4 COGR1 SS Unknown TN 1 TN 1 TN 1 TN 1 MR 1523
5 COGR2 SS Unknown TN 1 TN 1 TN 1 TN 1 MR 1523
6 INRC16335 RR Landrace TN 1 RP2068 TN 1 TN 1 TN 1
7 RPE349 RR TN 1 X Abhaya TN 1 RP2068 TN 1 TN 1 TN 1
8 RP4519-1-12 RR Abhaya X MR1523 TN 1 RP2068 TN 1 TN 1 TN 1
9 INRC17790 SR Landrace TN 1 RP2068 TN 1 TN 1 TN 1
10 COGR5 RS Unknown TN 1 RP2068 TN 1 TN 1 TN 1
11 CAUR1 SS Thampaphou TN 1 RP2068 TN 1 TN 1 TN 1
12 SKL3-11-25-30-36 RR Daya X SKL-6 TN 1 Phalguna TN 1 TN 1 TN 1
13 RAU462-86-7-2 RS Sita X IR36 X BG90-2 TN 1 Phalguna TN 1 TN 1 TN 1
14 INRC17459 RR Landrace TN 1 TN 1 Abhaya TN 1 TN 1
15 RPE27 RR TN 1 X Abhaya TN 1 TN 1 Abhaya TN 1 TN 1
16 JGL533 RR Swarna X CR 309-262 TN 1 TN 1 TN 1 TN 1 TN 1
17 JGL1851 RR BPT 5204 X CRV309-262 TN 1 TN 1 TN 1 TN 1 TN 1
18 INRC18108 RR Landrace TN 1 TN 1 TN 1 TN 1 TN 1
19 JGL542 SR Swarna X CR 308-408 TN 1 TN 1 TN 1 TN 1 TN 1
20 RGL3393 NS MTU-2400 X CR 157-212 TN 1 TN 1 TN 1 TN 1 TN 1
21 COGR3 SS Unknown TN 1 TN 1 TN 1 TN 1 TN 1
22 KD-5-3-14 SS KD awangba TN 1 TN 1 TN 1 TN 1 TN 1
23 KD-7-9-20 SS Ereimaphou TN 1 TN 1 TN 1 TN 1 TN 1
* Reaction against GMB1 in greenhouse reaction at DRR and against GMB4M in field test at Warangal. N-Not tested, R- resistant (<20% plant damage); S- susceptible (>20% plant damage)
Figure 1 Amplification pattern of selected genotypes with gm3del3 marker for presence of Gm2 and gm3 genes L- 100bp ladder, T- TN1, R- RP2068-18-3-5, P- Phalguna,1-8 test genotypes: 1. INRC16335, 2. JGL533, 3. RPE349, 4. Tellagarikasannavari, 5. JGL245, 6.
Figure 2 Amplification pattern with gm3del3, LRR-del, PRP functional markers in susceptible TN1 and resistant RP2068, Abhaya and Aganni for the presence of gm3, Gm4 and Gm8 genes. L- 100bp ladder.
The Gm4 gene specific reported marker LRR (Himabindu, 2009) was refined to develop LRR-del marker which amplified a band size of 350bp in the Gm4 donor source Abhaya and 600bp in susceptible source TN1(figure. 2). The Gm8 gene specific marker PRP developed from the sequence information of the reported candidate gene Proline Rich Protein 3 putative (Yasala et al., 2012) amplified a band size of 300bp in gene source Aganni and 350bp in TN1. One of the flanking SSR markers, RM28574 for Gm11 amplified a band size of 500bp in MR1523 and 450bp in TN1. Other flanking SSR marker RM235 amplified a band size of 150bp in MR1523 and 110bp in TN1.
3.3 Genotypic screening
Genotyping data of the test entries in respect of three candidate gene based markers and two pairs of flanking markers for the six gall midge R genes revealed presence of single or multiple R genes in several of the test lines. Gm1 flanking markers detected the presence of gene in only 6 test entries, Another set of flanking SSR markers for Gm11 gene identified the presence of the gene in 37 test entries, of which five (RP 4516-5-3, COGR-1, COGR-2, Madhuri L9, INRC16458) recorded presence of this gene only (Table 2). The gm3del3 functional marker detected gm3 gene in 43 of the test entries while six of these entries (INRC16335, INRC17790, CAUR-1, COGR-5, RPE-349, RP4519-1-12) displayed only gm3 specific allele. This marker also detected Gm2 allele in 14 test genotypes but only two (RAU 462-86-7-2, SKL3-11-25-30-36) had this gene alone. Interestingly, the marker amplified a fourth unknown allele in two test lines i.e. S2207 and Tellagarikasannavari. The newly designed LRR-del marker for Gm4 detected the gene in 42 test entries. Of these, INRC17459 and RPE-27 carried only Gm4 gene. PRP marker developed for Gm8 gene confirmed its presence in 15 test entries but none had only this gene. Among the test entries 8 did not amplify gene specific alleles with any of the markers used. These displayed susceptibility or TN1 allele. Three entries did not amplify any allele with all the markers tested.
