Predictive Motor Learning of
Ob j ect Manipulation
Alice Geraldine Witney
Sobell Departm ent of Neurophysiology,
Institute of Neurology,
University College London
Submitted for examination for the PhD degree: November, 2000
Viva: 9th February, 2001
All rights reserved
INFORMATION TO ALL USERS
The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest U643088
Published by ProQuest LLC(2015). Copyright of the Dissertation is held by the Author.
All rights reserved.
This work is protected against unauthorized copying under Title 17, United States Code. Microform Edition © ProQuest LLC.
ProQuest LLC
789 East Eisenhower Parkway P.O. Box 1346
Abstract
Anticipating the consequences of our m otor com m ands is a fundam en
tal component of sensorimotor control. For example, w hen one hand pulls
on an object held in the other hand, there is an anticipatory increase in grip
force in the restraining hand that prevents the object from slipping. This an
ticipation is thought to rely on a forward internal m odel of the m anipulated
object and the m otor system, enabling the prediction of the consequences
of our actions. This thesis examines the learning of prediction using a vir
tual object paradigm . Subject's held an object in each han d whose prop
erties were under computer control. This allowed instant changes in the
behaviour of the objects being m anipulated on a trial to trial basis, w ithout
providing any cues to the subject. To investigate the developm ent of the
prediction that a single object is being m anipulated betw een the hands, the
forces on each object were controlled so that the two objects were linked
together, and therefore acted as a single object. Decay of prediction was ex
am ined after the linkage betw een the forces on the two objects was removed
and they again acted as two independent objects. Prediction of single ob
ject m anipulation was found to be quick to build up after the occurrence of
a linkage betw een the objects, but slow to decay after this linkage was re
moved. The effect of previous experience of either linkage or independence
betw een the objects was then examined, w ith as systematic effect of link
age being found on the current predictive response. W hen the properties of
the virtual object were altered to form objects w ith novel spatial properties,
prediction was found to be specific to the direction that the subject had pre
viously experienced. The learning of anticipatory grip force m odulation to a
virtual object w ith novel properties w as examined by the addition of a tem
poral delay in the linkage between action and effect. A second prediction
developed coupled with the suppression of the earlier predictive response.
These experimental findings have been related to the presence of forward
internal models, and compared with the conditioning of a predictive motor
I w ould like to thank my supervisor. Dr Daniel W olpert for giving me the
opportunity to do this project, and for his help, enthusiasm , and guidance
throughout the three years.
Thanks to Dr Susan Goodbody for her help.
The model in Chapter 6 was done by Phillip Vetter, and I w ould like to
thank him for this.
Thanks to Chris Seers, Bill Cameron for their technical support.
Thanks to Pierre Baraduc for help w ith ETpXand provision of fine cheeses;
Julie Savvides for extensive photocopying and Ana Caballero for translat
ing the experimental protocol into Spanish.
Many thanks to the Wolpert-lab and Sobell D epartm ent for making my
time in the departm ent enjoyable.
Thanks to Joan Cinnelly of the National Hospital for securing cheap ac
commodation for me for three years.
Contents
1 Introduction 13
1.1 Internal m o d e l s ...13
1 . 2 Uses of forward m o d e l s ...16
1.2.1 Sensory p r e d ic tio n ... 22
1.2.2 Distal supervised le a rn in g ... 24
1.2.3 Mental p r a c tic e ...25
1.2.4 M O SA IC ...26
1.3 Learning and representation of an internal m odel ...27
1.4 Grip force as a paradigm for forward m o d e l s ...29
1.4.1 Reactive grip force m o d u la tio n ... 32
1.4.2 Development of grip force r e s p o n s e ... 34
1.4.3 Learning the appropriate grip force re s p o n s e ...36
1.4.4 Bio-mechanics of grip f o r c e ... 38
1.4.5 Cutaneous afferents ... 38
1.4.6 S l i p ...40
1.4.7 Central c o n t r o l ... 41
1.4.8 Cortical a r e a s ... 43
1.4.9 Anticipatory grip fo rc e ... 44
1.4.10 Earlier theories of predictive grip force m odulation . . 45
1.5 Aims of t h e s i s ...47
2.1.2 Grip s u r f a c e s ... 52
2.1.3 Force tra n s d u c e rs... 52
2 . 2 A n a l y s i s ... 54
2.2 . 1 Calculation of grip f o r c e ... 54
2.2.2 Calculation of load f o r c e ... 54
2.2.3 Am plitude and t im i n g ... 54
3 Learning and decay of prediction in object manipulation 55 3.1 Abstract ... 55
3.2 In tro d u c tio n ...56
3.3 M e t h o d s ...58
3.3.1 Subjects ... 58
3.3.2 P r o c e d u r e ... 58
3.3.3 One link c o n d itio n ... 59
3.3.4 Three link c o n d it i o n ... 60
3.3.5 Interval c o n d itio n ... 60
3.3.6 Non-physical c o n d it i o n ... 60
3.4 A n a l y s i s ...61
6
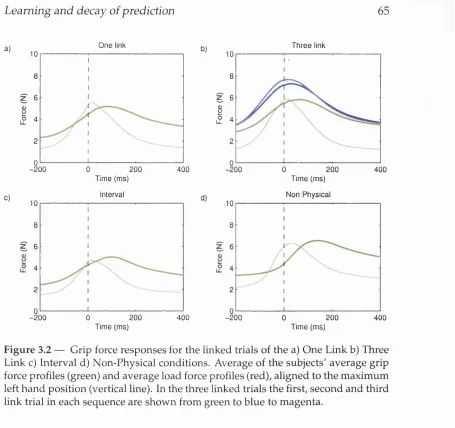
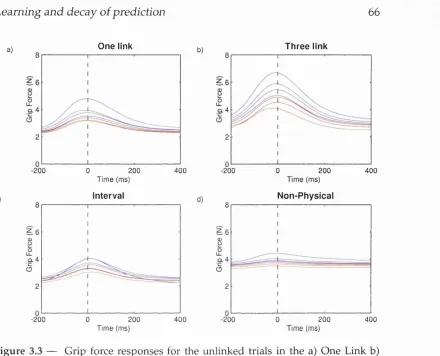
3.5.1 One linked co n d itio n ... 63
