JOURNN OF
IMNGE MhNFGEMENT
Published bimonthly-January, March, May,July, September, November-by the
TABLE OF CONTENTS:
Vol. 31, No. 5, September 1978
Society for Range Management 2760 West Fifth Avenue
ARTICLES 134 Denver, Colorado 80204
Copyright @ 1978 by the Society for Range Management Managing Editor
DAVID A. SMITH 2760 West Fifth Avenue Denver, Colo. 80204
“IT Symposium on Poisonous Plants-Introduction by Lynn F. James
Editor REX D. PIEPER
Dep. Animal and Range Science New Mexico State Univeristy Las Cruces, N. Mex. 88003 Book Review Editor PAT 0. CURRIE
Rocky Mountain Forest and Range Experiment Station 240 West Prospect Street Fort Collins, Colo. 80521 Copy Editor
PATRICIA G. SMITH 2760 West Fifth Avenue Denver, Colo. 80204 Editorial Board 1976-78
RAYMOND EVANS, Reno, Nev. CLIFFORD E. LEWIS, Marianna, Fla. JOHN MALECHEK, Logan, Utah C. J. SCIFRES, College Station, Tex. 1977-79
JAMES 0. KLEMMEDSON, Tucson, Ariz. GEORGE W. SCOTTER, Edmonton, Alta. M. JOE TRLICA, Fort Collins, Colo. J. ROSS WIGHT, Logan, Utah 1978-80
DONALD A. KLEBENOW, Reno, Nev. JAMES T. NICHOLS, North Platte, Neb. MICHAEL D. PITT, Vancouver, B.C. RONALD E. SOSEBEE, Lubbock, Tex.
INDIVIDUAL SUBSCRIPTION is by membership in the Society for Range Management.
LIBRARY or other INSTITUTIONAL SUBSCRIPTIONS, on a calendar year basis, are $22.00 postpaid to all countries. Payment from outside the United States should be remitted in US dollars by international money order or draft on a New York bank.
CHANGE OF ADDRESS notices should be sent to the Managing Editor, 2760 West Fifth Ave., Denver, CO 80204, no later than the first day of the month of issue. Copies lost due to a change of address cannot be replaced unless adequate notice is given. To assure uninterrupted service, provide your local postmaster with a Change of Address Order (POD Form 3575) indicating thereon to guarantee forwarding postage for second class mail. POSTOFFICE: Please return entire journal with address change. BUSINESS CORRESPONDENCE, concerning subscrip- trons, advertising, reprints, back issues, and related mat- ters, should be addressed to the Managing Editor, 2760 West Fifth Ave., Denver, CO 80204.
EDITORIAL CORRESPONDENCE, concerning manu- scripts or other editorial matters, should be addressed to the Editor, Dep. Animal and Range Sciences, New Mexico State Univ., Las Cruces 88003.
INSTRUCTIONS FOR AUTHORS appear each year In the March issue; copies of these instructions are available from the editor.
TECHNICAL NOTES
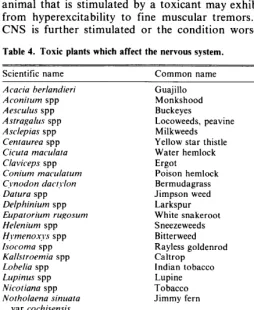
392 Artemisia vulgaris L.: An Ornamental Plant for Disturbed Land Reclamation by Gerard P. Schuman and Gene S. Howard
393 Measuring Fireweed Utilization by Edmund P. Harshman and Richard Forsman
3% Predicting Green Weight of Mesquite (Prosopis glandulosa Torr.) by Steven G. G. Whisenant and Donald F. Burzlaff
398 Control of Black Grass Bugs (Labops hesperius Uhler) in Northern New Mexico
by George W. Dickerson
BOOK REVIEWS
399 Creosote Bush: Biology and Chemistry of Larrea in New World Deserts edited by T. J. Mabry, J.H. Hunziker, andD.R. Difeo, Jr.;Mesquite, Its Biology in Two DesertShrub Ecosystems. edited by B.B. Simpson; Terrestial Vegetation of California edited by Michael G. Barbour and Jack Major.
325
328
335
343 347
351
355
361
366
371 375
378
381
386
The Economic Impact of Poisonous Plants on the Range Livestock Industry in the 17 Western States by Darwin B. Nielsen
The Ecological Niches of Poisonous Plants in Range Communities by E . H . Crmin,
Phil Ogden, James A. Young, and William Laycock
Coevolution of Poisonous Plants and Large Herbivores on Rangelands by W.A. Laycock
Physiologic Responses of Livestock to Toxic Plants by E. Murl Bailey, Jr.
Management Approaches to Reduce Livestock Losses from Poisonous Plants on Rangeland by William C. Krueger and Lee A. Sharp
. ,
Grazing Management Practices Affect Lifestock Losses from Poisonous Plants by Leo B. Merrill and Joseph L. Schuster
Reducing Incidence of Plant-Caused Congenital Deformities in Livestock by Grazing Management by Richard F. Keeler
Tiller Development and Growth in Switchgrass by E.R. Beaty , J. L. Engel, and John
D. Powell ._ I
High Rates of Nitrogen Change Composition of Shortgrass Rangeland in South- eastern Wyoming by Frank Rauzi
Germination of Goldenweed Seed by H. S . Mayeux, Jr., and C. J. Scifres
Herbaceous Vegetation Changes Following Application of Tebuthiuron for Brush Control by C.J. Scifres and J.L. Mutz
Global Desertification and Range Management: an Appraisal by E.G. Van Voorthuizen
Net Aerial Primary Production of an Andropogon-Paspalum Grassland Ecosystem
by C.M. Britton, J.D. Dodd, and A.T. Weichert
Acute Impact of Herbicide Strip Treatment on Mixed-Brush White-Tailed Deer Habitat on the Northern Rio Grande Plain by G. W. Tanner, J.M. Inglis, and L.H. Blankenship
.’
SECOND CLASS POSTAGE paid at Denver, Colorado.
