Copyright 0 1996 by the Genetics Society of America
Low Frequency of Mouse t Haplotypes in
Wild Populations Is Not
Explained
by
Modifiers of Meiotic Drive
Kristin
G .
Ardlie andLee
M.
Silver
Department of Ecology and Evolutionaly Biology and Department of Molecular Biology, Princeton Univasity, Princeton, New Jersey 08544
Manuscript received March 1, 1996
Accepted for publication September 13, 1996
ABSTRACT
t haplotypes are naturally occurring forms of mouse chromosome 17 that show non-Mendelian trans- mission from heterozygous + / t males. In laboratory studies, transmission ratios of 20.90 or higher are typically observed. With transmission ratios of this level, theoretical analyses predict high frequencies of
t haplotypes (- 75%) in wild populations. In contrast, empirical frequencies of only 15-25% are typically found. This has led to the suggestion that modifiers of drive may play a role in reducing t frequencies. We have measured transmission ratio distortion (TRD) levels in wild +/tmice to examine this hypothesis. TRD was very high in both litters collected from wild-caught pregnant females, and in wild litters bred in the laboratory (mean = 0.9). Contrary to the results of other studies, we found no difference in TRD levels between semilethal and lethal t haplotypes nor between litters conceived from cycling or postpar- tum estrus. We found three litters with aberrantly low TRDs that were all multiply sired, although the role this might play in natural populations is unknown. These findings show a general absence of modifiers of drive in natural populations and suggest that other factors are responsible for the low observed frequencies of wild t haplotypes.
T
HE t haplotypes of the mouse are the best known example of meiotic drive in mammals. They are a variant of the proximal third of chromosome1 7
that exist in natural populations of all subspecies of the Musmusculus species complex (DELARBRE et al. 1988; HAM- MER and SILVER 1993). All t haplotypes are character- ized by a series of four nonoverlapping inversions that suppress recombination between t and wild-type DNA
and result in the inheritance of t haplotypes as a unified genetic entity (ARTZT et al. 1982b; HERRMANN et al. 1986;
SARVETNICK et al. 1986b; HAMMER et al. 1989). The most important property of t haplotypes is their capacity to distort Mendelian segregation in their favor, through a male-specific phenotype known as Transmission Ratio Distortion (TRD). Although segregation in females is normal, >95% of the offspring of heterozygous + / t males can inherit the t-bearing chromosome (DUNN
and SUCKLING 1956; DUNN 1957a; SILVER 1985). This powerful selective advantage is counterbalanced by both the complete sterility of t/t homozygous males
(LYON 1986) and by the recessive lethal mutations car- ried by most t haplotypes (BENNETT 1975; ARTZT et al. 1982a; ARTZT 1984). Thus t haplotypes have not become fixed in mouse populations. These counterbalancing forces alone, however, do not adequately explain the low frequencies of t haplotypes observed in most popu- lations. For a transmission distortion of 95%, balanced
Corresponding author: Kristin Ardlie, Museum of Comparative Zool- ogy, Harvard University, 26 Oxford St., Cambridge, MA 02138. E-mail: [email protected]
Genetics 144: 1787-1797 (December, 1996)
by homozygous lethality, deterministic models predict an equilibrium frequency of
77%
( + / t ) heterozygotes(BRUCK 1957). Typically only 15-25% ofwild mice actu- ally carry t haplotypes as + / t heterozygotes (KLEIN et al. 1984; LENINGTON et al. 1988).
A number of models have addressed this discrepancy between predicted and observed frequencies o f t haplo- types. These have demonstrated that factors such as systematic inbreeding, interdemic selection, and re- duced fitness of + / t heterozygotes will result in lowered frequencies o f t haplotypes (LEWONTIN and DUNN 1960;
PETRAS 1967; YOUNG 1967; LEVIN et al. 1969; NUNNEY
and BAKER 1993). However, empirical data in support of these are largely lacking. All models illustrate the necessity of high levels of TRD in maintaining the poly- morphism for t haplotypes. Even small reductions in transmission ratio can result in reduced equilibrium frequencies and t haplotypes will be rapidly lost from populations if TRD is not high.
In the few studies of TRD performed on wild + / t males brought into the laboratory for breeding, trans- mission ratios in excess of 90% were typically observed
(DUNN 1957b; LEWONTIN and DUNN 1960; SILVER 1985;
1788 K. G. Ardlie and L. M. Silver
from the homologous chromosome (BENNETT et al.
1983; GUMMERE et al. 1986). A delay in the timing of mating can also reduce the TRD of some t haplotypes
(BRADEN 1958; BRADEN 1972).
Although a reduction in TRD, through either behav- ioral means or genetic modifiers of drive, could account for the low observed frequencies of t haplotypes (LEVIN
et al. 1969), no currently available empirical data ad- dress these possible mechanisms in natural populations. In this study, we report the first estimates of transmis- sion ratio distortion for mice in the wild and provide evidence that genetic modifiers of drive are not wide- spread, and are thus unlikely to account for the lower than predicted frequencies of t haplotypes. Using our data on the transmission ratios of males, we further test whether TRD levels differ between lethal and semilethal
t haplotypes and whether the type of estrus cycle during conception can influence transmission ratios, both of which have been reported in previous studies.
MATERIALS AND METHODS
The wild mice used in this study were trapped as part of a larger survey of the frequency and distribution of t haplotypes in natural populations (IS. &LIE and L. SILVER, unpublished data). Measures of transmission ratio distortion in wild mice were obtained in two ways: by opportunistically collecting DNA from pregnant females and their embryos at the site of capture in the field and by retaining live wild mice, from a subset of the trapping sites, for further breeding studies in the laboratory. All mice were sampled blind with respect to
t haplotypes, as the presence of t haplotypes could not be determined until DNA analyses were performed in the labora- tory.
TRD measured in litters collected from wild-caught preg-
nant females: All visibly pregnant female mice were trapped and killed to obtain both maternal and fetal tissue for DNA analysis. Maternal tissue was collected as a tail biopsy and stored in 95% ethanol. Pregnant females were dissected in the field, at the site of capture, and all fetuses were removed. The total number in each litter was recorded, and small or resorbed fetuses were recorded, but not collected. Fetal tissue was stored in 95% ethanol.
A total of 179 mothers and their litters were assayed for t
haplotypes with DNA markers as described below. Only a sub- set of those litters, containing + / t pups from a wild-type
+/
+
mother, were informative in estimating TRD, however. As the t haplotypes in the litter must have been inherited from the father, the proportion of young in the litter that contained a t haplotype was thus a direct measure of the effective trans- mission distortion from the presumed + / t father.TRD estimates from laboratory breeding of wild-caught mice: Mice: Wild mice from 18 different populations were retained for laboratory breeding studies. Due to space limita- tions in the breeding facility, ongoing mark-release-recapture studies, and the refusal of some wild mice to breed, the popu- lations analyzed for TRD are only a subset of all 64 indepen- dent populations trapped (K. ARDLIE and L. SILVER, unpub- lished data). The mice represented in this study were from four separate populations trapped near Princeton, NJ, two trapped near Knoxville, TN, two trapped near Ithaca, N u , six trapped near Carbondale, southern IL, and four from Somerset County, MD. Mice were considered as members of the same population if they were trapped in close location on
the same farm. Independent populations represent distinct locations, typically different farms, that were always separated by >5 km.