3.5.2 Three linked c o n d itio n ... 69
3.5.3 Interval co n d itio n ... 71
3.5.4 Non-physical c o n d itio n ...72
3.6 D isc u ss io n ...73
3.6.1 Development of p r e d ic tio n ...75
3.6.2 Decay of p red ictio n ... 76
3.6.3 Importance of p h y s i c s ...77
4 Predictive motor learning of a temporal delay 78 4.1 Abstract ...78
4.2 In tro d u c tio n ...78
4.3 M e t h o d s ...82
4.3.1 Subjects ... 82
4.3.2 A p p a r a tu s ... 82
4.3.3 P r o c e d u r e ... 83
4.3.4 Non-cued c o n d it i o n ... 84
4.3.5 Self-generated c o n d itio n ...84
4.5.1 Pre-exposure ...8 8
4.5.2 Self-generated c o n d itio n ... 91
4.5.3 Cued c o n d itio n ... 95
4.6 D is c u s s io n ...98
4.6.1 Performance in the absence of a d e l a y ...99
4.6.2 Performance in the presence of a d e l a y ...102
4.6.3 Modularity and internal m o d e l s ... 104
5 The effect of externally generated and novel consequences on pre diction 106 5.1 Abstract ... 106
5.2 In tro d u c tio n ... 107
5.3 M e t h o d s ... 109
5.3.1 P r o c e d u r e ...I l l 5.4 A n a l y s i s ...1 1 2 5.5 R e s u l t s ... 114
5.5.1 Condition LU:- L inked-U nlinked... 114
5.5.2 Condition LE:-Linked-Extem al... 115
5.5.4 Condition D ecay-External... 117
5.5.5 Condition D eca y -N o v e l... 122
5.6 D isc u s s io n ... 125
6 The effect of previous experience and prior expectation on predic tion 129 6.1 Abstract ... 129
6.2 In tro d u c tio n ... 129
6.3 M e t h o d s ... 132
6.4 R e s u l t s ... 135
6.5 D isc u ss io n ... 141
7 Spatial representation of a forward model 145 7.1 Abstract ... 145
7.2 In tro d u c tio n ... 145
7.3 M e t h o d s ... 147
7.4 A n a l y s i s ... 149
7.5 R e s u l t s ... 152
7.5.1 L ^ jjc o n d itio n ... 152
7.5.2 Z/Q c o n d itio n ... 153
7.5.5 LqL iso condition ... 158
7.6 D is c u s s io n ... 161
8 Discussion 167 8.1 Grip force and forward m o d e l s ...167
8.1.1 Development and decay of a forward m o d e l ... 168
8.1.2 M odularity ... 169
8.1.3 The effect of previous ex p erien ce...174
8.1.4 Spatial representation of a forward m o d e l... 175
8.2 Comparison with classical c o n d itio n in g ...176
8.2.1 A c q u is itio n ... 177
8.2.2 Re-acquisition and E x tin ctio n ...178
8.2.3 The Effect of other s tim u li...178
8.2.4 N eural substrates of predictive m otor le a r n in g 179 8.3 S u m m a ry ... 180
10
List of Figures
1.1 Grip force ... 30
1.2 Internal model for predictive grip force m o d u la tio n ... 32
2 . 1 Schematic diagram of the virtual object ... 51
2.2 Bandwidth of the virtual object a p p a r a tu s ... 53
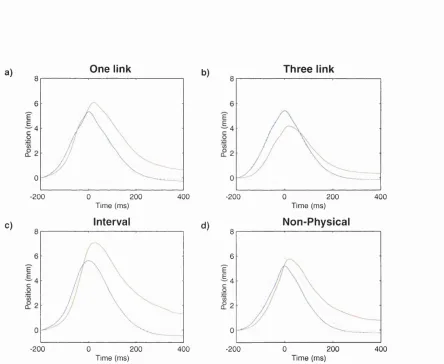
3.1 H and positions ... 64
3.2 Grip force responses for linked t r i a l s ...65
3.3 Grip force responses for unlinked trials ...6 6 3.4 Individual grip force profiles ... 67
3.5 Grip force m odulation over linked and unlinked tr ia ls 6 8 3.6 Grip force modulation over batches ...70
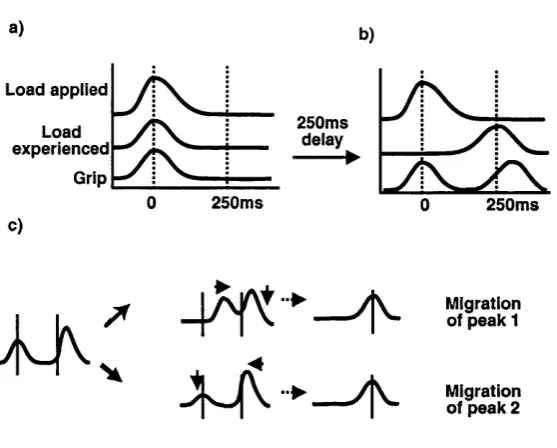
4.1 Hypotheses of possible adaptation to d e l a y ...80
4.2 Schematic diagram of apparatus ...83
4.3 Left hand p o s itio n s ... 89
4.4 Grip response in pre-exposure p h a s e ...91
4.5 Grip force m odulation in self-generated c o n d itio n ...92
4.6 Grip force response for individual subjects ...93
4.7 Coefficient of determination of grip w ith load f o r c e ...94
4.9 Grip force m odulation in cued co n d itio n ...97
5.1 Individual subject's grip force modulation dependent on pre vious trial ... 116
5.2 Grip profiles dependent on previous t r i a l ... 118
5.3 Grip force profiles dependent on occurrence of an external trial 121 5.4 Peak grip force dependent on occurrence of an external . . . . 122
5.5 Grip force profilesdinked, unlinked, l in k e d - d o w n ... 123
5.6 Grip force m odulation dependent on occurrence of a linked- dow n trial ... 124
6.1 Grip force m odulation on linked and unlinked t r i a l s ... 136
6.2 Unlinked grip force p r o f ile s ...138
6.3 Binary tree of grip force modulation and sim ulated data . . . 140
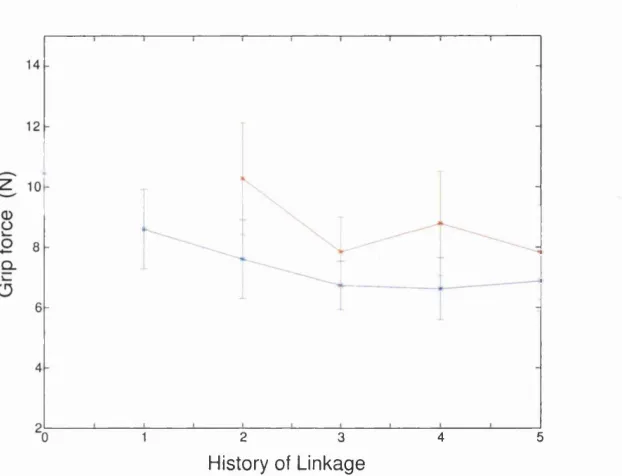
6.4 Regression param eters of the history of lin k a g e ... 141
7.1 Paradigm for studying generalization of predictive grip force 150 7.2 Average load force trajectory ...152
7.3 Grip force modulation: Condition ...153
7.4 Grip force profiles during training p h a s e ...154
7.5 Grip force modulation: condition Lq ...155
7.6 Grip force m odulation on first test t r ia l...156
12
7.8 Grip force modulation: condition LoC/ 4 5 ... 157
7.9 Individual subject's grip force modulation: condition Lot/ 4 5 . . 158
7.10 Grip force modulation: condition L0L4 5 ... 159
7.11 Individual subject's grip force modulation: Condition L0L4 5 . 159
7.12 Grip force modulation: condition LqLi8q ... 160
7.13 Individual subject's grip force modulation: condition L0L1 8 0 . 161
1
Introduction
We are able to move and interact w ith our environm ent effortlessly. H ow
ever, the task of movement and control is deceptively difficult. The simplest
action requires the coordination of m any muscles and joints. Additionally,
during m otor behaviour the problems of coordination, tim ing and interac
tion between neural, muscular and skeletal structures have to be overcome.