Symposium on Poisonous Plants
Held at the Annual Meeting
Society for Range Management
U.S. Department
San Antonio, Texas
February 6-9, 1978
Lynn F. James-Chairman
of Agriculture Poisonous Plant Research Laboratory Logan, Utah
Introduction
Poisonous plants are one of the principal causes of economic loss to the livestock industry of the world. There is a general tendency to think of poisoning in terms of death. There are, however, many ways in addition to death that poisonous plants exert their detrimental effects on livestock. Some of these additional effects include chronic illness and debilitation, decreased weight gains, abortion, birth defects, and photosensitization. In addition to these more obvious losses, we must include increased costs associated with managing ranges and pastures infested with poisonous plants. Some of these less obvious but important costs include fencing, loss of forage, increased labor costs in herding and moving livestock, and in some cases, supplemental feeding.
A large portion of the earth’s land area is devoted to the grazing of livestock. The forage on these lands can best be harvested by grazing animals. Poisonous plants ofttimes interfere with the proper harvesting of this forage. Proper range management is important in the prevention of livestock poisoning by plants.
The purpose of this symposium is to bring together for discussion problems related to livestock poisoning by plants and management methods related to prevention.
Manuscript received March 30, 1978.
The Economic Impact of Poisonous Plants on
the Range Livestock Industry in the
17
Western States
DARWIN B. NIELSEN
HIGHLIGHT: Poisonous plants cause serious economic losses in many areas of the West. However, there is no systematic way of accounting for the magnitude of these losses. A significant proportion of the poisonous plant loss is reflected in annual death loss in livestock and in calf and lamb crop percentages. By concentrating one’s effort on the effect of poisonous plants on these measures, one should be in a better position to make reasonable estimates of the economic costs of poisonous plants. Other losses from poisonous plants should be considered as data become available. Based on the assumptions outlined, the econom- ic loss in the 17 western states is about $107 million annually. Poisonous plants have the potential on many ranches of causing financial ruin to the business. It has been shown that poisonous plants can be economically controlled and losses kept at manage- able levels.
The following statements typify efforts to assess the economic importance of poisonous plants to the livestock industry. “Poisonous plants cause great losses on the western range in death of animals and in decreased value, essentially through lesser gains” (Stoddart and Smith 1943). “Poisonous plants are a principal cause of economic loss to the livestock industry in the western United States” (Keeler et al. 1978). “Loss from p&sonous plants is one of the major economic problems in livestock production” (Sperry et al. undated). “Yearly livestock losses from plant poisoning in the U.S. amount to many millions of dollars”(Gilkey 1958). “Each year these plants exact a costly toll of the livestock of this state, often as much as 8 percent” (Durrell and Newson 1939).
Poisonous plants have been considered a serious problem on western ranges for many years yet there have been few serious attempts to quantify the magnitude of the losses. The reluctance is understandable since one is faced with several problems that seem insurmountable. First, many ranchers do not recognize poisonous plants on their rangelands nor is the task of identifying the cause of death on animals found on the range an easy one.
Estimating losses from veterinarian reports where at- tempts to save animals or find cause of loss are involved is not adequate. If the veterinarian can offer little aid to the rancher when called concerning a poisonous plant problem, the rancher is not going to call and pay a veterinarian the next time he has problems. What usually gets reported as poisonous plant losses are cases where the loss is severe, and/or it is an unusual case in a given range area.
The author is Professor, Department of Economics, Utah State University Logan 84322.
Manuscript received March 30, 1978.
Ranchers tend to figure a certain amount of poisonous plant loss in the average annual death loss in their herds. This idea is expressed in a study in New Mexico, “Livestock poisoning from certain native range plants is one of the hazards to ranching in New Mexico” (Norris and Valentine 1957). Thus, some level of death loss is considered a normal cost of doing business and is not given special attention unless unusually heavy losses occur. Therefore, some people contend that poisonous plant losses are part of the normal cost of doing business in a given ranching area and are reflected in the value of the properties, so why worry about them. However, ranching is not a high-profit business that can stand losses without concern about them. Undoubtedly there are cases where it would cost more to control poisonous plants than the value of animals saved would justify, but there are many more cases where loss prevention is profitable.
Losses from poisonous plants affect the economic returns to ranching in several ways. The most obvious is animals that are killed directly from the consumption of these plants. Death is the loss most often associated with poisonous plants and is probably the easiest loss to evaluate. Some poisonous plants do not kill the animal but severely reduce the production from that animal and/or its offspring. This loss would be reflected in weaning weights, percent calf or lamb crop, unmarketable animals due to birth deformities, and reduced longevity of the breeding herd. Probably the least obvious economic loss caused by poisonous plants on rangelands is that the presence of poisonous plants cause the range resource to be used and developed at less than its optimum. For example, the decision on what class of animal to graze (sheep or cattle) could be a result of trying to avoid poisonous plant problems rather than which class of animal is best suited to utilize the rangeland on the basis of available forage and topography. Poisonous plant problems often dictate the season of use of rangelands, thus not allowing these lands to be used as efficiently as they could without this potential danger.
Livestock and range management costs are often higher where poisonous plants are a problem. These costs include plant control costs, fencing, and more intensive manage- ment associated with the presence of poisonous plants.
It is much easier to speculate on the ways poisonous plants cause economic losses to the livestock industry than it is to estimate the magnitude of these losses. Given present knowledge on the subject, it is impossible to make objective estimates of the economic loss caused by poisonous plants. The fact that there is very little empirical data on aggregate estimates of the economic losses from poisonous plants
points to an area where some research effort can be expended.
Annual death loss statistics on beef cattle and sheep include animals that died from poisonous plants. Therefore, in the aggregate, poisonous plant death loss must be less than or equal to the total death loss. It is recognized that individual ranchers or ranching areas where poison plants are particularly serious could have losses higher than the overall average death loss to all causes. But one would find himself in a logically indefensible position if he used local figures on poison plant death loss and applied them to a state or group of states and found poison plant death loss higher than the total death loss to all causes. To avoid this trap, let us look at the reported death loss, to all causes, for adult cattle and sheep in the western states.
22,269,OOO (U.S. Dep. Agr. 1977). Each 1% death loss in these cows amounts to 222,690 head or $55,672,500, assum- ing $250 value per head. Suppose one decides that one third of the death loss is due to poisonous plants. In this case the estimated annual value of the death loss because of poison- ous plants would be 1% of the total number of cattle or $55,672,500.