Wild mice of both sexes (from 5 to 10 individuals per popu- lation, depending on population size) were retained ran- domly from each of the different trapping locations and were brought back to Princeton University where they were assayed for the presence of t haplotypes. t haplotypes were at low frequency in wild populations overall (K. ARDLIE and L. SII, WR, unpublished data), and mice carrying t haplotypes were present in five of the 18 populations. All mice carrying t haplo- types were retained for breeding, and the t haplotype chromo- some was uniquely labeled with reference to the site at which it was caught and by the ID number of the mouse that origi- nally carried the chromosome (for example, CF109 was from mouse number 109 caught in a chicken farm in Ithaca, NY, and so forth). Wild-type mice, predominantly from the popu- lations that did not contain t haplotypes, were also retained to provide wild females for breeding.
Complementation analysis: Mice carrying each wild-derived t
haplotype were bred, primarily infmse, to determine whether they carried a lethal allele. Mice found to be carrying lethal
t haplotypes were then bred to one another to determine whether they carried complementing or noncomplementing lethal alleles. Determining the identity of the complementa- tion groups of each lethal allele was not a goal of this study, however, mice carrying wild lethal alleles were also bred to the laboratory-maintained Y5 haplotype to check for comple- mentation with this lethal t haplotype which is common among North American wild mice (BENNETT 1978).
Transmission Ratio Distortion analysis: TRD was estimated for all t haplotypes by breeding wild-derived + / t males to wild- derived wild type (+/+) females. Males were mated sequen- tially to 1-5 females (mean = 2), although each male re- mained caged with the same female to produce four or more litters if possible. Females were allowed to raise their litters to weaning age (3-4 wk), pups were then sexed, and tail biopsies were taken to determine chromosome 1 7 genotypes using the DNA techniques described below. Wildderived
+/
t males were also bred to 129-SvJ inbred strain (+/+) females as a control for female genetic background.
In total, 12 original founder males and 47 first and second generation wild-bred males were examined for TRD. First- and second-generation males were offspring of founder males bred to wild
+/+
females derived predominantly from popu- lations different from that of the male. Variation in TRD among males and among different t haplotypes was examined by breeding all wild-derived + / t males to wild-derived+/+
females, both from populations where t haplotypes were pres- ent and absent, as well as to 129-SvJ females as a control. These varied breeding protocols were conducted to reveal possible effects of different wild genetic backgrounds and female genotype on TRD and/or the presence of modifiers of drive in some populations but not others. Differences in TRD among different chromosomes or males (e.g., lethal us. semilethal, PPE us. CE, etc.) were analyzed by one-way analysisof variance on arcsine transformed transmission ratios (ZAR 1996).
TRD of t Haplotypes in Wild Mice 1789
following the methods of LENINGTON and HEISLER (1991). Only litters born 5 3 0 days after the preceding litter were scored as being conceived during PPE. All other litters were scored as being CE-conceived. To compare the results ob- tained here with those of the earlier study, a female's first litter, which always results from cycling estrus, was excluded from the analysis (LENINGTON and HEISLER 1991). Because no significant difference in TRD was found between matings of wildderived males with either wildderived or inbred 129 females (see RESULTS), data from both of these types of mat- ings were pooled for use here.
DNA analysis: The presence of a t haplotype was deter- mined with markers that identify restriction fragment length polymorphisms (RFLPs) between t haplotypes and wild type forms of chromosome 17 (HERRMANN et al. 1986; SCHIMENTI
et al. 1987). High molecular weight DNA was prepared from tail clippings following standard protocols (HOGAN et al.
1994). DNA was isolated from field-collected fetuses either by cutting small pieces from large embryos, or by taking small embryos whole. Different embryos within each litter were treated separately from one another, and DNA was prepared in the same way as for the tail samples.
Approximately 10 pg of DNA was digested to completion with Tap1 restriction endonuclease (New England Biolabs). The digested DNA samples were electrophoresed, blotted, and bound to nylon membranes by UV crosslinking. The membranes were probed with labeled Bb-40 or Tu119 clones overnight at 65" according to the protocol of CHURCH and GILBERT (1984). Bb40 is a cosmid subclone which defines the locus D l 7Leh66band detects four TaqI restriction fragments in complete t haplotypes that are not found in wild-type chromo- some 17 DNA (SCHIMENTI et al. 1987). Tu119 is a genomic clone derived by microdissection (ROHME et al. 1984), which detects different alleles in t haplotype and wild-type DNA (HERRMANN et al. 1986). Probes were radiolabeled using ran- dom oligonucleotides on templates of denatured DNA (FEINBERG and VOGELSTEIN 1984). Individuals were scored as homozygous wild type (+/+) , heterozygous for a t haplotype ( + / t ) , or occasionally, homozygous t haplotype ( t / t ) .
PCR paterniq analysis: Among several litters with unusually low TRD levels, multiple paternity was unambiguously demon- strated in one by the presence of multiple Tu1 19 RFLP alleles among the offspring, indicating that at least three parents produced the litter. Thus the remaining litters with very low TRDs were also tested for evidence of multiple paternity. PCR amplification of microsatellite loci was used to detect the pres- ence of multiple sires. As paternity assignment was not a goal, only a few microsatellite loci were scored to enable detection of multiple paternity, Microsatellite loci used were D6Mitl2, D7Nds4, D9Mit4, DlONdsl, and D13Mit9 (Research Genetics).
PCR reactions were performed by radioactively labeling one of the primers and visualizing the products on sequencing gels. Genomic DNA from the mother and each offspring of a litter was amplified independently in a lG-pl PCR reaction using 0.25 units of Taqpolymerase. The reactions were ampli- fied for an initial cycle of 95" for 3 min, followed by 25 cycles at 95" for 1 min, 55" for 1 min, and 72" for 2 min.
RESULTS
Transmission ratio distortion in litters collected from the field A total of 179 litters were obtained from preg- nant females caught in the wild, and all fetuses and mothers were assayed individually for the presence of a t haplotype. The vast majority of litters were noninfor- mative. Because t haplotypes are rare in natural popula- tions of mice (K. ARDLIE and L. SILVER, unpublished
data), most litters had parents who were both wild type (+/+). Only 29 litters (16.2%) had t haplotypes segre- gating in them. These fell into five different classes (Table 1). In the seven litters of class 1, where the female was wild type ( + / + ) and the male carried a t haplotype, it was possible to estimate TRD directly. TRD was high in all of these litters, ranging from 0.8 to 1.0, with a mean TRD of 0.95.
In the two classes of litters where both parents carried t haplotypes (classes
2
and 4), complications from in- complete complementation make it difficult to derive an accurate estimate for TRD (BENNETT 1975). The litters produced from ( + / t ) females with wild-type males (class 3) showed near Mendelian inheritance, as expected, with a mean TRD of 0.45. A subset of litters derived from wild-type females had only a small number of offspring with a t haplotype that clearly must have been inherited from their father. TRD in these three litters was aberrantly low ranging from 0.1 to 0.2 with a mean of 0.17. By paternity analysis, all of these litters were shown to be sired by more than one male. These low TRD, double paternity litters define class 5 in Table 1.Complementation analysis Most complete t haplo- types from wild populations are associated with one or, less frequently, two recessive lethal mutations. To date, genetic studies have identified 16 lethal complementa- tion groups (KLEIN et al. 1984), each of which is likely to correspond to an independent locus within the inverted regions. All complete t haplotypes that do not carry a lethal mutation still cause embryonic lethality in a vari- able proportion of homozygous t / t embryos; these t haplotypes are referred to as semilethal t haplotypes.