The concept of a motor program to successfully overcome such problems
of coordinating movement has become prevalent (Pew, 1974; Rosenbaum,
1980; Jordan, 1995; Jeannerod, 1988)
1.1 Internal models
An early formulation of the presence of a representation of the m otor system
has existed since H ead's (1926) notion of a schema. Such a representation
is thought crucial for the formation of a m otor program. One of the early
definitions of the motor program was by Keele (1968) w ho stated that the
motor program was "a set of muscle commands that are structured before a
Introduction 14
out
However, such early conceptualizations of internal representations were
vague, although an im portant development of the concept of a motor pro
gram was m ade by Held (1962,1965). He suggested that the internal refer
ence of position during goal-directed movements should be derived from
the program of these movements as a corollary of the m otor output, inde
pendent from peripheral feedback. This model was based on the effects of
visuomotor adaptation, in situations where the relation betw een the posi
tion of a visual target in space, and the direction of m ovem ents towards
that target was systematically modified by the use of laterally displacing
prisms. Held argued that in order for the movements to become accurate in
this condition, their program had to be modified on a trial to trial basis, and
rebuilt according to the new visuomotor rules.
A theoretical framework for models of motor representation was pro
vided by M arr's (1982) levels of functioning for information-processing de
vices. Three levels of functioning were proposed, the highest level being
the "computational theory" which defines the goal of the computation, and
the strategy that could be used to carry it out. Second, the "representation
level", where the computational theory is im plemented into a representa
tion of the input and output of the system and into an algorithm for trans
forming input to output. And finally the "hardware im plem entation" of the
representation and the algorithm. This thesis will examine the "representa
tion level" of predictive control.
sys-tern, whose inputs are the motor commands from the 'controller' w ithin the
CNS and outputs are sensory feedback. Knowledge of just the input to a
dynamical system is not enough to predict the output. However, w hen the
'state' of the system is known in addition to the m otor command, the be
haviour of the system can be predicted. The state therefore contains all the
relevant time-varying information needed to predict or control the future of
the system. For example, to predict the effects of applying a torque around
the knee joint, it is also necessary to know the configuration and m otion of
other body segments. One of the problems the m otor system has to over
come is how to estimate the state from the m otor com m and and sensory
feedback.
Representations of the sensori-motor transformations betw een m otor and
sensory variables are thought to occur within the CNS. Such internal m od
els are systems that simulate the behaviour of natural processes and trans
form sensory signals to motor commands and m otor com m ands to sensory
signals. There are two main groups of internal models, know n as forward
and inverse models (Jordan, 1995; Wolpert, 1997; Wolpert and Ghahram ani,
2000).
The forward model is a causal representation of the m otor apparatus
that represents the behaviour of the motor system in response to outgoing
motor commands (Kawato et al., 1987; Jordan and Rumelhart, 1992; Jordan,
1995; Wolpert et al., 1995; Miall and Wolpert, 1996; Wolpert, 1997). That is,
forward models capture the causal relationship betw een actions an d out
Introduction 16
1950; V. Holst, 1954; Jeannerod et al,, 1979).
Inverse models simulate the transformation from outputs to inputs. Un
like the forward model, the inverse model is not necessarily unique as the
transformation can be one-to-many. Fast feed-forward control can be achieved
through an inverse model of the controlled object, and therefore, an accu
rate inverse model could act as ideal feed-forward controllers. Due to this,
the training of inverse models is crucial for performance in feed-forward
control.
1.2 Uses of forward models
Forward models can be separated into forward dynamic models and for
ward sensory models according to their role; forward dynamic models are
responsible for the prediction of the next state given the current state and
motor command; whereas forward sensory models predict the sensory feed
back given the estimated state.
A forward model, given it simulates the motor-sensory transform ation
can have a num ber of potential fundam ental roles in the m otor system, in
cluding motor planning, execution and learning (Kawato et al., 1987; Jordan
and Rumelhart, 1992; Jordan, 1995; Wolpert et al., 1995; Miall and Wolpert,
1996; Wolpert, 1997). The importance of a forward m odel to the m otor sys
tem arises in several areas including state estimation; state prediction; sen
sory prediction; distal supervised learning; mental practice and m odular
State estim ation
The State of the motor system forms a compact representation w ith far lower
dimensionality than the full array of sensory feedback. However, as the
state of a system is not directly accessible to the controller, the state has to
be estim ated using the only sources of information available to the CNS,
sensory feedback and the motor commands. A forward m odel is an impor
tant component in the sensori-motor integration process that occurs during
state estimation. An estimate of the current state is m ade by m onitoring the
inputs, that is the motor commands, and the outputs, the sensory feedback
using an 'observer' (Merfeld et al., 1993; Jordan, 1995; Wolpert et al., 1995).
A Kalman filter m odel can be used to estimate the state. The Kalman fil
ter produces this estimate using both motor outflow and sensory feedback
combined w ith a model of the motor system. The Kalman filter assumes
that there is both variability in the response of the m otor system to the mo
tor command, and variability in the sensory feedback. The Kalman filter
uses two components to try and reduce the effects of these variability on
the estimate of the state. The feed-forward component of the m odel uses
the efference copy together with the current state estimate to predict the
next state by simulating the movement dynamics w ith a forw ard model.
The feedback component compares the sensory inflow w ith a prediction of
the sensory inflow based on the current state. This difference betw een the
actual and predicted sensory feedback, the sensory error is used to correct
the state estimate m ade by the forward model. The contributions of both the
mod-Introduction 18
ulated by the Kalman gain so that optimal state estimates are produced.
Damage to the parietal cortex has been show n to result in the failure
to m aintain state estimates; a patient with a parietal lesion was show n to
experience perceptual fading (Wolpert et al., 1998a).
State prediction
Forward models are im portant for coordination and tim ing in the m otor
system. Prediction from a forward model can help to overcome the prob
lems associated with feedback delays. In sensori-motor loops, feedback de
lays are large in the order of 100 ms (Cordo et al., 1994). If rapid m ovements
are attem pted under feedback control instability could result (Miall et al.,
1993). However, a forward model is able to provide internal feedback of the
predicted consequences of an action and instability can be prevented. If an
efference copy of the actual motor command is input to the forw ard model,
the forward model predicts the feedback. The neural com putation time for
this prediction is much shorter than the external visual or proprioceptive
feedback delay. Therefore, if a forward model is used in the internal feed
back loop, feedback control performance is improved as the large external
feedback delays can be avoided (Ito, 1970). This has been show n w ithin a
Smith predictor model where a forward model is used to provide internal
feedback of the predicted outcome of an action before sensory feedback is
available (Miall et al., 1993).
Forward models are thought to enable predictive responses that can help
the action of one part of the body on another (Massion, 1992; Lacquaniti,
1992; Nashner, 1982; Gahery and Massion, 1981). Different responses are
m ade to self-produced and externally produced perturbations in unload
ing, ball catching and other postural adjustments (Dufosse et al., 1985; Mas
sion, 1992). The predictive response is thought to be dependent on predict
ing the consequences of the descending motor comm and, as signalled by
efference copy, using an internal model of both one's ow n body and the
object (v. Helmholtz, 1867; Sperry, 1950; v. Holst, 1954; Wolpert, 1997).