U.S. Department of Agriculture (1973) reported annual adult cattle death loss at 2% in 1969. Kearl(1967) shows the following annual death losses in Wyoming: cows- 1.6%, l-2 year old heifers-3.0%, and bulls--5%. Gray (1970) esti- mates death losses on adult cattle in New Mexico at 2.2- 2.7ojo. Roberts and Gee (1963) report cattle death losses ranging from 3-7%. Gray and Baker (1953) reported death losses ovet the period 1930-52 in Montana; the average for cattle was 3.6%, the high during the period was 5.970, and the low was 2.4%. Myles (1963) found the average death loss on adult cattle for a 14-year period to be 3.3% in Nevada. Adult sheep losses are usually higher than adult cattle losses. Relatively low salvage value for cull ewes cause ranchers to run them longer than would be the case if they were worth more as culls. Goodsell and Belfield (1973) reported an average death loss on adult ewes at 6.5% for migratory sheep ranches in Utah and Nevada. U.S. Department of Agricul- ture (1977) reports the number of ewes 1 year and older on January 1 for each state; they also report an “estimate of adult sheep death losses.” Based on these figures for the 17 Western States, the average death loss is about 12%.
There were about 7,136,OOO head of ewes 1 year old and older in the Western States on January 1, 1977 (Table 2). Each 1% loss would amount to 7 1,360 ewes or a loss of $3,568,000, given the value of a breeding ewe at $50 per head. Since sheep usually spend more time on rangeland than cattle, one could assume the death loss from poison- ous plants to be higher. If the poisonous plant death loss is 3.5%, then the value of animals lost would be: $3,568,000 x 3.5 = $12,488,000.
Table 2. Number of sheep, 1 year * and older, as of January 1, 1977, for 17 Western States and estimated annual death loss of adult sheep, 1975.
State Number of ewes’ Estimated death loss2
Utah Idaho Wyoming Oregon Colorado Nevada New Mexico California Arizona Montana Washington Texas North Dakota South Dakota Nebraska Kansas Oklahoma
475,000 86,000 420,000 37,000 827,000 156,000 247,000 40,000 426.000 50,000 I 00,000 I 7,000 400,000 78,000 710,000 68,000 283,000 25,000 410.000 80,000 50,000 I 2,000 1,865,OOO I00,000 158,000 28,000 5 10.000 45,000 9 I ,000 22,000 I15,000 14,000
49,000 6,000
It appears from the above studies that the adult cattle death loss is somewhere around 3.0% and sheep losses are about S-10% annually. Again, it should be emphasized that the death loss can be much higher on an individual ranch or in an area where poisonous plants are particularly serious. The economic significance of this will be discussed in detail later. Poisonous-plant-caused deaths in adult cattle and sheep are part of the annual death loss. The exact propor- tion is not known, but an upper limit on the estimated loss has been set. The number of cows 2 years old and older, excluding dairy, for the 17 Western States is given in Table 1. The total number of cows as of January 1, 1977, is
TOTAL 7,136,OOO 864,000
1 U.S. Department of Agriculture. 1977. Breeding sheep, I year and older. January I, 1977. Crop Reporting Board, Statistical Reporting Service.
2 U.S. Department of Agriculture. 1975-76. Meat animals, production. disposition. income. Crop Reporting Board, Statistical Reporting Service.
If one accepts these estimates, the value of poison plant death loss is $68.2 million annually in the 17 Western States. One should remember that there is no way of estimating, with current information, an exact figure on death losses. However, one could consider them “ball park” figures with a limit on the size df the ball park.
Table 1. Number of cows (excluding dairy) 2 years old and older as of January 1, 1977, for 17 Western States.’
State Utah Idaho Wyoming Oregon Colorado Nevada New Mexico Arizona California
No. of cows State ( 1,000 head)
335 Montana 608 Washington 721 Texas 599 North Dakota 889 South Dakota 298 Nebraska 644 Kansas 319 Oklahoma 991
TOTAL
TOTAL = 22.269,OOO head
No. of cows ( I.000 head)
I.549 355 6,482 I.060 I.388 2,082 1,690 2,259 22,269
Many poisonous plants do not result in the death of animals but they reduce the performance of the animals. It has been stated by some that this loss could be higher than the actual death loss. Pine needle (Pinus spp.) abortion, locoweed (certain species of Astragalus and Oxytropis) caused abortions, and abortions plus birth deformities caused by several other poisonous plants have serious effects on calf and lamb crop production in many areas of the West.
James ( 1978) reports:
1 U.S. Department of Agriculture. 1977. Beef cattle numbers, January I. 1977. Crop Reporting Board. Statistical Reporting Service.
In recent years plant toxins have been recognized as important causes of reproductive problems. Certain plant toxins have been shown to cause birth defects, embryonic and fetal deaths, and abortions. The estrogen found in certain plants has adversely affected sexual activities and reproduction in some livestock. The locoweeds and plants of the
Astragalus and Ox,‘tropis genera depress spermatogenesis, oogenesis, and general sexual activity in sheep, cattle, and horses.
To be most useful in an economic sense, calf and lamb crop percentages should be calculated on the number of cows or ewes in the breeding herd to the number of calves and lambs marketed or weaned. Calf and lamb death loss up to market or weaning time would be figured in the calf or lamb crop percentage. Lamb and calf crop percentages figured this way are considerably lower than those reported by many U.S. Department of Agriculture statistics. We have been led to believe that there is not much that can be done to improve lamb and calf crop. There appears to be more slack in this measure of performance efficiency than one might suspect. Reduction of poisonous plant problems is one of the areas of research and management where improvements can be made in the reproduction efficiency of the range livestock industry.
The number of calves available for sale could be increased 222,690 head for each percentage point increase in calf crop and each 1% increase in lamb crop would provide an addi- tional 7 1,360 head of lambs for sale. The value of calves and lambs for each 1% calf and lamb crop would be as follows: follows:
Calves: 222,690 head x $160 = $35,630,400 Lambs: 71,360 head x $ 45 q $ 3,211,200
Poisonous plants could cause reductions in lamb and calf crop by affecting the reproductive capabilities of the breed- ing herd, and any lamb or calf death loss from birth to weaning. When a lamb or calf is lost, prenatal or postnatal, a rancher loses the opportunity to sell a calf or lamb at weaning. He has essentially incurred all the costs of producing the additional animals because he has to main- tain the breeding herd.
Let us assume calf and lamb crop could be increased 1% if we did not have poisonous plant problems. Thus, the cost for lost calves and lambs would be: $35,630,400 + $3,211,200 = $38,841,600.