Inter se breeding studies were performed on t haplo- types from different populations to distinguish among those that carried lethal mutations and those that were semilethal (Table
2).
Microsatellite analysis revealed that three populations (BM in NJ, MV in NJ, and PVin southern IL) each carried what appeared to be a single semilethal t haplotype, although the t haplotype in each population was distinguishable from the thaplo- types of other populations (ARDLIE and SILVER 1996). Complementation analysis demonstrated the presence of tW5 chromosomes in two other laboratory-analyzed
populations from Ithaca NY, and Tennessee. In addi- tion, the Tennessee population contained a second le- thal t haplotype of undetermined complementation group.
1790 K. G . Ardlie and L. M. Silver
TABLE 1
Estimates of TRD from litters obtained from wild-caught female mice
~ ~ ~~~~
Class Male genotype Female genotype No. of litters Mean litter size Mean TRD
~ ~~~~ ~~
1 + / t
+/+
2 + / t +/t
3
+/+
+/t4 +/t t/ t
5 + / t and unknown male
+/+
7 8.3 (5-10) 0.95
9 6.4 (5-9) NA
7 6.4 (4-9) 0.45
3 6.6 (5-9) NA
3 6.7 (5-10) 0.17
Range in parentheses. NA, not available.
1 ) . Eighty-one percent of all litters showed TRD levels between 0.8 and 1.0 (mean = 0.90). TRD levels were also calculated for each individual male with the vast majority ( ~ 9 0 % ) exhibiting values between 0.8 and 1.0 (mean male TRD = 0.902, and see Figure 2).
As
female genotype has previously been found to have an effect on the TRD of some t haplotypes (MCGRATH and HILLMAN 1980a), wild-derived+/
t males were also mated to 129-SvJ inbred females, for comparison, to determine if the transmission ratio in matings to wild female genotypes might be lower than in matings to an inbred laboratory female genotype (Figure 1 ) . Litters from wildderived and inbred females differed slightly in size, mean = 5.3 us. mean = 5.8,respectively (Mann-Whitney
U,100,2451
= 10,359, P = 0.025). However, there was no difference in the t trans- mission bias in these litters (Mann-Whitney UL100,245] = 11,606.5, P = 0.444). Thus, no evidence was found for a general effect of female genotype on TRD.Only 1.8% of all litters showed TRDs of 50.5, which was near that expected for a TRD of 0.9 (Figure 1 ) .
These were as likely to be derived from lethal or semile- thal t haplotypes and from postpartum estrus as cycling estrus (see below). No individual male produced a con- sistent string of low TRD litters and no t haplotype chromosome had a substantially lower TRD than all others (Table 2). Nor did any combination of wild
male/wild female pair produce litters with consistently reduced TRD. In general, although intramale heteroge- neity in TRD was observed, and a given male/female pair may produce a single litter with a low TRD ( 5 0 . 5 ) ,
in all instances both prior and subsequent litters pro- duced by the same pair were always high (0.9, data not shown). Additional litters sired by the same male with different females also always showed high levels of TRD.
We observed some intermale heterogeneity in TRD
levels. This was not related to the type of t haplotype or wild-type chromosome 17 the male carried, nor to the estrus cycle of the female, but was largely a function of the total number of offspring generated from each male (Figure 2). GUMMERE et al. (1986) observed similar levels of both intra- and intermale variability in TRD when they tested t haplotypes maintained on certain inbred strains in different heterogeneous genetic back- grounds and suggested that large numbers of offspring
( > l o 0 per male) need to be generated to accurately estimate TRD. Because of the difficulties of breeding wild mice of both sexes, we were unable to obtain this many offspring from our males, however, our data showed reduced variation in TRD levels above -50-60 offspring per male.
The expected binomial distribution of TRD levels was generated for the 345 litters for an assumed true TRD of
0.90. Although the two distributions were similar overall
TABLE 2
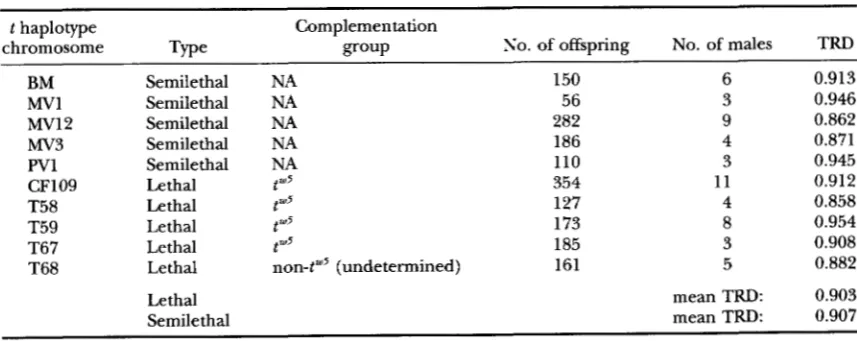
Transmission ratios of t haplotype chromosomes collected from wild populations
t haplotype Complementation
chromosome Type group No. of offspring No. of males TRD
BM Semilethal NA 150 6 0.913
Mvl Semilethal NA 56 3 0.946
Mv12 Semilethal NA 282 9 0.862
Mv3 Semilethal NA 186 4 0.871
PV1 Semilethal NA 110 3 0.945
CF109 Lethal t w5 354 11 0.912
T58 Lethal tu5 127 4 0.858
T59 Lethal r"5 173 8 0.954
T67 Lethal ,"5 185 3 0.908
T68 Lethal non-tw5 (undetermined) 161 5 0.882
Lethal mean TRD: 0.903
Semilethal mean TRD: 0.907
TRD of 1 Haplotypes in Wild Mice 1791
Matings to wild female mice Mean TRD = 0.90. n=245
Mean TRD = 0.91. n=100
Matings to 129-SvJ female mice
Expected distribution of all litters
for TRD = 0.9. n=345
0
Transmission Ratio Distortion
FIGURE 1.-Frequency distribution of TRD levels in litters sired by wild + / 1 males. Males were mated to both wild
+/
+
females and to 129-SvJ inbred+/+
females. There was no difference in TRD between these two types of matings. The expected distribution of litters for a TRD of 0.9 is also shown.(Figure l ) , there was, nevertheless, a significant differ- ence between the observed and expected number of litters (GI41 = 25.81, P
<
0.001-TRD classes 0.0-0.5 were pooled because of low numbers of observations). This difference was due to a large deficiency of litters observed in the 0.8-0.9 TRD class and an excess in the 0.9-1.0 class. The excess in the 0.9-1.0 TRD class may partly reflect the lack of a 0.8-0.9 TRD class for litter sizes of four or less, although these do not account for many litters in total (most litters ranged in size from five to seven). Within each separate litter size class, there was an observed deficiency of litters with TRDs of 0.8-0.9, and an excess with TRDs of 0.9-1.0, sug- gesting that while mean TRD is -0.9, there are proba- bly some t haplotypes with stronger transmission ratios than this.0.3
-I
I
o’2
0. I
11
0
I‘
I I I I 1::
s
8 Pz
0
Number of Offspring per male
FIGURE 2.-Relationship between male transmission ratio and the total number of offspring produced per male. Mean TRD for all males in this study is 0.9 (-
-
-). Solid lines represent the upper and lower 9.5% CIS calculated for a trans- mission ratio of 0.9.TABLE 3
Transmission ratios of + / t males carrying wild type (+) chromosome 17’s from populations where t haplotypes
were present or absent
Population in which wild-type (+)
chromosome I 7 originated Mean TRD“
haplotypes present 0.899 (32)
/ haplotypes absent 0.911 (17)
All males were first generation males bred in the laboratory
“Number of males is shown in parentheses.