Performance of a voluntary movement by one body segment is usu
ally accompanied by an adjustment of posture aim ed at preventing the im
balance which w ould otherwise occur. Such anticipatory postural adjust
m ents have been demonstrated to occur in the back and legs w hen a sub
ject is standing and makes a rapid arm m ovem ent affecting the equilib
rium (Bouisset and Zattara, 1987; Cordo and Nasher, 1982; Lee et al., 1987)
The "waiter task" is an example of the use of a forw ard m odel to m ini
mize postural disturbances. The waiter task illustrates how a waiter is able
to unload plates and bottles from a tray with one han d while the tray is
balanced on the contralateral forearm. In this situation the position of the
tray does not move, although unloading the tray should provoke an u p
w ard arm movement. Such adjustments of the limb compliance dependent
on the environmental context are a feature of our m otor system (Lacquaniti
et al., 1992).
The waiter task has been examined extensively experimentally (Dufosse
Mas-Introduction 20
sion, 1992; Lum et al., 1992). In these unloading tasks, one arm is held in
a horizontal position whilst supporting a 1 kg weight. This weight is u n
loaded either by the experim enter's hand (imposed unloading) or by the
subject's other hand in response to a tone (voluntary unloading). The EMC
activity from the brachioradialis of the postural arm, and from the biceps of
the active arm are recorded (Hugon et al., 1982). This enabled a comparison
of the latency of the EMC from the postural arm and from the active arm,
and therefore a measure of the anticipatory nature of the response. The
degree of displacement of the postural forearm is a further m easure used
to determine w hether a predictive response has occurred. Minimal distur
bances reflect that a prediction of unloading was available whilst larger dis
turbances suggest anticipation was not possible.
The position of the w eight-supporting forearm w as found to change
very little w hen the weight is taken off by the contralateral hand. EMC
recordings have shown that an inhibition in the brachioradialis muscle pre
cedes the onset of the change in force caused by the opposite hand. This
indicates that an anticipatory postural adjustm ent occurred prior to the u n
loading so that the position of the loaded hand remains unchanged (Du
fosse et al., 1985; Massion, 1992). W hen the loading w as self-generated,
forearm displacement was found to be 5° ± 2°. In contrast, if the unloading
w as carried out by the experimenter, there was no anticipatory inhibition
of the postural forearm flexors and w hen unloading did occur there was an
upw ard rotation of the postural forearm by 16° ± 2° (Hugon et al., 1982; For
by a w arning tone at a fixed interval before removal, there was still no an
ticipatory response in the forearm muscles. Therefore, self-generated move
m ent was crucial for the anticipation to occur. Further studies demonstrate
that not every active movement is able to result in an anticipatory adjust
ment. If the subject simply pressed a button with their thum b to remove the
load, anticipation did not occur even after several h u n d red presentations
of the situation, and the forearm displacement was large at 20° ± 2° (Du
fosse et al., 1985). Additionally, the anticipatory response did not transfer
between limbs (Ioffe et al., 1996). If subjects were first trained on an un
loading task with a right active arm and a left postural arm and switched
on testing to the right arm being the postural arm, no anticipatory response
was found to occur. However, w hen the leg rather than the contralateral
arm was used as the postural limb in the unloading task, an anticipatory re
sponse occurred in the quadriceps, and postural disturbance was minimal
at 6° ± 5° (Forget and Lamarre, 1995)
Further studies have examined the affect of an additional tem poral de
lay of 100-150 ms between the time between the subject's lifting w ith one
hand and unloading with the other. Subjects were able to leam to predict
this novel situation and anticipation became appropriately tim ed for the
forthcoming unloading (Forget and Lamarre, 1995).
Together these findings suggest that the requirements for a predictive
response are specific.
A further example of the use of forward models is dem onstrated in ball
Introduction 22
1987; Lacquaniti and Maioli, 1989). W hen w e catch a bail that falls ver
tically to the hand, both predictive and reactive mechanisms are used to
produce an appropriate catching action. The forw ard m odel needs to pre
dict time, location and m om entum of the impact so that limb kinematics
and kinetics can be appropriately controlled. This control of limb position
and compliance has to adhere with the timing requirem ents of the task for
catching to be successful. EMC recordings show ed there were early and late
components, the early component is thought to be related to anticipation.
The am plitude of this early EMG response was inversely proportional to the
height of the fall and is thought to correspond to the readiness reaction that
incorporates the fall duration. The role of prediction has been examined by
modification of different variables in the task, for example the height of fall
and mass of ball. W hen balls of identical external appearance were used but
different masses were used, and the mass of ball is unexpectedly changed,
subjects scale their response to the expected m om entum . The prediction
used is thought to be based on a model of the ball's flight combined w ith an
internal model of the current position and m ovem ent of the arm. This for
w ard model is accessed to predict a target, but is u p d ated using peripheral
information from vision to produce an adapted response.
1.2 . 1 Sensory prediction
Cancelling of sensory reafference is essential in the distinction of self-generated
and external-generated disturbances. The apparent disregard for self-produced
ac-tions caused by ourselves to be reduced Qeannerod, 1988).
The cancellation of sensory reafference is necessary in the distinction of
m ovem ent of the world from movement made by our ow n m otor system.
This was described in eye movements by Von Holst and M ittelstaedt (1950)
who defined the role of the efference copy of the m otor command. In their
theory, the role of the efference copy is to cancel undesirable sensory mes
sages resulting from self-generated movements.
This ability to distinguish self from other has been show n in tickling,
where self-generated tickle is perceived by the subject as less ticklish (Blake-
more et al., 1998b); an attenuation, which is argued to be possible by the
cancellation of self-produced stimuli. The problems that occur w hen there
is a failure to monitor the consequences of action include the false attribu
tion of movement, and a false perception of the world. Grusser and Landis
(1991) report a patient, LM, with akinetopsia after bi-lateral lesions in the
occiptio-temporal region who fails to distinguish m ovem ent caused due to
her own action from movement in the outside world. According to their re
port "w hen walking across the garden or along the street, she reported that
the objects in her extra-personal space were moving up and dow n". This
m ay be caused by a difficulty in discriminating visual image m otion origi
nating from external sources and motion from the patient's ow n activity.
Differences in the response to external and self generated disturbances
are reflected neurophysiologically in the differing response that are m ade
betw een self and externally generated touch. Cells in the tem poral lobe,
move-Introduction 24
m ent generally, and respond to the type of m ovem ent rather than the form
of movement. In particular, they have been shown to respond selectively
to unexpected movements. If the monkey itself creates m ovem ent in the
w orld as a consequence of its own actions, the cells are silent. For example,
these cells have been found to respond to the sight of an object moved by
the experimenter but do not respond w hen the same object is moved by the
monkey (Hietanen and Perrett, 1993; Hietanen and Perrett, 1996b; Hietanen
and Perrett, 1996a). These cells are thought to contribute to the m onitoring
of the consequences of action (Mistlin and Perrett, 1990). In one experiment,
a monkey was trained to move a grating by rotating a striped drum . This
was compared w ith the response to a grating moved by an experimenter.
Therefore, the link between the m onkey's actions and the changes in the
world were correlated. After training the nervous system registered the
correlation between action and consequence, so that changes in the world
generated by the monkey no longer activated the STPa neurones.