In summary, if one accepted the above estimates of poi- sonous plant losses to the livestock industry in the 17 West- ern States, the loss would be:
Cattle-1% of total adult animals $55.7 million
Cattle-l% of calf crop $35.6 million
Sheep--3.5% of total adult animals $12.5 million
Sheep-l% of lamb crop $ 3.2 million
TOTAL $107.0 million1
Because of the diversity of local situations and the com- plete lack of data, no attempt will be made to estimate the losses caused by less than optimum use of the range resources, reduced weaning weights, and the extra cost of management caused by poisonous plants. However, these losses are real and should be included if data were available.
It is hoped that those of you who do not agree with the above estimate will do some serious thinking and research to make better estimates of these losses.
Poisonous plants pose another serious problem to the rancher that was eluded to above. A rancher operating in an area where poisonous plant problems exist may have learned to manage his animals and his rangelands so as to keep death and other losses caused by these plants at man- ageable levels. Ranchers in this situation have to continually face another problem. At any time, a given set of circum-
1 Increases in the supply of beef and lamb of this magnitude would have a negative impact on prices, depending on the elasticity of demand. As more refined estimates of poisonous plant losses are made. this should be given consideration.
stances can cause them losses of such magnitude that they are ruined financially. For example, a windstorm blows pine trees down and his cattle consume enough needles to cause an abortion rate of SO-90% in his cow herd. A sheep rancher grazes his sheep on a range infested with halogeton. He knows how to manage his herds in such a way that he gets by with low losses for several years. Then a combination of factors occurs at one time and he loses most of his herd in one day.
Dr. Lynn James, Director, Poisonous Plant Research Laboratory, Logan, Utah, provided the following examples of severe losses. All of the following losses were reported from halogeton. Sheep herds grazing on the Utah-Nevada line had losses of 450 head, 600 head, and 800 head. Some
1,200 head of sheep were killed in Antelope Valley, Utah. A Nevada sheep rancher lost 1,300 sheep at one time. Another rancher lost over 2,000 head of sheep over a period of a couple of years.
Locoweed is another poisonous plant that can cause losses severe enough to put a rancher out of business. In 1972 it was estimated that 20% of all cattle in Southwestern New Mexico were poisoned on locoweed. They were not all killed but their productivity was reduced. In 1958 over6,OOO sheep were killed on locoweed in the Uintah Basin of Eastern Utah. In 1964 one rancher lost $125,000; another rancher lost $55,000; and a third rancher lost $65,000 worth of sheep to locoweed poisoning. Abortion rates of up to 100% in cattle and over 70% in sheep were reported from locoweed poisoning.
Tall larkspur (DeZphinium barbeyi Huth) poisoning caused the death of 103 mature cattle (valued at $25,750) on one Forest Service allotment in Utah in 1958. The average annual loss was 36 head of cattle (valued at $9,000) over a 15-year period (Nielsen and Cronin 1977).
There is not an abundance of material in the literature concerning the economics of poison plant loss prevention. Nielsen and Cronin (1977) reported on the economics of tall larkspur control. Control of tall larkspur plants with chemical herbicides resulted in a 90+ % decrease in cattle death losses. The return on ranchers and Forest Service investment in this project ranged from 65% to 74% This is an excellent return, much better than most investment opportunities in the ranching business. There was enough tall larkspur on this allotment to cause an average annual loss of 36 adult cattle. Yet this amount of larkspur could be economically controlled if only ten cows were saved each year for ten years. The point is that there are many situations where losses are not as heavy as in the above cited study but could still be controlled economically.
In summary, there appears to be a need and an oppor- tunity for more research on the economic importance of poisonous plants and on the economics of control projects that would reduce these losses. The net result would be a more efficient livestock industry and more food and fiber from our natural resources. It appears that many of these projects are environmentally safe and economically feasible.
Literature Cited
Durrell, L. W., and I. E. Newsom. 1939. Colorado’s poisonous and injur- ious plants. Colorado Agr. Exp. Sta. Bull. 455.
Gilkey, H. M. 1958. Livestock-poisoning weeds of Oregon. Oregon Agr. Exp. Sta. Bull. 564.
Goodsell, W. D., and M. Belfield. 1973. Costs and returns, migratory-sheep ranches, Utah, Nevada, 1972. U.S. Dep. Agr., Agr. Res. Serv. 523.
Gray, J. R. 1970. Production practices, costs and returns in the central mountains of New Mexico. New Mexico Agr. Exp. Sta. Res. Rep. 166.
Gray, J. R., and C. B. Baker. 1953. Cattle ranching in the northern Great Plains. Montana Agr. Exp. Sta. Circ. 204.
James, L. F. 1978. Livestock poisoning by plants. U.S. Dep. Agr., Agr. Res. Serv. Unpublished mimeo.
Kearl, W. G. 1967. Comparative livestock systems and technologies for Wyoming northern plains cattle ranching. Special report to the W-79 Technical Committee.
Keeler, R. F., K. R. Van Kampen, and L. F. James (editors). 1978. The Effect of Poisonous Plants on Livestock. Academic Press, New York.
Myles, G. A. 1962. Cost and returns on livestock ranches in north-western Nevada in 1961. Nevada Agr. Exp. Sta. Mimeo Circ. 166.
Nielsen, D. B., and E. H. Cronin. 1977. Economics of tall larkspur con- trol. J. Range Manage. 30:434-438.
Norris, J. J., and K. A. Valentine. 1957. Principal livestock-poisoning plants of New Mexico ranges. New Mexico Agr. Extension Serv. Circ. 274.
Roberts, N. K., and C. K. Gee. 1963. Cattle ranches using public ranges year-long. Utah Agr. Exp. Sta. Bull. 440.
Sperry, 0. E., J. W. Dollahite, G. 0. Hoffman, and B. J. Camp.(Undated).
Texas plants poisonous to livestock. Texas Agr. Extension Serv., Texas Agr. Exp. Sta. Bull. 1028.
Stoddart, L. A., and A. D. Smith. 1943. Range Management. McGraw- Hill Book Co., Inc., New York and London. 430 p.
U.S. Department of Agriculture. 1973. Cattle raising in the United States. U.S. Dep. Agr., Econ. Res. Serv., Agr. Econ. Rep. 235.
U.S. Department of Agriculture. 1975-76. Meat animals, production, dis- position, income. Crop Reporting Board, Statistical Reporting Service.