The 47 first and second generation wild-derived
+/
t males resulted largely from, and were outbred to, wild- derived+/+
females from different populations with the general aim of surveying a large number of wild chromosomes for potential modifiers of TRD. Many of the females were extracted from populations that did not contain t haplotypes, with the idea that the absence of t haplotypes in these populations might reflect the presence of TRD-reducing “modifier” wild-type chro- mosomes. However, there was n o evidence that the mean TRD for + / t males whose wild-type chromosome17 was derived from a population where t haplotypes were absent, was different from the TRD of males whose
+
chromosomes were derived from populations where t haplotypes were present (Mann-M’hitney U,,,,,, = 263.5, P = 0.858, Table 3). A one-way ANOVA on trans- mission ratios of the individual males comprising thetwo groups in Table 3, also found no significant be-
tween-group variation in TRD levels (I;il.471 = 0.0096,
P = 0.922). Although this survey was by no means a systematic search, chromosomes from some 18 different populations, and a large number of individual mice, were tested. The failure to find evidence of any consis- tent reduction in TRD from any male, or male-female combination, suggests that modifiers were not preva- lent, at least in the populations we surveyed.
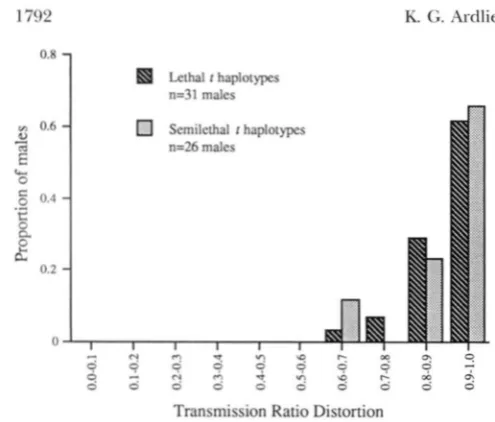
Lethal and smikthal t haplotypPs: While this study has shown generally high levels of TRD in all litters and males tested, previous studies found that semilethal t haplotypes may have lower TRD levels than lethal t h a p
lotypes. To determine whether the presence of a lethal mutation might affect the TRD levels associated with individual t haplotypes, the data for lethal and semile- thal t haplotypes were analyzed separately. The distribu- tion of TRD levels associated with males of each of the
1792 K. G . Ardlie and L. M. Silver
o'8
1
Lethal I haplotypes
n=31 males
VI 0.6
e,
m C
L
0
5
0.4.-
E
ac
e
0.2
0
Transmission Ratio Distortion
Flc;t'KE 3.-Frequency distributions of TRD levels for wild
+ / t males carrying lethal or semilethal haplotypes. Mean TRD of males with lethal t haplotypes is 0.89 and of males
w i t h semilethal t haplotypes is 0.91.
n o significant difference in mean TRD between these two classes (Gill = 1.16, P = 0.282). However, a hetero- geneity Gtest on the unpooled data for all 10 chromo- somes did reveal some significant heterogeneity in TRD among the chromosomes (G,l[s), = 20.98, 0.01
<
P<
0.025). Because there was a higher level of heterogene- ity among the semilethal than lethal chromosomes
when the two classes were analyzed separately (data not shown), this may, in part, reflect the smaller numbers of offspring generated from some of the semilethal
chromosomes (Table 2 and see Figure 2). The TRD levels of the semilethal t haplotypes were not consis- tently lower than those of the lethal t haplotypes, how- ever.
PPE nnd (2; l i t t m : LENINCTON and HEISLER (1991) have reported a difference in the mean levels of TRD associated with litters resulting from cycling estrus (CE) relative to those resulting from postpartum estrus
(PPE). In particular, they found a reduction of t haplo- type transmission to near Mendelian levels (0.5) in PPE litters. To investigate this phenomenon, we compared transmission bias in litters conceived from each of these two estrus classes. We observed high transmission levels in both types of litters (Figure 4). A 2 X 2 contingency test found no difference in the overall numbers of t
haplotype and non-t haplotype carrying offspring pro-
duced in all PPE us. CE litters
(x'II
I = 1.66, P = 0.198, Table 4). Moreover, paired-data from a subset of 26 males that produced both PPE and CE litters with the same female, further confirmed the absence of any dif- ference in transmission bias d u e to the type of c o n c e p tion (one-way ANOVA, F11,501 = 0.644, P = 0.426).As we found no difference in TRD between the PPE a n d CE litters, where an earlier study had found a sig- nificantly lower TRD in the PPE class, we examined the power of our test, and sample sizes, to detect such a
PPE litters
0.6
-
e,
.-
.-
I-
u+
0
8
0.4-
.d
I
8
a
E
0.2
-
0 F1 I
I I
o o o o o o c o ~ o
Transmission Ratio Distortion
FIGURE 4.-Frequency distributions of TRD levels in litters that resulted from cycling estrus (CE) or postpartum estrus (PPE). The type of conception had no effect on the transmis- sion ratio. Mean TRD of males that sired PPE litters is 0.87, and 0.90 for males that sired CE litters.
difference assuming there really was o n e . " h e n t h e null hypothesis is false, the test statistic follows a
noncentral
x'
distribution, with the noncentrality pa-rameter, A, as o n e of its parameters. Using the total numbers of offspring obtained in each class (from Ta-
ble 4) we calculated A for a combination of possible TRD values, and obtained corresponding values of the power
P(A)
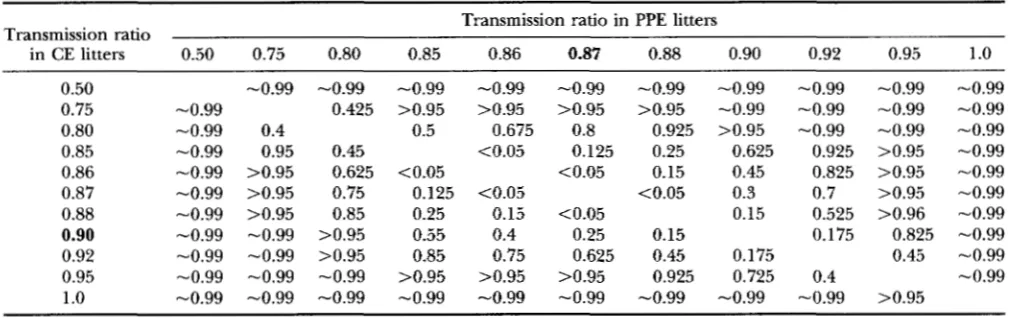
(for a = 0.05 a n d df = 1 ) from tables of the noncentralx*
(FIX 1954). These are shown in Table5. For the TRD level obtained in our CE litters (0.9),
TRD's varying over the range of 0.85-0.92 in the PPE class are associated with low to moderate values of
P
(0.2-0.6). The same is true in reverse for the PPE value of 0.87 we obtained. In contrast, for a TRD of 0.9 in the CE litters, a n d a hypothesized TRD of 0.5 in the PPE litters, we calculated A l l ! = 143.6, which is associ- ated with a value ofP
in excess of 0.99. Thus we would have had a high probability of detecting a difference in TRD this large with our sample sizes, had there really been one.TABLE 4
Transmission ratios of + / t males for litters conceived during postpartum estrus and cycling estrus
Male parent PPE litters CE litters
All males combined
TRD" 0.90 5 0.12 0.89 2 0.16
No. of males 42 33
No. of young 756 297
TRD" 0.89 2 0.14 0.92 2 0.12
No. of males 26 26
No. of voune: 229 178
Males that produced both litters
TRD of t Haplotypes in Wild Mice 1793
TABLE 5
Values of
fl
corresponding to values of A calculated for pairs of transmission ratios for cr = 0.05 and d. f. = 1Transmission ratio Transmission ratio in PPE litters
in CE litters 0.50 0.75 0.80 0.85 0.86 0.87 0.88 0.90 0.92 0.95 1
.o
0.50 -0.99 -0.99 -0.99 -0.99 -0.99 -0.99 -0.99 -0.99 -0.99 -0.99
0.75 -0.99 0.425 >0.95 >0.95 >0.95 >0.95 -0.99 -0.99 -0.99 -0.99
0.80 -0.99 0.4 0.5 0.675 0.8 0.925 >0.95 -0.99 -0.99 -0.99
0.85 -0.99 0.95 0.45 <0.05 0.125 0.25 0.625 0.925 >0.95 -0.99 0.86 -0.99 >0.95 0.625 <0.05 <0.05 0.15 0.45 0.825 >0.95 -0.99 0.87 -0.99 >0.95 0.75 0.125 <0.05 <0.05 0.3 0.7 >0.95 -0.99 0.88 -0.99 >0.95 0.85 0.25 0.15 <0.05 0.15 0.525 >0.96 -0.99 0.90 -0.99 -0.99 >0.95 0.55 0.4 0.25 0.15 0.175 0.825 -0.99
0.92 -0.99 -0.99 >0.95 0.85 0.75 0.625 0.45 0.175 0.45 -0.99
0.95 -0.99 -0.99 -0.99 >0.95 >0.95 >0.95 0.925 0.725 0.4 -0.99 1
.o
-0.99 -0.99 -0.99 -0.99 -0.99 -0.99 -0.99 -0.99 -0.99 >0.95Transmission ratios shown in bold are those obtained in our PPE and CE litters.