Forward models are thought to be an im portant p art of systems that use
an efference copy of the motor comm and to anticipate and cancel the sen
sory effects of a given movement. Forward sensory m odels are responsible
for this prediction that is derived from the state prediction of a forward dy
namic model.
1.2.2 D istal supervised learning
A problem faced by the motor system is that of translating goals and errors
controller. Jordan and Rumelhart (1992) have show n how a forw ard model
can be used to estimate the motor errors during perform ance by the back-
propagation of sensory errors through the model. This is know n as distal
supervised learning as the supervision of the task, that is the detection of
errors is distal to the desired detection of m otor signal errors. A forward
model could be used to transform errors between the desired and actual
sensory outcome of a movement into the corresponding errors in the motor
comm and (Jordan, 1995).
1.2.3 M ental practice
Mental practice can lead to improved performance at a m otor task (Yaguez
et al., 1998), particularly in the early stages of learning a task (Bohan et al.,
1999). This improvement could be due both the m onitoring of performance
and m otor learning taking place in the absence of actual movement. A
forward model could be useful in mental practice, as it could be used to
predict the outcome of different imagined actions (Jordan and Rumelhart,
1992; Wada and Kawato, 1993; Miall and Wolpert, 1996). This w ould en
able the controller to select between the imagined actions, or allow the
controller to adapt. The notion that the underlying m echanisms for both
physical and imagined practice are similar is supported by im aging (De-
cety et al., 1994; Decety, 1996); the compliance of im agined m ovem ents to
FitTs Law (Decety et al., 1989; Decety and Jeannerod, 1996; Decety, 1996);
patient studies (Kagerer et al., 1998) and a similarity in the interference ef
Introduction 26
during m otor imagery have been suggested to be related to the functional
and neural changes that are observed in the consolidation of motor m em
ories (Shadmehr and Brashers-Krug, 1997; Shadm ehr and Holcomb, 1997).
This alteration in stability could be due to increased learning by motor im
agery (Bhushan and Shadmehr, 1999).
1.2.4 MOSAIC
The MOSAIC model (MOdular Selection A nd Identification for Control)
(Kawato and Wolpert, 1998) proposes the use of paired forward and in
verse m odels for control. The forward models compete to predict the sen
sory feedback given a m otor command, so that each becomes specialized for
a different dynamic situation, such as m anipulating different objects. The
prediction of each model can be compared to the actual feedback to provide
a set of prediction errors. The smaller the prediction error of a particular
model, the more likely that model is to model the current dynamics. The
set of forward models, therefore, act as a set of hypothesis testers and their
errors can be used to identify the current dynamics. The errors are then
used and to gate the contribution of a set of paired controllers to the final
motor command. Such m odularity has a num ber of benefits to the motor
system. Firstly, this architecture enables the retention of previously learned
behaviours to each situation. Secondly, a response to novel situations can
be derived from combinations of previously experienced context. Thirdly,
as the world is m odular and we act in a variety of different contexts, this
1.3 Learning and representation of an internal m odel
Both forw ard and inverse models simulate properties of the m otor system.
For such models to be functional, it is essential that they are not static, and
can be modified as new environments are encountered, for example novel
objects are m anipulated, and also to take into account changes that occur
in the properties of the arm due to growth and injury. The ability to leam
and use new internal models is, therefore, a fundam ental property of the
m otor system (Shadmehr and Brashers-Krug, 1997; Wolpert, 1997; Wolpert
and Ghahramani, 2000).
An example of this learning occurs if subject's are asked to move a 2.5 kg
weight, where after training the trajectory is the same as before the weight
w as added (Lacquaniti et al., 1982). Dynamic learning has been used to
study the development of internal models appropriate for novel force fields
(Shadmehr and Mussa-Ivaldi, 1994; Gandolfo et al., 1996). Such paradigm s
for studying the development of inverse models of the m otor system are
useful frameworks for the examination of learning of forw ard m odels of
m otor control; the focus of this thesis. Within the studies examining the
adaptation of inverse models, subject's were trained to m ake point-to-point
m ovem ents in a novel force field. Initially the force field caused a signif
icant divergence from the normally observed trajectory (Shadmehr and
Mussa-Ivaldi, 1994; Gandolfo et al., 1996). With practice in the novel en
vironment, the trajectory eventually converged to the trajectory before the
introduction of the field. This dynamic learning was assessed by the exam
Introduction 28
during one point-to-point movement. This enabled the aftereffects of the
exposure to the force field to be examined. It was found that the trajec
tories occurring during these "catch trials" were the mirror-images of the
observed trajectories w hen the subjects were initially exposed to the force
field, w ith the m agnitude of these after-effects gradually increasing over
the training period. The presence of such after-effects w hen no force field
was present shows that the CNS had constructed an internal model of the
force field which was generating patterns of force that anticipated the pre
viously encountered perturbing forces (Shadmehr and Mussa-Ivaldi, 1994).
The learning of such internal models has been show n to be initially fragile,
but becomes robust after a few hours (Brashers-Krug et al., 1996). W hen
subjects learned to make reaching movements under 2 conflicting m echan
ical environments it was found that two motor m aps could be learned and
retained. However, it was only w hen the training of the two tasks w as sep
arated by approximately 5 hours that performance on both tasks was not
impaired. If the training separation was less than this, the performance on
the first task was impaired; that is reteroactive interference occurs. Once
successfully learned, the memory was present over a 5 m onth period. This
suggests that the learning of the internal models had tw o components; ini
tially memory was fragile but after consolidation of this internal m odel with
practice, memory became robust.
Generalization paradigm s have been used to examine the representa
tion of internal models. The generalization paradigm can be sum m arized
re-gion of input space. Secondly, subjects are exposed to a novel input-output
rem apping over a limited region of input space. Finally, subjects are once
again tested on their input-output m apping on the full region tested in the
first phase. The pattern of generalization outside of the learned region re
flects the structure and constraints underlying the internal m odel (Ghahra
m ani and Wolpert, 1997; Ghahramani et al., 1996). Such paradigm s have
been used to examine the representation of inverse m odels (Shadmehr and
Mussa-Ivaldi, 1994; Gandolfo et al., 1996).
In this thesis we will examine the learning of forward models, including
the use of catch trials to assess the development of an appropriate predic
tion. In Chapter 7, a generalization paradigm will be used to examine the
spatial representation of a forward model.
1.4 Grip force as a paradigm for forward m odels
Grip-force m odulation provides an ideal paradigm to study internal model
learning, due to the differing latency of response that occurs to alterations
in load force that are self-generated compared to adjustm ents in load that
are externally generated. These differences are thought to be due to the in
volvem ent of a forward model of both the m otor system and the controlled
object.
W hen we hold an object in a precision grip betw een the thum b and fore
finger, sufficient grip force (perpendicular to the surfaces) m ust be gener
ated to prevent the object from slipping (Johansson and Westling, 1984; Jo
Introduction 30
Acceleration (a)
iGrip Force
Load Force (mg +ma)
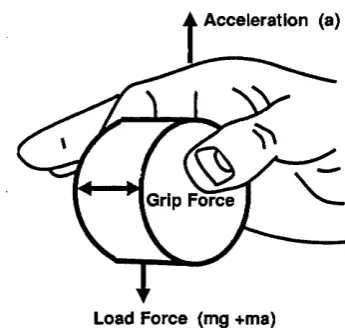
Figure 1.1 — Grip force is generated perpendicular to the object surfaces. The level of grip force required is dependent on the load force that the object exerts on the fingers.