U.S. Department of Agriculture. 1977. Beef cattle numbers, January I, 1977. Crop Reporting Board, Statistical Reporting Service.
U.S. Department of Agriculture. 1977. Breeding sheep, I year and older. January I, 1977. Crop Reporting Board, Statistical Reporting Service.
The Ecological Niches of Poisonous Plants
in Range Communities
E. H. CRONIN, PHIL OGDEN, JAMES A. YOUNG, AND WILLIAM LAYCOCK
Highlight: So many diverse plant species are poisonous to domestic livestock that it seems highly improbable that a universal competitive advantage is common to all poisonous species. Plant poisons may have originated in mutations that were of no direct adaptive value. Once these secondary chemical products became established in the physiologic systems of plants, their interactions with insects and rodents that consumed the seeds, seedlings, or herbage of the plant may have given them adaptive value. Evolution of resistance to poisons of plants by the herbivores probably has led to the proliferation of species in genera that contain many poisonous species such as Astragalus. The study of such relationships should be given priority in range ecology to achieve effective management of the range resource. Some poi- sonous plant species have also evolved allelopathic defense mechanisms that enhance competitive advantages. The coevolu- tion of poisonous plants, large herbivores, and rumen microfloras offers intriguing possibilities for study that may answer questions basic to the future success of range management.
Because of the tremendous economic losses that the livestock industry incurs from plant-induced poisoning, it becomes imperative that range managers study the ecology of poisonous plants in rangeland communities. This topic is difficult to cover adequately in a short presentation because of the number of poisonous plants and the variations among them. We have relatively few precise data or facts, which are the finished products of research. Instead we must deal with
Authors are plant physiologist. U.S. Department of Agriculture. Science and Education Administration, Federal Research, Poisonous Plant Research Laboratory, Logan. Utah 84321: Dep. of Range Management, University of Arizona, Tucson 84719; professor of range management, School of Renewable Resources, University of Arizona, Tucson 84719; range scientist, U.S. Dep. Agr., SEA-FR, Renewable Resource Center, University of Nevada, Reno 89512; and range scientist, U.S. Dep. Agr., SEA-FR. Crops Research Laboratory. Fort Collins. Colo. 80521.
Manuscript received March 30. 1978.
328
ideas, which are the essential raw materials of research. We will discuss the diverse habitats occupied by poisonous species and speculate on the role of secondary chemical compounds in the ecology of plants.
Spectrum of Poisonous Plants
Many authors(e.g., Sampson and Malmsten 1935; Muen- scher 1958) have attempted to formulate a definition of a poisonous plant. The following elements are basic to all definitions: (a) a poisonous plant contains some specific substance, which (b) when consumed by herbivores under specific circumstances, (c) causes injury to susceptible animals. Purely mechanically injurious plants are usually exempted from such definitions because they lack the specific injurious chemical agent.
The “specific circumstances” under which the poisonous plants must be consumed pertains to the modifying role of variable environmental conditions in determining what plants are poisonous. For example, a plant may provide excellent forage while it is rapidly growing, but become toxic after a frost.
Species, breeds, and individual grazing animals differ in susceptibility to plant poisoning. Losses of sheep from halogeton (Halogeton glomeratus) are sometimes very great, but cattle often use halogeton-infested ranges without apparent ill effects even though cattle can be poisoned from consuming halogeton. Conversely, larkspur (Delphinium spp.) is not considered to be a major threat to sheep, but it is a major threat to cattle (Stoddart et al. 1975). The physical condition of the consuming animal also influences its sus-
Gray, J. R. 1970. Production practices, costs and returns in the central mountains of New Mexico. New Mexico Agr. Exp. Sta. Res. Rep. 166.
Gray, J. R., and C. B. Baker. 1953. Cattle ranching in the northern Great Plains. Montana Agr. Exp. Sta. Circ. 204.
James, L. F. 1978. Livestock poisoning by plants. U.S. Dep. Agr., Agr. Res. Serv. Unpublished mimeo.
Kearl, W. G. 1967. Comparative livestock systems and technologies for Wyoming northern plains cattle ranching. Special report to the W-79 Technical Committee.
Keeler, R. F., K. R. Van Kampen, and L. F. James (editors). 1978. The Effect of Poisonous Plants on Livestock. Academic Press, New York. Myles, G. A. 1962. Cost and returns on livestock ranches in north-western
Nevada in 1961. Nevada Agr. Exp. Sta. Mimeo Circ. 166.
Nielsen, D. B., and E. H. Cronin. 1977. Economics of tall larkspur con- trol. J. Range Manage. 30:434-438.
Norris, J. J., and K. A. Valentine. 1957. Principal livestock-poisoning plants of New Mexico ranges. New Mexico Agr. Extension Serv. Circ. 274.
Roberts, N. K., and C. K. Gee. 1963. Cattle ranches using public ranges year-long. Utah Agr. Exp. Sta. Bull. 440.
Sperry, 0. E., J. W. Dollahite, G. 0. Hoffman, and B. J. Camp.(Undated).
Texas plants poisonous to livestock. Texas Agr. Extension Serv., Texas Agr. Exp. Sta. Bull. 1028.
Stoddart, L. A., and A. D. Smith. 1943. Range Management. McGraw- Hill Book Co., Inc., New York and London. 430 p.
U.S. Department of Agriculture. 1973. Cattle raising in the United States. U.S. Dep. Agr., Econ. Res. Serv., Agr. Econ. Rep. 235.
U.S. Department of Agriculture. 197576. Meat animals, production, dis- position, income. Crop Reporting Board, Statistical Reporting Service.
U.S. Department of Agriculture. 1977. Beef cattle numbers, January I, 1977. Crop Reporting Board, Statistical Reporting Service.
U.S. Department of Agriculture. 1977. Breeding sheep. I year and older. January I, 1977. Crop Reporting Board, Statistical Reporting Service.