3.4 Genotype characterization
Based on phenotypic sets and presence of single or multiple R genes, the test genotypes were characterized. Among the first set of 17 genotypes recording nil gall midge damage at both Warangal and DRR, against gall midge biotypes GMB4M and
GMB1, respectively, eight entries carried one of the R genes viz., gm3 (n=3), Gm4 (3) or Gm11 (2). Rest of the nine entries carried combination of these genes. Three of the latter also carried Gm11 gene. None of the entries, probably, carried novel resistance gene.
Among the 31 test entries in set 2 that displayed low level of damage at both the test locations, three did not amplify any known resistance gene specific allele with the markers. They carried TN1 allele for all the markers. Since the damage level still qualified these entries to be resistant and hence these entries may likely to carry novel resistance gene(s). Two of the entries (JGL533, JGL1851) are breeding lines while the other one is a land race. Information regarding the one entry was not traceable. Other test entries carried gm3, Gm4, Gm8 or Gm11 alone or in combination.
Of the 18 entries in set 3 displaying resistance at DRR and susceptibility at Warangal, four entries carried either Gm2 alone or combinations of Gm1 Gm2 and Gm11 genes. These genes do not confer resistance against GMB4M prevalent in Warangal. The set 4 with six test entries displayed resistance at Warangal and carried combination of effective genes like gm3, Gm4 and Gm8. However these entries recorded up to 50% plant damage in greenhouse test at DRR. We attribute susceptibility in these entries at DRR to likely seed impurity. The last of the sets, set 5 with 22 test entries displayed susceptibility at both the test locations. Six of the entries did not carry any resistance gene tested. Five more entries carried Gm11 gene alone. Rest of the entries carried combination of genes.
4 Discussions
RM23914 encompass 0.19Mb region of the rice genome on chromosome 9 and are at the genetic distance of 1.0 and 1.1 cM from the gene. Thus the probability of detecting the presence of this allele using both the markers is 0.99. In present study this gene was detected in only 6 genotypes tested, of which the breeding line WGL31996 had Kavya as one of the parents. Landraces IC114788 and Tellagarikasannavari could be alternative sources of the gene. In silico analysis of the 0.5Mb region of the rice genome encompassing Gm1 gene revealed 62 putative genes, of which 15 were, annotated genes (Yasala et al., 2012). A set of genes reported to be part of plant defense mechanism against biotic stresses, including those mentioned above, was validated in Real-Time reverse transcription quantitative PCR (RT-PCR) assay in Kavya rice variety carrying Gm1 gene (Rawat et al., 2012a). But none of the genes showed signs of induction or repression in plants infested by gall midge as compared with the uninfested plants, which was ascribed to a novel mechanism of constitutive gall midge resistance in Kavya conferred by Gm1 gene. This is delaying our attempt to develop a functional marker for the gene.
Himabindu et al. (2010) mapped the new resistance gene Gm11 using 466 F10 RIL mapping population derived from the
cross TN1 / MR1523. The closest flanking reported markers RM28574 and RM28706 covered 1.7Mb genome of chromosome 12 encompassing the gene and separated from it by genetic distance of 4.4 and 3.8 cM. Thus the probability of detecting the presence of the gene with the flanking markers was only 0.83. This gene was detected in 37 genotypes, many of which were derived from the gene source CR57-MR1523. The flanking marker delimited region of the rice genome contained 134 annotated genes, of which one coded for NBS-LRR class of resistance protein (Yasala et al., 2012). Investigating the molecular basis of Gm11 conferred gall midge resistance in the rice variety Suraksha through analysis of suppression subtraction hybridization (SSH) cDNA libraries and RT-PCR analysis of selected candidate genes (Rawat et al., 2010; Rawat et al., 2012b) identified six genes within the marker delimited region. However, none of these genes showed length polymorphism when amplified with sets of designed primer pairs in the contrasting parents. Development of a functional marker for the gene is on hold since wide spread virulence in gall midge populations against Gm11 conferred resistance is limiting the utility of the gene in pest management.