Johansson, 1996). The level of grip force required depends on the load force
(tangential to the surfaces) the object exerts on the fingers, that is its weight
w hen at rest, and the frictional properties of the surface of the object. Fig
ure 1.1.
W hen the object is moved by the subject the load force on the fingers
m ust change to accelerate the gripped object. W ithout a corresponding
change in grip force the object w ould slip. In this self-generated condition
the grip force tends to parallel load force with negligible delay.
Such anticipatory m odulation is seen in discrete self-generated move
ments w hen pulling on an object in different directions and at different
speeds (Johansson and Westling, 1984; Johansson et al., 1992). Anticipa
tion has also been shown to occur during continuous self-generated move
m ents (Flanagan and Wing, 1993; Flanagan and Wing, 1995). W hen an ob
ject is m oved by the arm, grip force modulates in phase w ith load force.
Grip force increases as the load increases, and also falls as the load de
Grip force m odulation has been examined in other grips to assess the
specificity of these parallel m odulation between grip and load force to the
precision grip (Flanagan and Tresilian, 1994). Norm al, inverted and bi
m anual grips were examined. In the inverted grip, or "Pirg", outw ard
rather than inw ard force was required to hold the object. Additionally the
direction was varied to produce different patterns of load force changes
during the movement. It was found that regardless of the grip used, m od
ulation of grip force occurred in parallel with alterations in load force. In
creased variability that was present in the pirgs w as explained by the re
duced sensory feedback from cutaneous afferents available in these grips.
This coupling of grip and load has also been show n in m ultidigit grips (Burst-
edt et al., 1999).
Anticipation was found not to be specific to increases in load force caused
by arm movements and was present w hen subjects were instructed to jum p
so that the object was actively m oved by the subject w ithout arm move
m ent (Flanagan and Tresilian, 1994). Similar parallel m odulation of grip
and load have been found w hen the movement on the object is caused by
w alking and running (Kinoshita et al., 1993).
This invariance of the grip-load force coupling across grip types and
m odes of transport suggest a general strategy for coordinating grip and
load forces during active transport of a held object. The forw ard model
incorporates movements of the arm and body. Grip is m odulated w hen the
net result of all movements leads to object acceleration and therefore fluctu
Introduction 32
Motor C om m and
Predicted Hand
Internal Acceleration Internal
Model of --- ► Model of
Arm Object
Anticipatory Grip Force
Efference Copy
Figure 1.2 — Internal model for predictive grip force modulation
Despite the relatively rapid response of cutaneous afferents, such an
ticipatory control cannot be explained as a reaction to peripheral feedback
(Johansson and Westling, 1984; Flanagan and Wing, 1995) due to unavoid
able feedback delays (Johansson and Westling, 1984; Forssberg et al., 1992).
A system based solely on feedback control w ould be ineffective for m anip
ulative actions w ith frequencies above about 1 Hz, which w ould exclude
some complex skills (Kunesch et al., 1989; Johansson and Cole, 1994).
This suggests that the skilled m anipulation of objects requires the cen
tral nervous system (CNS) to use the motor command, in conjunction with
internal models of both the body and the object, to anticipate the resulting
load force and thereby adjust grip force appropriately (Flanagan and Wing,
1997b; Wing et al., 1997; Blakemore et al., 1998a), Figure 1.2.
1.4.1 Reactive grip force m odulation
The predictive response present in response to self-generated loads can be
contrasted w ith the response when the load force is unexpectedly increased,
for example by someone else tapping on the object. In this externally-generated
fingertips w ith a latency of around 60-100 ms after load force (Johansson
and Westling, 1988; Cole and Abbs, 1988; Johansson et al., 1992; Blakemore
et al., 1998a).
This reactive grip force response m ust depend on the feedback from cu
taneous afferents as afferents from intrinsic and extrinsic han d muscles and
the interphalangeal joints do not respond to alterations in load force fast
enough (Macefield and Johansson, 1996).
The automatic nature of adjustments m ade to slip, and the reliance of
feedback from cutaneous afferents to signal alterations in object properties
have been examined in three ways (Westling and Johansson, 1987; Johans
son and Westling, 1990). Firstly, alterations in force coordination that usu
ally w ould follow object slip could also be elicited by afferent volleys in tac
tile units (Westling and Johansson, 1987). W hen the tactile afferents inner
vating the skin areas in contact with the object were electrically stimulated,
a m otor response occurred that was similar to that following a slip.
Secondly, latency data shows that the m otor responses to natural slips, as
well as the similar responses to weak cutaneous electrical stim ulation were
automatic in nature and not voluntarily initiated. The latency betw een a
slip and the onset of the force ratio change w as twice the latency of the
most rapid spinal reflex in intrinsic hand muscles at approximately, 75 ms,
but is less than half the voluntary reaction time. This latency is associated
with long-latency reflex responses to suddenly im posed finger or thum b
movements (Johansson and Westling, 1987).
Introduction 34
is illustrated by the observed 'm agnet phenom enon' observed w hen sub
ject's attem pt to slowly separate their fingers while holding the object in
the air. The regulation of the g rip /lo ad ratio by signals from cutaneous
afferents mechanisms appears to interfere w ith the voluntary drive, and
subject's are reported to experience the sensation of the object sticking to
their fingers. The effort required to overcome this 'm agnet phenom enon'
becomes stronger the closer the grip force is to the slip force, ie high fric
tional surfaces (Johansson and Westling, 1987).
1.4.2 Development of grip force response
Object properties are learnt through development, indicated by increas
ing ability to correctly parameterize grip force to the object being m anip
ulated (Forssberg et al., 1991; Forssberg et al., 1992; Eliasson et al., 1995;
Forssberg et al., 1995).
In contrast to the highly program m ed and invariant grip and load forces
generated by adults in separate lifts with objects, children exhibit substan
tial variability in both the force am plitudes generated and the tem poral
response (Newell and McDonald, 1997). This variability of m otor output
in children is not restricted to the control of grip force and is present in
a num ber of other tasks including postural control, reaching and locomo
tion (Manoel and Connolly, 1997; Horak, 1996; Konczak et al., 1995; Ashby
et al., 1997; Schmitz et al., 1999). It has been suggested that this increased
variability in force production, which w ould norm ally be viewed as a mal
Schmitz et al., 1999). This variability may allow the CNS to evaluate dif
ferent response patterns and relate the intended m ovem ent to the motor
consequences (Newell and McDonald, 1997). Therefore variability w ould
facilitate the establishment of relations between the m ovem ent param eters
and interactions within the environment. Additionally such variability may
also reflect constant adaptation to a system that is rapidly changing due to
changes in limb length and mass during growth.
Grip force m odulation in children is less precisely tim ed than in adults,
w ith the tight correlation between grip and load force increases absent dur
ing loading. Instead, the strategy used is to initiate a grip force increase
before the increase in loading due to object movement. Therefore, once the
object has been accelerated, the children were already gripping the objects
substantially, w ith any further increases grip force not in parallel w ith the
load force. In experiments where both adults and children were required to
perform a task where load force ram ped up, and then rem ained constant,
show ed that in children the movement phases of the lift are m uch longer
than in adults (Gordon and Forssberg, 1997). This increase is thought to be
partly due to prolonged delays from the mechanical events signaling the
term ination of one phase and the onset of the motor activity beginning the
next phase.