The Ecological Niches of Poisonous Plants
in Range Communities
E. H. CRONIN, PHIL OGDEN, JAMES A. YOUNG, AND WILLIAM LAYCOCK
Highlight: So many diverse plant species are poisonous to domestic livestock that it seems highly improbable that a universal competitive advantage is common to all poisonous species. Plant poisons may have originated in mutations that were of no direct adaptive value. Once these secondary chemical products became established in the physiologic systems of plants, their interactions with insects and rodents that consumed the seeds, seedlings, or herbage of the plant may have given them adaptive value. Evolution of resistance to poisons of plants by the herbivores probably has led to the proliferation of species in genera that contain many poisonous species such as Astragalus. The study of such relationships should be given priority in range ecology to achieve effective management of the range resource. Some poi- sonous plant species have also evolved allelopathic defense mechanisms that enhance competitive advantages. The coevolu- tion of poisonous plants, large herbivores, and rumen microfloras offers intriguing possibilities for study that may answer questions basic to the future success of range management.
Because of the tremendous economic losses that the livestock industry incurs from plant-induced poisoning, it becomes imperative that range managers study the ecology of poisonous plants in rangeland communities. This topic is difficult to cover adequately in a short presentation because of the number of poisonous plants and the variations among them. We have relatively few precise data or facts, which are the finished products of research. Instead we must deal with
Authors are plant physiologist, U.S. Department of Agriculture, Science and Education Administration, Federal Research. Poisonous Plant Research Laboratory. Logan. Utah 84321; Dep. of Range Management. University of Arizona, Tucson 84719; professor of range management. School of Renewable Resources, University of Arkona. Tucson 84719: range scientist. U.S. Dep. Agr., SEA-FR, Renewable Resource Center, University of Nevada, Reno 895 12; and range scientist. U.S. Dep. Agr.. SEA-FR, Crops Research Laboratory, Fort Collins. Cola. 80521.
Manuscript received March 30. 1978. 328
ideas, which are the essential raw materials of research. We will discuss the diverse habitats occupied by poisonous species and speculate on the role of secondary chemical compounds in the ecology of plants.
Spectrum of Poisonous Plants
Many authors (e.g., Sampson and Malmsten 1935; Muen- scher 1958) have attempted to formulate a definition of a poisonous plant. The following elements are basic to all definitions: (a) a poisonous plant contains some specific substance, which (b) when consumed by herbivores under specific circumstances, (c) causes injury to susceptible animals. Purely mechanically injurious plants are usually exempted from such definitions because they lack the specific injurious chemical agent.
The “specific circumstances” under which the poisonous plants must be consumed pertains to the modifying role of variable environmental conditions in determining what plants are poisonous. For example, a plant may provide excellent forage while it is rapidly growing, but become toxic after a frost.
Species, breeds, and individual grazing animals differ in susceptibility to plant poisoning. Losses of sheep from halogeton (Halogeton glomeratus) are sometimes very great, but cattle often use halogeton-infested ranges without apparent ill effects even though cattle can be poisoned from consuming halogeton. Conversely, larkspur (Delphinium spp.) is not considered to be a major threat to sheep, but it is a major threat to cattle (Stoddart et al. 1975). The physical condition of the consuming animal also influences its sus-
ceptibility. Animals may be susceptible to plant poisons that produce birth defects (teratogenic effects) only at specific stage of pregnancy (Binns et al. 1965). Animals may be sus- ceptible to a plant poison only if they are preconditioned by a specific diet. For example, for sheep to be photosensitized by eating horsebrush (Tetradymia spp.) they must have pre- viously eaten black sagebrush (Artemisia nova) (Johnson 1978). In general, animals in poor condition are less suscept- ible to nitrate poisoning than animals in good condition.
The type of injury produced by the plant poison may be highly variable. We usually associate poisonous plants with acute symptoms and death losses, but chronic ill health or pregnancy-associated symptoms also may be a result of plant poisoning.
In ruminants, which are our major consumers on range- lands, rumen microflora break down high-cellulose-content forage and synthesize nutrients. Some types of plant poisons, such as essential oils, may indirectly harm rumi- nants by interfering with the function of the rumen micro- flora (Nagy et al. 1964). The rumen microflora of sheep can break down oxalates if they have foraged on plants con- taining small quantities of oxalates for a short period (2-3 days). Then they can ingest as much as 75% more halogeton without ill effects (James and Cronin 1974).
Community Ecology of Poisonous Plants
The great variation in poisonous plants find them repre- sented in a host of plant communities. In the traditional view of the ecology of poisonous plants, the occurrence of poisonous plants is equated with poor range condition. For example, Heady (1975) states in a recently published textbook on range management, “If rangeland is properly maintained in excellent condition, losses from poisonous plants can be effectively reduced.” This assumption is probably influenced by three factors: (a) some poisonous plants are colonizers that increase in density with over- grazing; (b) many plant poisons are dosage dependent and as desirable forage plants decrease in density with declin- ing range condition, grazing animals consume proportion- ately larger amounts of the toxic plants; and (c) animals in poor physical condition from grazing degraded rangelands are often most susceptible to plant poisoning (except for nitrate poisoning as previously noted).
Pristine Communities
The water hemlocks (Cicuta spp.) are among the acutely poisonous species that occupied specific habitats under pristine conditions and that still are restricted to specific habitats (Brewer et al. 1876; Greene 1889; March et al. 19 14; Fleming et al. 1920). Water hemlock may be the source of reports by early trappers and explorers of poison waters in the West (Vestal 1974). The plant is certainly responsible for the naming of two streams-the “Malad River” (from the French word “maladie” meaning sickness); one is on the Utah-Idaho border and empties into the Great Salt Lake, and the other is a tributary of the Snake River near Twin Falls, Idaho (Howell 1960). The specific marshy habitats favored by water hemlock probably have decreased with the drainage of wet meadows to favor forage production.
Although they are adapted to more variable and contrast- ing environments than water hemlock, the larkspurs (Del-
phinium spp.) also occur in pristine plant communities in equilibrium with their environments (Sampson 1944; Flem- ing et al. 1933; Kurameto and Bliss 1970; Franklin and Dyrness 1973; Sawyer and Thornburgh 1977). The lark- spurs have persisted after the introduction of domestic livestock to western rangelands. The tall larkspurs grow in the mountains on sites with deep soils. They tend to remain green throughout the growing season. Tall larkspurs require an almost continuous supply of soil moisture throughout the growing season. They are climax species as an under- story under trees or in open meadows. They can be the most tenacious species of the pristine vegetation, persisting after the associated species are destroyed. But some species of tall larkspur, such as D. barbeyi, may also increase in density when grazing pressure is reduced (Ellison 1954).