A gene encoding NB-ARC class of resistance protein was identified as the candidate gene at gm3 locus and it was functionally validated through RT-PCR studies (Sama, 2011; Sama et al., 2014). Based on the sequence information of the gene marker gm3del3 was developed which distinguished the gene source RP2068-18-3-5 from the susceptible TN1 rice variety. It also distinguished Gm2 gene carrying Phalguna cultivar from these two genotypes. In the present test, the marker suggested presence of gm3 gene in 43 of the test entries including two land races INRC16335, INRC17790 that may be alternative sources of the gene. It also identified Gm2 gene in
14 of the genotypes. Further, a fourth allele was amplified by the marker in two of the test entries S2207 and Tellagarikasannaari. Unexpected detection of the allele in Abhaya with Gm4 gene and several of its derived lines needs further studies to know if the detected allele is indeed functional.
Attempts to fine map Gm4 gene in two alternative populations consisting of 233 progeny tested F2:3 plants from ISM / Abhaya
cross and 91 F10 RILs of TN1 / Abhaya cross led to in silico
analysis of 0.2Mb genomic region on chromosome 8 delimited by the flanking markers RM547 and RM22562 (Himabindu, 2009). Of the 70 genes in the region, two coded for LRR proteins known to be involved in plant resistance to pathogens and pests.
One of the marker LRR designed to amplify fragments of these genes from the parents showed polymorphism suggesting this gene (LOC_Os08g09670.1) to be the candidate gene for Gm4 locus. Subsequently this gene was validated through RT-PCR studies. In the present study, further improvement in this marker and designed it as LRR-del, targeted it more closer to the deletion site for greater resolution of the alleles. This marker detected the Gm4 gene in 42 genotypes tested, including 15 of the 21 breeding lines derived from Abhaya. One of the land races tested INRC17459 could be alternative source of this gene.
Sama et al. (2012) fine mapped Gm8 gene to chromosome 8 within a 400-kbp region. In silico analysis of the region revealed 49 functionally annotated genes (Yasala et al., 2012). Designing several sets of primers and testing these with the contrasting parents and their derived progeny led to narrowing down to the candidate gene that code for Proline Rich Protein. The gene has been validated through RT-PCR studies (Divya et al., 2013). The marker-PRP identified the gene in 15 genotypes tested, including INRC3021 earlier confirmed to have Gm8 gene (Sama et al., 2012).
Acknowledgements
We thank the Project Director, Directorate of Rice Research, Hyderabad for the facilities and encouragement. This work was supported by a grant from the Department of Biotechnology (F.No: BT/AB/FG-2 (PH-II)/2009), Government of India. SSD is thankful to ICAR for Fellowship and The Director of Institute of Biotechnology, ANGRAU, Hyderabad for his support and transcendent suggestions.
References
Bentur JS, Srinivasan TE, Kalode MB (1987) Occurrence of a virulent rice gall midge (GM) Orseolia oryzae Wood-Mason biotype (?) in Andhra Pradesh, India. International Rice Research Newsletter 12: 33-34.
Bentur JS, Pasalu IC, Sarma NP, Rao UP, Mishra B (2003) Gall midge resistance in rice. DRR Research paper Series 01/2003. Directorate of Rice Research, Hyderabad, India, pp. 20.
Chen X, Temnykh S, Xu Y, Cho YG, McCouch SR (1997) Development of a microsatellite frame work map providing genome wide coverage in rice (Oryza sativa L.). Theoretical and Applied Genetics 95: 553-567.
Divya D, Bentur JS, Nair S (2013) Identification of putative candidate gene (s) for gall midge resistance Gm8 gene in Aganni Rice. In: Abstracts: National symposium on Innovative Approaches to Crop Improvement and adaptation: Meeting Challenges of Climate Change, 22-24 Febuary. UAS, Bangalore, p. 75.
Herdt RW (1991) Research priorities for rice biotechnology. In: Khush GS, Toenniessen GH (eds) Rice biotechnology. CABI and International Rice Research Institute, Manila, Philippines, pp. 19-54.
Himabindu K, Vijaya Lakshmi P, Sundaram RM, Neeraja CN, Mishra B, Bentur JS (2007) Flanking SSR markers for allelism test for the Asian rice gall midge (Orseolia oryzae) resistance genes. Euphytica 157: 267-279.
Himabindu K (2009) Identification, tagging and mapping of rice gall midge resistance genes using microsatellite markers. PhD thesis submitted to Acharya Nagarjuna University, Guntur, pp. 241.
Himabindu K, Suneetha K, Sama VSAK, Bentur JS (2010) A new rice gall midge resistance gene in the breeding line CR57-MR1523, mapping with flanking markers and development of NILs. Euphytica 174: 179-187.