By the end of 2 years, children's grip and load forces begin to increase
more in parallel, with this coupling increasing to adult values by 8 years,
through improvements can occur as late as 14 years (Forssberg et al., 1991;
Introduction 36
1.4.3 Learning the appropriate grip force response
Grip force levels can be set w ithout somatosensory feedback, anticipating
the physical properties of the object, that include the object's weight, shape
and friction at its surface (Johansson and Westling, 1984; Johansson and
Westling, 1988; Johansson and Cole, 1994; Jenmalm and Johansson, 1997).
Gordon et al.(1993) examined the grip force scaling to commonly m anip
ulated objects, including crispbread packages and tins. It was found that
w hen lifting these 'common objects', grip force is program m ed prior to ini
tiation the lift to match the expected weight of the object based on previous
experience. Test objects were then presented to subjects; objects w ith a mass
that had not previously been experienced. W hen subject's lifted these test
objects, grip force became correctly scaled to load force w ithin the first few
trials. Once learned, the subject was able to correctly scale their grip force
appropriate for this test object after a delay of 24 hours. However, if a novel
object, that is an object w ith the visual appearance of a "common object"
but differing in density, was presented to the subject, the default grip force
based on the visual appearance of the object was used, w ith up to ten trials
needed to correctly adjust the grip force.
Flanagan and Wing (1997) show ed that the anticipatory increase in grip
force that occurs w hen a subject accelerates an object is adaptable to novel
force environments. W hen the subject was exposed to inertial, viscous or
composite force fields during point-to-point m ovem ents it w as found that
the level of grip force quickly updated to m odulate in parallel w ith the new
Wing and Flanagan, 1998).
Similar rapid adaptations to novel environments have been show n w hen
the gravitational force has been altered (Hermsdorfer et al., 2000). Load
force changes on an object occur during parabolic flights, where gravity
ranges from hypergravity (2g) and a short period of m icrogravity (Og). Sub
ject's m anipulating an object under different gravitational loads quickly
leam to m odulate grip as the novel load force changes. The predictive cou
pling betw een grip and load force occurred in transitions betw een different
gravity levels, illustrating the rapid adaptation to changing load conditions.
These findings are strong evidence for the existence of a forw ard internal
model that is predicting the properties of both the m otor system and the ob
ject being manipulated. If subject's were unable to adapt to the new loads,
it w ould be suggestive of a simple association between the m otor command
and grip force that leads to appropriate m odulation of grip force during ob
ject acceleration. However, this is clearly not the case, and the m otor system
is able to compensate for changes in the environmental context, resulting in
appropriate predictions being generated.
The importance of previous m anipulative experience in correctly scaling
grip force is illustrated by the force development pattern that can be seen
in children where m ulti-peaked force rate profiles can be seen (Forssberg
et al., 1991). This strategy is similar to the probing strategy used by adults
w hen lifting a novel object, or w hen they are uncertain as to the physical
properties of the object. The dependence of anticipatory control is reduced,
Introduction 38
breakage is avoided.
1.4.4 Bio-mechanics of grip force
The precision grip is an example of the redundancy of the muscular sys
tem, and requires the activity of both intrinsic and extrinsic hand m us
cles (Smith, 1981; Muir, 1985; Maier et al., 1993). At least 15 muscles are
thought to have a contribution in the exertion of force w hen holding an ob
ject with a precision grip (Napier, 1956; Napier, 1962). The grip requires
stabilization of the three joints of each finger, which can be considered as
a bio-mechanical bar linkage system (Hepp-Raymond et al., 1996). It also
requires that the compression force, that is the static equilibrium, as well
as the arch-like posture is m aintained between the thum b and index finger.
According to biomechanical constraints the extrinsic muscles w ith tendons
spanning all four links are best suited for the provision of a continual force
output, whereas the intrinsic muscles can adjust and m odulate grip force.
Long and Brown (1964) showed that the extrinsic muscles provide the m ain
compression force, assisted by intrinsic muscles that include the first dorsal
and palm ar interossei (IDI and 1 PI). Of the thenar muscles, the flexor polli-
cis brevis (FPB) exerts compresion force by the m etacarpophalangeal flexion
of the thum b w ith the adductor pollicis (ADP) providing compression force.
1.4.5 C utaneous afferents
In a precision grip task, cutaneous input is crucial for the updating of grip
Westling, 1987; Westling and Johansson, 1987). If an object is slippery, for
example silk material rather than sandpaper, the ratio betw een the grip
force and load force has to be higher. W hen the surface m aterial changes
on a trial by trial basis, the motor output is influenced by the new frictional
conditions at different phases of the lift (Johansson and Westling, 1987).
The glabrous skin of the hand, especially the fingertips are densely in
nervated by mechanoreceptors (Johansson and Vallbo, 1979; Johansson and
Vallbo, 1983; Lofvenberg and Johansson, 1984). Early studies show ed the
basic differences in tuning between mechanoreceptors in monkeys and com
pared these characteristics to hum an psychophysical data (Mountcastle et al.,
1969). Four types of mechanoreceptors have since been characterized (FAI,
FAII, SAI, SAII) (Knibestol and Vallbo, 1970). Mechanoreceptors are subdi
vided into two categories dependent on their response to a sustained step
indentation of the skin. 44% of the units are slowly adapting, that is they re
spond with a sustained discharge, and 56% of the units are fast adapting, re
sponding w ith a burst of impulses at the onset and removal of the stimulus.
Within these two broad subdivisions, two different units are distinguish
able on their receptive fields. Fast adapting Type I (FA I) and Slow adapting
Type 1 (SA I) have small and discrete receptive fields. This is in comparison
w ith the FA II and SA II units, which have wider receptive fields. The den
sity of glabrous skin receptors is highest in the fingertip, where the density
of FA I units and SA I units is about 140 units/cm ^ and 70 units/cm ^. At
a given point, the m ean overlap of the receptive fields of the FA I units is
Introduction 40
for the SAI units the overlaps are 16,6, and 2 respectively (Lofvenberg and
Johansson, 1984; Johansson and Vallbo, 1983).
The different mechanoreceptors also differ in their potential functions
for the characterization of object properties.
The contribution of each type of mechanoreceptor has been examined
at different stages of lifting, holding, releasing objects, and during abrupt
changes in tangential forces (Cole and Johansson, 1993; Johansson and West
ling, 1987; Johansson and Westling, 1984; Macefield et al., 1996). FA 1 units
are found to respond optimally to small force changes related to initial con
tact, slip and release of the object and are sensitive to the location and edge
contours of objects in contact with the skin. FAll units are highly responsive
to mechanical transients, so are especially active during the initial acceler
ation and deceleration of the object. SAI units are found to be sensitive to
changes in grip force, and SAll units are particularly responsive to changes
in tangential load force. The coupling of grip and load forces in m anipula
tion forms the basis for an efficient sensory control of the m otor output to
accommodate the physical properties of the object. Such synergies represent
strategies to simplify the dem ands on the control m echanism by reducing
the num ber of degrees of freedom of the musculoskeletal system that have
to be explicitly controlled (Bernstein, 1967).