Seral Dominants
Horsebrush (Tetradymia canescens) may occur as an occasional species in Artemisia communities in high con- dition, but often forms pure stands after disturbance of a community (Young and Evans 1974). A prolific root sprouter, horsebrush assumes dominance when the non- sprouting big sagebrush (Artemisia tridentata) is burned in wildfires. After a period of 15 to 20 years without repeated burning, big sagebrush reestablishes and horsebrush de- creases in dominance.
If the frequency of disturbance is increased as with promiscuous burning, the tenure of horsebrush dominance is prolonged. As a component of the natural environment, the poisonous horsebrush will not completely disappear with succession, which we usually equate with higher range condition, but it will greatly decrease in abundance.
Decreaser With Burning
Burroweed (Haplopappus tenuisectus) is a half-shrub that commonly grows on sandy loam and sandy clay-loam soils of the upper southern desert-shrub and lower desert- grassland regions of southwestern Texas, New Mexico, Arizona, and Mexico. Schmutz et al. (1968) listed burro- weed as one of the 30 major livestock poisoning plants of Arizona. The poisonous principle is a higher alcohol, tremetol, which also is the poisonous principle of white snakeroot (Eupatorium rugosum), a species that grows in most habitats of the eastern U. S. and Canada (Kingsbury 1964). Both of these species are members of the Compositae plant family, and livestock poisoning from their consump- tion is most prevalent when more palatable forage is in short supply (Kingsbury 1964; Schmutz et al. 1968). Jimmyweed
(Haplopappus heterophyllus) is closely related to burro- weed and also contains tremetol. Livestock raised in a burroweed area seem to develop an aversion to it, so locally raised livestock are seldom poisoned by burroweed inges- tion unless they are stressed for forage. Severe losses have occurred when livestock not raised in burroweed areas were grazed on ranges with abundant burroweed, especially when grass forage was dry, in May or June.
Burroweed germinates and grows in the cool winter and spring months, and mortality is high during dry winters. The density of burroweed stands, therefore, fluctuates greatly from year to year (Martin 1966). Mortality of burroweed is
also high in spring or summer fires (Martin 1966). Burro- weed seeds germinate immediately following maturity (Mey- er 1973) when cool-season moisture is available and, therefore, may become established on sites where warm- season grasses have difficulty in becoming established. Burroweed plants may, therefore, provide protection and favorable microsites for the establishment of desert-grass- land grass species on harsh desert shrub sites in favorable years. Cable (1969) and W. Meyer, (unpublished data Univ. of Arizona Student Report) showed that the competition for moisture between warm-season perennial grasses and bur- roweed is not great.
The ecological niche that burroweed occupies seems to be mostly related to its adaptations for germination and growth in the cool season. The low palatability of burro- weed, whether this is a result of the poisonous principle or some other factor, and the reduced frequency of fire on desert grasslands have increased the abundance of this species in the past century. Note that horsebrush increases through rootsprouting after promiscuous burning, whereas burroweed, which depends on seedlings for establishments, increases when fires are suppressed.
Grazing Increaser
Orange sneezeweed (Helenium hoopesii) is another ex- ample of poisonous species that occurs naturally and is of minor importance in pristine plant communities, but in- creases with misuse or overgrazing. Orange sneezeweed occurs in a variety of vegetation types on high elevation rangelands in most of the western states. Where the range is properly managed, orange sneezeweed is not a common species (Ellison 1954) and often occurs in small, scattered patches. On overgrazed ranges, sneezeweed may be locally abundant or may even be the dominant species on extensive areas (U.S. Forest Service 1937).
Alien Species
Considering the host of alien annuals that have been introduced to western rangelands (Young et al. 1972), it is remarkable that so few species are poisonous. One major exception is halogeton. This annual weed was introduced to the intermountain area of western North America sometime before 1934. Cronin (1965) described halogeton as “a pioneer invader on disturbed sites in the salt-desert shrub vegetation.” Abundant halogeton usually indicates de- graded rangelands, but even if such communities could be restored to their pristine balance, limited amounts of halogeton could probably persist on unused environmental potential.
The Ecological Advantage of Being Poisonous
Poisonous plants obviously have a variety of successional roles and ecological advantages that defy generalization. The basic question considered in this review is, “Pees being poisonous always convey any competitive advantage in this wide spectrum of habitats?”
No matter how carefully we state our purpose, we must approach the question obliquely because, in the words of Stebbins (1974): “The evolutionist cannot trace directly the alterations of genes and gene-controlled processes that have
been responsible for evolutionary trends. He can observe only the outcome of these changes in terms of alterations in the morphology and the reactions of the adult organisms.” In this case the alterations are in the physiologic systems of plants, making them poisonous.
Recently, our understanding of the evolutionary signifi- cance of secondary chemical substances in plants has been advanced through research stimulated by the realization that these substances can serve as markers for taxonomic relationships. The use of chromatographic characteristics to establish phylogenetic relationships for the Tridentatae section of Artemisia (Hanks et al. 1973) is an excellent example of such research. This heightened interest has also been expressed in speculation about their value to the plant. Why should most conifers of the temperate zone be richly endowed with terpenes (Levin 1971), and why should alkaloids be widespread in Ranunculaceae (McKay 1974), tannins in Fagaceae (Feeny 1970), essential oils in Labiatae (Stebbins 1974)?
One answer may be that these substances are inevitable products of cellular metabolism which are difficult to excrete and must, therefore, somehow be disposed of by some other route. Stebbins (1974) finds this explanation unsatisfactory for two reasons. First, it does not explain why certain groups of plants have evolved conducting systems, whose elaborate development must be controlled by a large number of genes (Mahlberg 1959), for secondary chemical substances. For example, the families Euphorbiaceae, Apo- cynaceae, and Asclepiadaceae have evolved not only latex substances, but the specialized and intricate conducting system that ensures the presence of abundant latex in all parts of the plant. Second, secondary chemicals in plants are probably not inevitable products of cellular metabolism. The process of artificial selection used in developing many cultivated derivatives of wild species, such as garden lettuce
(Lactuca sativa), has greatly reduced their content of these secondary substances (e.g., those producing bitter taste), but has not reduced the metabolic efficiency or vigor of cultivated lettuce. If the secondary chemicals were necessary byproducts of essential metabolic processes, their removal would surely have disrupted the growth and reproduction of the selections.