Kalode MB, Bentur JS, Prasad Rao U (1993) Rice cultivars resistant to rice gall midge (Gm) biotypes 1 and 4 under
artificial infestation in green house. International rice research newsletter 18: 17-18.
Katiyar SK, Verulkar SB, Adsul G, Dhundre M, Chandel G, Bennett J (2000) Molecular markers for gall midge resistance genes in rice: Stage set for MAS and map based gene cloning. In: Abstracts: IV rice genetics symposium, 22-27 October. International Rice Research Institute, Los Banos, Philippines, p. 81.
Katiyar SK, Tan Y, Huang B, Chandel G, Xu Y, Zhang Y, Xie Z, Bennett J (2001) Molecular mapping of gene Gm-6(t) which confers resistance against four Biotypes of Asian rice gall midge in China. Theoretical and Applied Genetics 103: 953-961.
Nair KVP, Devi A (1994) Gall midge biotype 5 identified in Moncompu, Kerala, India. International Rice Research Notes 19: 11.
Ramkumar G, Srinivasarao K, MadanMohan K, Sudershan I, Sivaranjani AKP, Gopalkrishana K, Viraktamath BC, Madhav MS (2011) Development and validation of functional marker targeting an InDel in the major rice blast disease resistance gene Pi54 (Pikh). Molecular Breeding 27: 129-135.
Rawat N, Sinha DK, Rajendrakumar P, Shrivastava P, Neeraja CN, Sundaram RM, Suresh Nair, Bentur JS (2010) Role of pathogenesis-related genes in rice gall midge interactions. Current Science 99: 1361-1368.
Rawat N, Neeraja CN, Sundaram RM, Nair S, Bentur JS (2012a) A novel mechanism of gall midge resistance in the rice variety Kavya revealed by microarray analysis. Functional and Integrative Genomics 12: 249-264.
Rawat N, Himabindu K, Neeraja CN, Nair S, Bentur JS (2012b) Suppressive subtraction hybridization reveals that rice gall midge attack elicits plant-pathogen like responses in rice. Plant Physiology and Biochemistry 63: 122-130.
Sama VSAK (2011) Identification, tagging and mapping of new resistance gene(s) against the Asian rice gall midge Orseolia oryzae in rice varieties. PhD thesis submitted to Osmania University, Hyderabad, pp.161.
Sama VSAK, Himabindu K, Naik SB, Sundaram RM, Viraktamath BC, Bentur JS (2012) Mapping and MAS breeding of an allelic gene to the Gm8 for resistance to Asian rice gall midge. Euphytica 187: 393-400.
Sardesai N, Kumar A, Rajyashri KR, Nair S, Mohan M (2002) Identification and mapping of an AFLP marker linked to Gm7, a gall midge resistance gene and its conversion to a SCAR marker for its utility in marker aided selection in rice. Theoretical and Applied Genetics 105: 691-698.
Sharma TR, Madhav MS, Singh BK, Shanker P, Jana TK, Dalal V, Pandit A, Singh A, Gaikwad K, Upreti HC, Singh NK (2005) High-resolution mapping, cloning and molecular characterization of the Pi-kh gene of rice, which confers resistance to Magnaporthe grisea. Molecular Genetics and Genomics 274: 569–578.
Sundaram RM (2007) Fine mapping of rice gall midge resistance genes Gm1 and Gm2 and validation of the linked markers. PhD thesis submitted to University of Hyderabad, Hyderabad, pp.181.
Sundaram RM, Vishnupriya MR, Biradar SK, Laha GS, Ashok Reddy G, Shoba Rani N, Sarma NP, Sonti RV (2008) Marker assisted introgression of bacterial blight resistance in Samba Mahsuri, an elite indica rice variety. Euphytica 160: 411-422.
Vijaya Lakshmi P, Amudhan S, Himabindu K, Cheralu C, Bentur JS (2006) A new biotype of the Asian rice gall midge Orseolia oryzae (Diptera: Cecidomyiidae) characterized from the Warangal population in Andhra Pradesh, India. International Journal of Tropical Insect Science 26: 207-211.
Yasala AK, Rawat N, Sama VSAK, Himabindu K, Sundaram RM, Bentur JS (2012) In silico analysis for gene content in rice genomic regions mapped for the gall midge resistance genes. Plant Omics Journal 5: 405-413.
Zheng KL, Shen B, Qian HR (1991) DNA polymorphism generated by arbitrary primed PCR in rice. Rice Genet Newsletter 8: 134-136.