Comparison of wildcaught and laboratory-bred litter
sizes: The TRD data are based on litters from both laboratory-bred wild mice and wild-caught mice. Thus litter sizes from these two matings were compared to determine whether the better quality environment of the laboratory might have had an effect on increasing litter size and thus, estimates of TRD. A total of 179 litters were collected as embryos from wild-caught fe- males, and a total of 245 litters were bred in the labora- tory in matings between wild male and wild female mice. Unexpectedly, laboratory-bred litter sizes were smaller on average than wild litter sizes (Mann-Whitney
U,179,2451
= 16,559, P<
0.0001) with mean = 5.2 pups in laboratory-bred litters and mean = 6.5 pups in wild- collected litters. These two classes of litters differed pri- marily with respect to the chromosome 17 genotypes of the parents. Most of the litters collected from the field resulted from matings among two wild-type (+/ +) parents (see above), whereas in contrast, all litters born in the laboratory had a male parent that carried a t haplotype ( + / t ) . The possibility that matings with + / t males might result in reduced litter sizes, either through a reduction in the fertility of + / t heterozygous males or as a reduced viability of + / t embryos has been investigated further, and these findings are the topic of a separate paper.DISCUSSION
t haplotypes derived from wild mice are typically re- ported to have transmission ratios of 20.90 (SILVER 1985; RUVINSKY et al. 1991). Transmission ratios of this order are expected to result in very high frequencies of t-carrying animals in wild populations (260%) based on various stochastic and deterministic models (BRUCK 1957; LEWONTIN and DUNN 1960; LEVIN et al. 1969). However, empirically observed frequencies are typically much lower, in the range of 15-25% karrying ani- mals (PETRAS 1967; LENINGTON et al. 1988; RWNSKY et
al. 1991). All models evince a strong effect of absolute levels of TRD on the frequency of t haplotypes expected in a population. Thus, if the effective levels of TRD in natural populations were lower than previously esti- mated due to the existence of TRD-reducing modifier genes, this could account for the low observed fre- quency of t haplotypes in natural populations (LEVIN et al. 1969). In fact, evidence for genetic modifiers of TRD has been obtained in two studies of t haplotypes main- tained in the laboratory for long periods of time (BEN-
NETT et al. 1983). BENNETT and her colleagues showed
that reduction in TRD was due to both a general effect of genetic background, and more specifically, to the homologous chromosome (GUMMERE et al. 1986).
Previous estimates of TRD in wild mice have all been greatly limited by the lack of a visible phenotype and the need to type the presence of a t haplotype by breed- ing to laboratory stocks carrying the Brachyury ( T ) mu- tation (DUNN 1957a; ANDERSON 1964; BENNETT 1978; LENINCTON and HEISLER 1991). Progeny testing of this kind is often unreliable because wild mice are difficult to breed in the laboratory (SAGE 1981), the diagnostic tailless T / t phenotype is not completely penetrant, and there is always the possibility of overlooking a t-bearing individual when only small numbers of offspring are generated from it. With the availability of molecular genetic markers, it is now possible to demonstrate the existence of a t haplotype directly in both adult and fetal DNA samples. Using t-diagnostic markers, we have obtained data on TRD levels based on matings between entirely wild animals, both in the field and the labora- tory.
1794 K. G . Ardlie and L. M. Silver
was 0.90. Overall, the mean TRD for all wild mice tested was 0.9. Although this represents neither a thorough nor exhaustive survey of genetic modifiers of TRD, ran- dom testing of individual mice derived from 18 inde- pendent populations provided no evidence for an effect of either different genetic backgrounds or the homolo- gous wild-type chromosome 17, in reducing the trans- mission distortion of thaplotypes in the wild. Only 1.8%
of all litters produced by wild mice in the laboratory had TRD levels of 50.5. These litters were not produced consistently by any male, nor by any particular combina- tion of wild male and wild female, nor with any given t haplotype, and thus did not appear related to the genotype of either the male or the female. That no mice showed reduced TRD levels in matings to any wild female indicates a general absence of modifiers of drive among wild mice, at least in the populations we tested, and suggests that they do not play an important role in determining the frequency of t haplotypes in the wild. Additionally, there was no significant difference in transmission ratios obtained with matings to wild-de- rived females as compared to matings with an inbred laboratory strain. This further suggests that there is no special effect of a wild genetic background on reducing TRD levels.
Strong selection to reduce the transmission bias of drive chromosomes is expected, particularly from the homologous chromosome (PROUT et al. 1973; WU and
HAMMER 1990), and there is evidence for the evolution
of suppressors of drive in the SD system in Drosophila melanogaster (HIRAIZUMI and THOMAS 1984; WERREN et al. 1988; HIRAIZUMI et al. 1994). Modifiers of SD are known to be present on many naturally occurring X chromosomes, and distorting chromosomes have been shown to be absent from populations where suppressors are present at high frequencies (HIRAIZUMI and THOMAS 1984). However, there are also examples of
drive systems where suppressors of drive have not been found, such as SR in Drosophila pseudoobscura, where there is a strong expectation that they should be Y- linked (Wu and HAMMER 1990). Possible components of selection acting against SR have been found, such as a reduction in viability and fertility in both sexes, and reduced fecundity of SR homozygous females (CURT-
SINGER and FELDMAN 1980; WU 1983a,b), although,
demonstrating selection acting against drive systems in natural populations in general has been difficult (BECK-
ENBACH 1996). Other forms of selection, such as viabil-
ity differences, have also been demonstrated to be act- ing against SD (WU et al. 1989). Thus there may be many different components of counterselection acting against different drive systems, and their relative impor- tance may depend strongly on the underlying molecu- lar genetics of the system and on the ecological context of the populations under study.