In a recent review of chemical interactions between species, Whittaker and Feeny (197 1) popularized an explan- ation of the function of secondary chemicals that was first proposed by Stahl (1888). Stahl suspected that the ability to form secondary substances that are bitter, toxic, or both, (phenylpropanes, acetogenins including flavones and antho- cyanidins, terpenoids, and alkaloids) evolved in plants in response to natural selection for the capacity for defense against herbivores. This conclusion is now supported by a large body of evidence (Whittaker and Feeny 1971).
Assuming that the production of a toxic substance by plants is always a defense against grazing predation may be erroneous. For example, the production of oxalates by halogeton may be part of a mechanism to reduce moisture stress. The succulence of halogeton is proportional to the concentration of sodium or potassium chloride in the growth medium, but the major role in promoting succulence is attributed to the chloride ions. The chloride ion was thought to be the element responsible for increased succu- lence (Williams 1960). If so, then the absorption of potas- sium or sodium chloride must result in an excess of potassium or sodium cations. Production of oxalic acid
provides a means of tieing up the excess cations to produce an acceptable sodium balance in halogeton (Waisel 1972). This explanation appears logical in view of the increasing concentration of oxalates in halogeton (Williams 1960) as the soil moisture is depleted over the growing season (Cronin 1965).
The evolution of secondary substances as defense mechan- isms appears to be an excellent example of the interaction of natural selection, chance, and adaptive modification along the lines of least resistance. The particular pathway taken by an evolutionary line depends to a certain extent upon chance combinations of genes that exist in the initial population of the line as a result of mutation. For poisonous plants the chance mutation controls the production of a toxic chemi- cal or the loss of the enzyme required to metabolize the toxic chemical. Stebbins (1950) argued that an equally or perhaps more important factor in determining the pathway of adaptation is the innate, genetically controlled pattern of development that exists in a population at any stage in its evolution. This pattern is important because the direction of adaptation will often be determined according to the principle of adaptive modification along the lines of least resistance, i.e., modification involving the least possible alteration of existing biosynthetic pathways.
We can illustrate this principle with the genus Astragalus. Barneby (1964) listed 368 species of North American Astragalus. Williams and Barneby (1977) determined that 204 of these species contain nitrocompounds that are poten- tially poisonous to grazing animals. Some ancestral species of Astragalus may have evolved by chance mutation the initial nitrocompound. As the genus has proliferated in a variety of habitats the process of speciation has probably been accompanied by modification of the original nitrocom- pound. Green and Palmblad (1975) found that chemicals in the seed pods were partly responsible for differential preda- tion on seeds of A. cibarius and A. utahensis. They concluded that selection pressure by seedeating insects may account for much of the chemical diversity exhibited by the genus Astragalus. These nitrocompounds may provide an adaptive advantage in protecting Astragalus plants from attacks by insects, rodents, or both during some portion of the plants’ life cycle. Diversification may well have been largely a matter of evolving new substances for defense against predators that had evolved immunity to the preexist- ing ones.
Thus, the operation of adaptive modification along the lines of least resistance during the speciation of Astragalus is demonstrated by the spread of nitrocompounds or their antilogies. It does not result in the evolution of new alkaloids or other families of toxic compounds; i.e., pre- existing bisynthetic pathways were altered as little as possible. Small changes lessen the chance of disrupting the physiology of the plant.
Allelopathic Plants
In addition to compounds that act as direct defenses against destructive herbivores, many adult plants produce allelopathic substances that prevent the growth of com- peting species in the immediate vicinity (Rice 1974). Allelo- pathic plants are of interest in this discussion because of the evolutionary parallel in selection for allelopathic and poi- sonous substances in plants. In many areas of former salt desert vegetation in eastern Nevada and western Utah, the
plant cover consists almost entirely of the alien species halo- geton. Small quantities of halogeton are utilized by livestock and wild herbivores throughout the growing season, regard- less of the amount of available forage, but lethal levels are consumed only when other feed is limited or absent. The poisonous principle in halogeton is soluble oxalates (James 1972). Most losses occur when hungry animals are intro- duced to vegetation with abundant halogeton and after they are watered following a prolonged period of acute thirst (James and Cronin. 1974).
Eckert and Kinsinger (1960) determined that the leachate from halogeton increased the soil pH, electrical conduc- tivity, exchangeable sodium and potassium, soluble sodium, potassium, calcium, and magnesium, modulus of rupture; and decreased percolation and capillary rise of water. The density of halogeton increases as the salinity of the soil increases (Williams 1960). Once halogeton is established and has modified the soil, the site is closed to the estab- lishment of other plant species, affording a tremendous advantage to halogeton (Kinsinger and Eckert 1961).
A similar type of allelopathic effect through the concen- tration of soluble salts occurs with the poisonous native shrub greasewood (Fireman and Hayward 1952). However, the widely spaced greasewood plants are less effective than dense herbaceous stands of halogeton in eliminating compe- tition from other species. The widely distributed bracken fern (Pteridium aquilinum) contains a cyanogenic glycoside (Cooper-Driver and Swain 1976) that is poisonous to livestock (Muenscher 1958). Water extracts from bracken fern significantly reduced growth, germination, or both in competing herbaceous vegetation (Gliessman and Muller
1972). The phytotoxic principle was suspected to be a phenolic compound.
Natural Selection and Poisonous Plants
If we assume that being poisonous reduces the amount of photosynthetic tissue removed by herbivores and, therefore, improves the survival characteristics of the plant, then the selective advantage is obvious. However, with large herbi- vores such as cattle, horses, and sheep on western range- lands of the United States this assumption is probably false for two reasons. First, being poisonous cannot reduce consumption of a plant’s photosynthetic tissue by livestock unless the plant is so acutely toxic that the animals drop dead at the first taste or unless the animals sense the toxicity of the plant and do not graze the herbage. We know from the tremendous economic losses from plant poisoning that both situations are not always met. An example of an effect opposite to that in our assumption is found in the Astragalus species that cause locoism: once animals graze on locoweeds they tend to seek and graze them to the exclusion of other forage. Second, plants in the United States west of the Rocky Mountains have been grazed by large herbivores for so short a time that grazing pressure may not have contributed to selection of plants in this region. Most of the large herbivores native to North America were lost by Pleistocene extinction (Martin 1967); west of the Rocky Mountains the depletion of large herbivores was especially significant. The major native plant species that cause poisoning of domestic livestock on far western rangelands probably evolved for a considerable period of time relatively free of grazing by concentrations of large herbivores.