1.4.6 Slip
Once the object is touched the most relevant afferent inform ation for the fi
the object is provided by the mechanoreceptors located in the glabrous skin
of the digits (Lofvenberg and Johansson, 1984). The importance of input
from cutaneous afferents can be illustrated in people w ith im paired sensi
bility in the fingers. Damage to the cutaneous afferents lead to difficulties in
holding and gripping objects, even with visual compensation. This crucial
role was dem onstrated by Mott and Sherrington (1895) who show ed that
w ith spared distal cutaneous innervation, little functional im pairm ent was
evident in the forelimb even if the rest of the limb was deafferented.
In general, subjects avoid excessive grip forces; applying enough to pre
vent the object slipping w ith grip force rarely exceeding the m inim um grip
force by more than 30% safety margin (Johansson and Cole, 1992). This
strategy avoids both object breakage and unnecessary muscle fatigue. There
fore, as the load force increases, the grip force m ust also increase to prevent
slippage and to maintain this safety margin.
1.4.7 Central control
The precision grip, with opposition of the thum b and index finger is charac
teristic of m anual skill and dexterity (Wiesendanger, 1999).This grip allows
the skilful m anipulation of objects coupled w ith the precise control of pre
hensile force (Napier, 1956; Napier, 1962). Based on anatomical, physiolog
ical and behavioural studies, the motor cortex and cortico-motomeuronal
pathw ays are essential for such precision grip and object m anipulation (Lemon,
1993; Lemon et al., 1995; Lemon et al., 1996).
fin-Introduction 42
ger m ovem ents was first demonstrated by Lawrence and Kuypers (1968)
who show ed that when the tract was lesioned in the m onkey the ability to
make individual finger movements was lost. Across species comparisons
highlight how the development of the corticospinal tract is mirrored by an
increasing ability to perform individual finger movements, culminating in
the ability to perform a precision grip (Lemon, 1993). A lthough the hands
of m ost prim ates are physically similar they substantially differ in the de
gree of dexterity. N apier (1961) classified primates by an index of dexterity
for prim ates that reflect these substantial differences in grasping. For ex
ample, tree shrews w ith a dexterity index of 3 have whole arm control w ith
a basic grasping action. These animals are lacking cortico-motoneuronal
projections. The lemur, with an index of 5 is skilled at prehension but not
at object m anipulation. In contrast, the old world monkeys w ith an index
of 6 can perform pow er and precision grips. These prim ates have substan
tial cortico-motoneuronal projections. Heffner and M asterton (1975,1983)
show ed that there were two factors that were highly correlated w ith m an
ual dexterity; firstly, the level that the corticospinal tract penetrated, and
secondly the presence of cortico-motoneuronal projections that reach into
the motoneuronal cell groups of lamina IX i.e. the deepest spinal lamina.
The developm ent of the precision grip in hum ans parallels the development
and m aturation of cortico-motoneuronal connections. Initially infants grasp
is m ainly caused by tactile and proprioceptive reflexes (Eliasson et al., 1995).
Gradually, these reflexive grasping patterns become more stereotyped as
children w ith Cerebral Palsy, who fail to develop the corticomotomeuronal
p ath have impaired m anual dexterity and tend to exhibit m anipulative pat
terns similar to those in young infants (Eliasson et al., 1991; Eliasson et al.,
1995).
1.4.8 Cortical areas
Four m ain cerebral areas are thought to have an im portant role in the control
of hand movements; prim ary motor cortex (Ml); the supplem entary motor
area (SMA); prem otor cortex (PM) and cingulate m otor areas (CM) (Ehrs-
son et al., 2000; Kinoshita et al., 2000). The output from low-threshold
mechanoreceptor afferents is transmitted to the cerebral cortex where they
contribute to the updating of the grip load force ratios (Johansson and Cole,
1992) w ith the somatosensory cortex sending direct projections to the motor
cortex (Jones and Powell, 1970). Reversible inactivations of the somatosen
sory cortex by injection w ith muscimol was found to disrupt fine finger
movements, w ithout any disturbance of reaching and hand shaping (Hikosaka
et al., 1985). The participation of the m otor cortical neurons w ith cutaneous
receptive fields in adapting grasping and lifting forces to different surface
textures and weights has been dem onstrated (Picard and Smith, 1992a; Pi
card and Smith, 1992b).
The primate motor cortex, known to play a key role in the control of
fine finger movements, receives detailed information over very rapid path
ways from the mechanoreceptive units of the glabrous skin. Lemon (1981)
Introduction 44
a m otor cortex neurone during grasping. In this study m ost of the neurones
w ith cutaneous inputs were more highly activated during a task which in
volved the use of a precision grip to reach rewards than during a lever-
pulling task.
A large area of M l is involved in the selection of distal m otor muscles of
the contralateral forelimb that can evoke movements of the fingers or wrist.
Each muscle and type of movement has been found to be m ultiply repre
sented. Therefore, M l is thought to be primarily involved in the selection
of the appropriate muscles to perform the movement, w ith the other corti
cal areas influencing output from motor cortex indirectly, especially by the
dense projections from these areas to the hand representation of M l (Porter,
1972). SMA is thought to be particularly relevant to the control of bim anual
tasks (Wiesendanger, 1986; Viallet et al., 1992)
1.4.9 Anticipatory grip force
Muller and Dichgans (1994a,1994b) reported patients w ith degenerative cere
bellar lesions who they found show ed a lack of co-ordination of grip and
load forces w hen performing lifting tasks using a precision grip. Grip and
load force had become decoupled in these patients, so that the forces did
not consistently change in parallel as has been observed in norm al patients
could adjust grip force rates, their ability to do so was significantly lower
than controls. Particular problems were observed in adjusting grip force to
the differing load forces associated w ith objects of differing weight. They
ad-just grip force in parallel w ith load force was due to a failure of anticipa
tory parameterization. Patients w ith unilateral cerebellar dam age showed
a selective impairm ent in ability to m odulate grip force on the affected
side (Muller and Dichgans, 1994b). Further cerebellar patient studies have
examine w hether adjustments in grip force occur to fluctuations in iner
tial load force during arm movements (Babin-Ratte et al., 1999). This study
show ed that unlike normal subjects who adapt to fluctuations in load force,
a patient with cerebellar degeneration was unable to accurately m odulate
grip force with load force. Grip force was found to be increased in m ag
nitude, and there were no timing differences between upw ards and dow n
w ards movements. The total load force is the sum of inertial load forces
and gravitational load force. Therefore, the point of m axim um loading dif
fers betw een upw ard dow nw ards movements and cerebellar patients were
not appropriately scaling their grip force.
Evidence for the involvement of the cerebellum in grip force m odulation
also comes from electrophysiology; neurones in the cerebellum as well as
prim ary m otor cortex have been reported to fire related to object weight and
texture prior to the onset of movement (Espinosa and Smith, 1990; Picard
and Smith, 1992a).
1.4.10 Earlier theories of predictive grip force modulation
The presence of forward models of the motor system to account for the
predictive m odulation of grip force to self-generated loads helps to qual