A number of previous studies have found that certain semilethal t haplotypes show lower levels of TRD than
lethal t haplotypes (PETRAS 1967; LENINGTON and HEISLER 1991), and this difference in TRD has typically been assumed when modeling t haplotypes (LEWONTIN 1962;YOUNG 1967; FRANKS and LENINGTON 1986). How- ever, high TRD levels have previously been reported for many other wild-derived semilethal t haplotypes (DUNN
1957a,b; DUNN and BENNETT 1966), and since the fertil- ity effects of t haplotypes are genetically separable from lethality effects (SILVER 1993), there is no a p o r i reason why the two should be correlated in any way. In this study, we found no evidence for a difference in TRD expressed by lethal and semilethal t haplotypes, with both types of chromosomes showing high levels of -0.90.
There was also no difference in TRD between litters that resulted from postpartum or cycling estrus. This contrasts with the findings of LENINGTON and HEISLER
(1991), who found that TRD was significantly lower and, in fact, not different from Mendelian expectations
(0.5) in litters produced as a result of PPE. Although the mechanism underlying this presumed difference is unknown, the authors suggested that PPE might be associated with a delay in the timing of mating, which has previously been shown to reduce transmission dis- tortion (BRADEN 1958, 1972; YANAGISAWA et al. 1961). This effect of delayed mating only occurs with some t haplotypes, however (YANAGISAWA et al. 1961; BRADEN
1972; ERICKSON 1973; GARSIDE and HILLMAN 1989a,b). While both the timing of mating and the environment of the sperm can be shown to result in a significant decrease in TRD for some t haplotypes, this effect is not consistent, and there are a number of t haplotypes that are transmitted at high ratios independent of most other factors (GARSIDE et al. 1991).
LENINGTON and HEISLER (1991) also found that the frequency of PPEconceived litters tended to be higher for matings between individuals derived from the same population than for individuals derived from different populations. As a consequence, t haplotype transmis- sion was effectively lower in within-population (largely PPE) than in between-population (largely CE) matings. Most of the litters produced in our study resulted from between-population matings, and in contrast, tended to be largely PPE-conceived rather than CE-conceived litters. TRD was, nevertheless, very high in these litters, and thus we find no evidence in support of this form of behavioral reduction of TRD in the populations of mice that we sampled.
The data presented here, however, do suggest an- other possible means of behavioral reduction of TRD. In three matings between wild + / t males and
+/
+
females, very low transmission ratios were observed-TRD of t Haplotypes in Wild Mice 1795
nation on other drive systems and in general. HAIG and
BERGSTROM (1995) showed that if competition between driven and nondriven sperm within an ejaculate is bal- anced by reduced competitiveness of the driven sperm in competition between ejaculates, this can lower the probability of an egg being fertilized by a meiotically driven sperm. Multiple mating may also effect the fre- quency of the two male genotypes where sperm dis- placement occurs, if the drive and nondrive genotypes differ in their sperm displacing ability, and/or their resistance to displacement, as has been demonstrated for SR males in D. pseudoobscura (WU 1983a,b).
There is some evidence that the t carrying sperm from
+/
t males may be less functional than the+
sperm from wild type males. Using a low sperm:egg ratio (9:l)assay,JoHNsON et al. (1995) found that
+ / p 5
sperm fertilized only 25% of the eggs fertilized by congenic wild-type+/+
sperm. This is less than the 2 5 0 % ex- pected, since t haplotypes effect TRD by inactivating only their meiotic partners containing+
chromosomes (SEITZ and BENNETT 1985). Moreover, both the mean TRD of 0.17 we obtained here and the value of 0.22obtained in controlled sperm mixing experiments by OLDS-CLARKE and PEITZ (1985), are lower than would be expected for a female copulating with both a
+/+
and a + / t male. An equal mixture of sperm from the two males should contain an effective ratio of -2:l wild- type to t-sperm, yielding 33% + / t offspring (32% for a TRD of 0.9). Neither value is significantly different from the expected value, however, and the sample sizes of both studies are small. This calculation assumes the two sperm types mix equally, which may also not be the case, but whether sperm displacement occurs during multiple mating in mice is unknown. Unlike the SRsystem in D. pseudoobscura, where the nondriving chro- mosome is eliminated, both sperm types are present in the ejaculate of + / t males. However, reduced motility of
+
sperm has been demonstrated (OLDSCLARKE1983, 1989), and this could influence the ability of
+/
t sperm to displace other sperm, or to resist displace- ment.
The absolute number of multiply-mated litters in our study is too low to address what, if any, effect multiple mating may have on the transmission ratios of + / t
males in natural populations. While it may play a role in reducing effective TRD under some conditions, further investigation of both the frequency of multiple matings in natural populations of mice and its specific effect on
t haplotypes are necessary.
The accumulated data from several studies have dem- onstrated an ancient origin for t haplotypes with a diver- gence from wild-type chromosomes > 2 mya (KLEIN et
al. 1985; DELARBRE et al. 1988; HAMMER et al. 1989).
Despite this, contemporary t haplotypes are very closely related, and phylogenetic analyses indicate that they have all descended from a common ancestral chromo- some much more recently, perhaps 100,000-10,000
years ago (WILLISON et al. 1986; MORITA et al. 1992;
HAMMER and SILVER 1993). To reconcile an ancient origin with a recent common ancestor, HAMMER and SILVER (1993) suggested a recent turnover of t haplo- types in response to selection from suppressors or mod- ifiers of drive. If suppressors were prevalent in natural populations, then the persistence o f t haplotypes would depend on recurrent mutation to higher distorting al- leles, forcing a rapid turnover of t haplotypes and per- haps accounting for the accumulation of additional in- versions which locked in new distorter loci (HAMMER and SILVER 1993).
In fact, evidence for possible modifiers of drive has been obtained in analyses of TRD in t haplotypes main- tained in the laboratory, and suppressors of drive may accumulate on the genetic background of laboratory populations where TRD is not continually monitored, and selection for high TRDs has been reduced (GUM-
MERE et al. 1986). However, different t haplotypes ap- pear to be differentially susceptible to reductions in TRD. The level of transmission bias expressed by some
t haplotypes (such as to, t6, and
t"")
has been shown to be affected by a number of factors, including genetic background (OLDSCLARKE and MCCABE 1981; BENNETTet al. 1983; GUMMERE et al. 1986), the timing of mating (BRADEN 1958, 1972), and i n vitro fertilization (MCGRATH and
HILLMAN
1980a,b). These t haplotypes show variable levels of TRD from one male to another. In contrast, other t haplotypes (such as P5) are always transmitted at high ratio's independent of all external factors (YANAGISAWA et al. 1961; GARSIDE and HILLMAN1989a). In at least some instances, the t haplotypes with variable transmissions are missing distortion genes di- rectly (FOX et al. 1985) or may have mutations in their distortion loci (BROWN et al. 1989). Mutations of this kind in wild t haplotypes would be likely to cause a reduction in TRD and result in the rapid loss of these chromosomes from wild populations. Similarly, the spe- cific differences in TRD demonstrated on some, but not all, inbred backgrounds in the laboratory may not be realized in the wild, where there may be strong selec- tion for high levels of drive.
The results of the current study do not necessarily contradict these earlier findings, but they do suggest that while modifiers may well have driven much of the past evolution of the chromosome, they are no longer an active force in suppressing drive in the current con- text of this chromosome in natural populations. Thus, other explanations must be invoked to explain the dis- crepancy between observed and predicted frequencies of t haplotypes in natural populations.
Thanks to D. STRATTON, R. C. LEWONTIN, A. G. CLARK, D. WEIK
REICH, and A. BERRY for help and cornmen& on the manuscript and
to A. BERRY, D. WAGNER, and J. MCDONALD for much appreciated help with the mouse colony on many occasions. This research was
supported by a grant from the National Institutes of Health ( H D
1796 K. G. Ardlie and L. M. Silver
LITERATURE CITED
ANDERSON, P. IC, 1964 Lethal alleles in Mus musculus local distribu- tion and evidence for isolation of demes. Science 145 177-178.
ARDLIE, K. G., and L. M. SILVER, 1996 Recent evolution of mouse t
haplotypes at polymorphic microsatellites associated with the t
complex responder (Tcr) locus. Genet. Res. 67: 1-10.
ARTZT, K., 1984 Gene mapping within the T/t complex of the mouse. 111. &lethal genes are arranged in three clusters on chro- mosome 17. Cell 39: 565-572.
ARTZT, IC, P. MCCORMICK and D. BENNETT, 1982a Gene mapping within the T/tcomplex of the mouse. I. t-lethal genes are nonalle- lic. Cell 2 8 463-470.
ARTZT, R, H.S. SHIN and D. BENNETT, 1982b Gene mapping within the T/tcomplex of the mouse. 11. Anomalous position of the H-
Z complex in &haplotypes. Cell 28: 471-476.
BECKENBACH, A. T., 1996 Selection and the “sex-ratio” polymor- phism in natural populations of Drosophila pseudoobscura. Evolu- tion 50: 787-794.
BENNETT, D., 1975 The T-locus of the mouse. Cell 6: 441-454.
BENNETT, D., 1978 Population genetics of T / t complex mutations, pp. 615-632 in h g z n s of Inbred Mice, edited by H. C. MORSE. Academic Press, New York.
BENNETT, D., A. K. ALTON and K. ARTZT, 1983 Genetic analysis of transmission ratio distortion by t-haplotypes in the mouse. Genet. Res. 41: 29-45.
BRADEN, A. W. H., 1958 Influence of time of mating on the segrega- tion ratio of alleles at the T locus in the house mouse. Nature
181: 786-787.
BRADEN, A. W. H., 1972 T-locus in mice: segregation distortion and sterility in the male, pp. 289-305 in Proc. Int. Symp. on The Genetics
of the Spermatozoon, edited by R. A. BEATTY and GLUECKSHON- WAELSCH. University of Edinburgh, Edinburgh.
BROWN, J., J. A. CEBRA-THOMAS, J. D. BLEIL, P. M. WASSARMAN and L. M. SILVER, 1989 A premature acrosome reaction is pro- grammed by mouse t haplotypes during sperm differentiation and could play a role in transmission ratio distortion. Develop ment 106: 769-773.
BRUCK, D., 1957 Male segregation ratio advantage as a factor in maintaining lethal alleles in wild populations of house mice. Proc. Natl. Acad. Sci. USA 43: 152-158.
CHURCH, G. M., and W. GILBERT, 1984 Genomic sequencing. Proc. Natl. Acad. Sci. USA 81: 1991-1995.
CURTSINGER, J. W., and M. W. FEI.DMAN, 1980 Experimental and the- oretical analysis of the “sex-ratio” polymorphism in Drosophila pseudoobscura. Genetics 94: 445-466.
DEIARBRE, C., Y . KASHI, P. BOURSOT, J.
s.
BECKMANN, P. KOURILSKY et ab, 1988 Phylogenetic distribution in the genus Mus of t- complex-specific DNA and protein markers: inferences o n the origin of t-haplotypes. Mol. Biol. Evol. 5: 120-33.DUNN, L. C., 1957a Evidence of evolutionary forces leading to the spread of lethal genes in wild populations of house mice. Proc. Natl. Acad. Sci. USA 4 3 158-163.
DUNN, L. C., 195713 Studies of the genetic variability in populations of wild house mice. 11. Analysis of eight additional alleles at locus
T. Genetics 42: 299-311.
DUNN, L. C., and D. BENNETT, 1966 Report on viability. Mouse News Lett. 34: 20-22.
DUNN, L. C., and J. SUCKLING, 1956 Studies on the genetic variability in wild populations of house mice. I. Analysis of seven alleles at locus T. Genetics 41: 344-352.
ERICKSON, R. P., 1973 Haploid gene expression versus meiotic drive: the relevance of intercellular bridges during spermatogenesis. Nature New Biol. 243: 210-212.
FEINBERG, A. P., and B. VOGELSTEIN, 1984 A technique for radiola- beling DNA restriction endonuclease fragments to high specific activity. Anal. Biochem. 137: 266-267.
FIX, E., 1954 Tables of noncentral x2, pp. 15-19 in University of
Calfomia Publications in Statistics, edited by G. M. KUZNETS, E. L. LEHMANN, M. M. LOVE, J. NEYMAN, 0. STRUVE et al. University of
California Press, Berkeley.
Fox, H. S., G. R. MARTIN, M. F. LYON, B. HERRMANN, A.-M. FRISCHAUF et al., 1985 Molecular probes define different regions of the mouse t complex. Cell 4 0 63-69.
FRANKS, P., and S. LENINGTON, 1986 Dominance and reproductive behavior ofwild house mice in a seminatural environment corre-
lated with T-locus genotype. Behav. Ecol. Sociobiol. 18: 395- 404.
GARSIDE, W., and N. HILLMAN, 1989a The in vivo and in vitro trans- mission frequencies of the tw5 -haplotype in mice. Genet. Res.
GARSIDE, W., and N. HILLMAN, 1989b The transmission ratio distor- tion of the thahaplotype in vivo and in vitro. Genet. Res. 53: 25- 8.
GARSIDE, W., C. RUANGVORAVAT, P. DOLAN and N. HILLMAN, 1991
The in vivo and in vitro transmission ratio distortion of one com- plete and two partial t haplotypes in mice. Genet. Res. 57: 153-
158.
GUMMERE, G. R., P. J. MCCORMICK and D. BENNETT, 1986 The in- fluence of genetic background and the homologous chromo- some 17 on &haplotype transmission ratio distortion in mice. Genetics 114: 235-245.
HAIG, D., and C. T. BERGSTROM, 1995 Multiple mating, sperm com- petition and meiotic drive. J. Evol. Biol. 8 265-282.
HAMMER, M. F., and L. M. SILVER, 1993 Phylogenetic analysis of the alpha-globin pseudogene4 (Hba9s4) locus in the house mouse species complex reveals a stepwise evolution o f t haplotypes. Mol. Biol. Evol. 10: 971-1001.
HAMMER, M. F., J. SCHIMENTI and L. M. SILVER, 1989 Evolution of mouse chromosome 17 and the origin of inversions associated with t haplotypes. Proc. Natl. Acad. Sci. USA 86: 3261-3265.
HERRMANN, B., M. B CAN, P. E. MAINS, A,”. FRISCHAUF, L. M. SILVER et al., 1986 Genetic analysis of the proximal portion of the mouse t complex: evidence for a second inversion within t haplo- types. Cell 44: 469-476.
HIRAIZUMI,~., and A. M. THOMAS, 1984 Suppressor system of Segre- gation Distortion (SD) chromosomes in natural populations of Drosophila melanogmter. Genetics 9 5 693-706.
HIRAIZUMI, Y., J. M. ALBRACHT and B. C. ALBRACHT, 1994 X-Linked elements associated with negative segregation distortion in the SD system of Drosophila melanogmter. Genetics 138: 145-152.
HOGAN, B., R. BEDDINGTON, F. COSTANTINI and E. LACY, 1994 Manipu- lating the Mouse Embryo: A Labwatoly Manual. Ed. 2. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
JOHNSON, L. R., S. H. PII.DER, J. L. BAILEY and P. OLDSCLARKE, 1995
Sperm from mice carrying one or two t haplotypes are deficient in investment and oocyte penetration. Dev. Biol. 168: 138-149.
KLEIN, J., D. NIZETIC, M. GOLUBIC, Z. DEMBIC and F. FIGUEROA, 1985
Evolution of the H-2 genes on t chromosomes, pp. 97-106 in Cell Biology of the Major Histocornpatability complex, edited by B. PERNIS and H. J. VOGEL. Academic Press, New York.
KLEIN, J., P. SIPOS and F. FIGUEROA, 1984 Polymorphism of &om- plex genes in European wild mice. Genet. Res. 44: 39-46.
LENINGTON, S., P. FRANKS and J. WILLIAMS, 1988 Distribution of t haplotypes in natural populations of wild house mice. J. Mammal.
LENINGTON, S., and I. L. HEISLER, 1991 Behavioral reduction in the transmission of deleterious t haplotypes by wild house mice. Am.
Nat. 137: 366-378.
LEVIN, B. R., M. L. PETRAS and D. I. RASMUSSEN, 1969 The effect of migration on the maintenance of a lethal polymorphism in the house mouse. Am. Nat. 103: 647-661.
LEWONTIN, R. C., 1962 Interdeme selection controlling a polymor- phism in the house mouse. Am. Nat. 96: 65-78.
LEWONTIN, R. C., and L. C. DUNN, 1960 The evolutionary dynamics
of a polymorphism in the house mouse. Genetics 45: 705-722.
LYON, M. F., 1986 Male sterility of the mouse tcomplex is due to homozygosity of the distorter genes. Cell 44: 357-363.
MCGRATH, J., and N. HILLMAN, 1980a The in vitro transmission fre- quency of the t6 allele. Nature 283: 479-481.
MCGRATH, J., and N. HILLMAN, 1980h The in vitro transmission fre- quency of the t12 mutation in the mouse. J. Emhryol. Exp. Mor-
MORITA, T., H. KUBOTA, K. MURATA, M. NOW, C. DEIARBRE et al.,
1992 Evolution of the mouse t haplotype: Recent and world- wide introgression to Mus musculus. Proc. Natl. Acad. Sci. USA
NUNNEY, L., and A. E. M. BAKER, 1993 The role of deme size, repro- ductive patterns, and dispersal in the dynamics of t-lethal haplo- types. Evolution 47: 1342-1359.
OLDSCLARKE, P., 1983 The nonprogressive motility of sperm popu- lations from mice with a tw3Z haplotype. J. Androl. 4 136-143.
53: 21-24.
69: 489-499.
phol. 60: 141-151.
TRD of t Haplotypes in Wild Mice 1797
OLDSCLARKE, P., 1989 Sperm from tw32/+ mice: capacitation is normal, but hyperactivation is premature and non-hyperacti- vated sperm are slow. Dev. Biol. 131: 475-482.
OLDSCLARKE, P., and S. McCABE, 1981 Genetic background affects expression of t haplotype in mouse sperm. Genet. Res. 4 0 249-
254.
OLDS-CLARKE, P., and B. PEITZ, 1985 Fertility of sperm from t/+ mice: evidence that +-bearing sperm are dysfunctional. Genet. Res. 47: 49-52.
PETRAS, M. L., 1967 Studies of natural populations of Mus. 11. Poly- morphism at the t locus. Evolution 21: 466-478.
PROUT, T., J. BUNDGARRD and S. BRYANT, 1973 Population genetics of modifiers of meiotic drive 1. The solution of a special case and some general implications. Theor. Popul. Biol. 4: 446-465.
ROHME, D., H. Fox, B. HERRMANN, A-M. FRISCHAUF, J.-E. EDSTROM et al., 1984 Molecular clones of the mouse t complex derived from microdissected metaphase chromosomes. Cell 36: 783-788.
RWINSKY, A,, A. POLYAKOV, A. AGULNIK, H. TICHY, F. FIGUEROA et al.,
1991 Low diversity of t haplotypes in the eastern form of the house mouse, Mus musculus L. Genetics 127: 161.
SAGE, R. D., 1981 Wild mice, pp. 40-90 in T h Mouse in Biomedical Research, edited by H. L. FOSTER, J. D. SMALL and J. G. FOX. Aca- demic Press, New York.
SARVETNICK, N., H. Fox, E. MANN, P. MAINS, R. ELLIOTT et al., 1986
Nonhomologous pairing in mice heterozygous for a t haplotype can produce recombinant chromosomes with duplications and deletions. Genetics 1 1 3 723-734.
SCHIMENTI, J., L. VOLD, D. SOCOLOW and L. M. SILVER, 1987 An unstable family of large DNA elements in the center of the mouse
t complex. J. Mol. Biol. 194 583-594.
SEITZ, A. W., and D. BENNEIT, 1985 Transmission distortion of t-
haplotypes is due to interactions between meiotic partners. Na- ture 313: 143-144.
SILVER, L. M., 1985 Mouse thaplotypes. Annu. Rev. Genet. 1 9 179- 208.
SILVER, L. M., 1993 The peculiar journey of a selfish chromosome: mouse t haplotypes and meiotic drive. Trends Genet. 9 250- 254.
WERREN, J. H., U. NUR, and C.4. Wu, 1988 Selfish genetic elements. Trends Ecol. Evol. 3 297-302.
WILLISON, K. R., K. DUDLEY and J. POTTER, 1986 Molecular cloning and sequence analysis of a haploid expressed gene encoding t
complex polypeptide-1. Cell. 44: 727-738.
Wu, C.-I., 1983a Virility deficiency and the sex-ratio trait in Drosoph-
ila pseudoobscura. I. Sperm displacement and sexual selection. Genetics 105: 651-662.
Wu, C.4, 1983b Virility deficiency and the sex-ratio trait in Drosoph-
ilapseudoobscura. 11. Multiple mating and overall virility selection. Genetics 105: 663-679.
Wu, C.-I., and M. HAMMER, 1990 Molecular evolution of ultra-selfish genes of meiotic drive systems, pp. 177-203 in Evolution at the
Molecular Level, edited by R. K. SELANDER, A. G. CLARK and T. S. WHITTAM. Sinauer, New York.
Wu, C.-I., J. R. TRUE and N. JOHNSON, 1989 Fitness reduction associ- ated with the deletion of a satellite DNA array. Nature 341: 248-
251.
YANAGISAWA, K, L. C. DUNN and D. BENNETT, 1961 On the mecha- nism of abnormal transmission ratio at the T locus in the house mouse. Genetics 4 6 1635-1644.
YOUNG, S. S. Y., 1967 A proposition on the population dynamics of the sterile t alleles in the house mouse. Evolution 21: 190-198. ZAR, J. H., 1996 Biostatistical Analysis. Prentice-Hall, Inc., NJ.