GENERAL SELECTION FOR SPECIFIC DNA-BINDING
ACTIVITIES
NICHOLAS BENSON, PAUL SUGIONO, STEVEN BASS, LYNN V. MENDELMAN A N D PHILIP YOUDERIAN Department of Biologzcal Sciences, University of Southern Calijornia,
Los Angeles, Calzfornia 9 0 0 8 9 - 1 4 8 1
Manuscript received October 1, 1985 Revised copy accepted May 7, 1986
ABSTRACT
We present a general strategy for the selection of bacterial clones that express DNA-binding activities corresponding to particular DNA recognition sites. T h e selection uses a “challenge phage” vector, P22 K n 9 a r c - a m H 1 6 0 5 , into which is substituted a synthetic DNA-binding site for a site that controls transcription of the P22 antirepressor ( a n t ) gene. Constitutive synthesis of antirepressor channels a challenge phage into lytic development and efficiently kills a n infected host, unless the substituted site is bound by a specific protein; in this case, the chal- lenge phage prefers lysogenic development, and the host survives and acquires an antibiotic-resistance phenotype. Infections with challenge phages carrying the E. coli Lac operator, phage XOL, operator, or synthetic, “idealized” E. coli Trp and T n l O T e t operators select clones that express each of the corresponding binding activities. T h e use of challenge phage vectors may be extended to select clones that express eukaryotic DNA-binding activities.
ENE expression is often regulated by transcription factors that bind to
G
specific, short sequences of DNA. These cis-acting regulatory sites are normally situated close to a regulated structural gene; often, they overlap the promoter (RNA polymerase binding site) for the structural gene. Presumably, many eukaryotic transcription factors act as their prokaryotic counterparts, repressors and activators, by binding specific sites to inhibit or stimulate the initiation of transcription.Motivated by the idea that some eukaryotic transcription factors, when ex- pressed in prokaryotic cells, will retain their DNA-binding activities, we have developed a selection for isolating clones that express binding proteins corre- sponding to specific DNA sites. I t is our hope that, by extending this selection
to eukaryotic regulatory sites, we shall be able to select clones that express corresponding transcription factors from plasmid cDNA libraries.
Our selection involves a derivative of temperate Salmonella phage P22 called a “challenge phage.” Challenge phages exploit the unique genetic properties of the P22 imml region (Figure 1). T h e primary feature of the imml region is the ant gene. T h e product of ant, antirepressor, can inactivate the primary prophage repressor, c 2 protein (LEVINE et al. 1975; BOTSTEIN et al. 1975;
2 N. RENSON E T A L .
a.
b.
-35
Pant arc
*
mnf-
Pmnr-10 +I +34
c.
~ ~ G A T A G A A G C A C T C T A C ~ V ~ ~ T ~ ~ T C A A T [ P G G T C C A C G G T G G A C C $ T A T T G T G A G G T G A A ~Panr Omnr arc
FIGURE 1 .-The P22 imml region. a , Organization of the P22 prophage genome, showing the pattern of early transcription (arrows). T h e imml region (thick line) is embedded within the late operon, in the right prophage arm. b, Transcriptional organization of the imml region. T h e a n t promoter and Mnt operator are represented by a thick line; genes are indicated by open boxes.
c , Partial sequence of the sense D N A strand of the a n t promoter and Mnt operator, showing the position of the operator with respect to the most highly conserved promoter sequences and the initiation codon of the arc gene (boxed).
SUSSKIND and B O T s T E I N 1975). P22 c 2 repressor, like X cI repressor, is directly
responsible for preventing the transcription of the phage early, vegetative func- tions (BRONSON and LEVINE 1971; POTEETE et al. 1980; POTEETE and PTASHNE
1982). Constitutive synthesis of antirepressor prevents the action of c2 repres- sor and, thus, prevents lysogeny.
P22 must produce t w o repressors to maintain the lysogenic state: c 2 repres- sor, to prevent the synthesis of early vegetative functions, and Mnt (maige- nance) repressor, to prevent the synthesis of antirepressor. Mnt protein acts by binding a single operator site located at the startpoint of transcription of the ant promoter I O prevent the binding of RNA polymerase (SAUER et al.
1983; \ r et al. ~ 1985a; D. ~ HAWLEY ~ and ~ W. R. MCCLURE, ~ ~ unpublished results).
Synthesis of antirepressor is also repressed during lytic development by the product of the first gene in the ant operon, arc (for antirepressor control) (SUSSKIND 1980). Upon infection of a sensitive host by P22 arc- phage, the state of occupancy of the M n t operator is a critical determinant of the devel- opmental course of the infection. I f the Mnt operator is free of Mnt repressor, the ant gene is expressed at high constitutive levels, antirepressor inactivates c 2 repressor, and the infection proceeds lytically. If, on the other hand, active
M n t repressor preexists in cells infected by P22 arc- phage, Mnt binds its operator, ant is not transcribed and antirepressor does not interfere with the establishment of lysogen),.
of the substituted site governs this decision for a challenge phage. Challenge phages carry an additional mutation, Kn9, the substitution of an antibiotic- resistance determinant for the mnt repressor gene, so that lysogens surviving a challenge infection may be recognized by a selectable phenotype. T h e chal- lenge phage arc gene is conditionally inactivated by an amber mutation to facilitate growth under permissive (suppressed) conditions (SUSSKIND 1980). These three mutations, arc-amber, an operator substitution and Kn9, conspire to enable a challenge phage to selectively lysogenize, and therefore spare from death, a host cell that produces a specific DNA-binding activity corresponding to its substituted site.
In previous reports, we demonstrated that infection with challenge phages carrying symmetric, mutant Mnt operators could be used to select mutant Mnt repressors with altered specificities of DNA binding (YOUDERIAN et al. 1983; VERSHON et al. 1985b). In this report, we extend this selection to show that challenge phages with any of several different binding sites of prokaryotic origin can be used to select clones that produce specific, corresponding DNA- binding activities.
MATERIALS AND METHODS
Bacterial and phage strains: Bacterial strains are derivatives of S . typhimurium L T 2
or E. coli K12. Salmonella strains DB7000 (leuA-am414 sup"; SUSSKIND, WRIGHT and BOTSTEIN 1971) and its supE derivative MS1363 (SUSSKIND 1980), used for the permis- sive growth of P22 arc-am phages, have been described. Otherwise isogenic hosts, MS1868 (sup") and MS1883 (supE), also carry a mutation ( r - ) that inactivates one of several Salmonella restriction systems preventing efficient transformation ( G R A ~ ~ A , YOUDERIAN and SUSSKIND 1985). MS1367 is a derivative of MS1363 that carries a P22
c2+ mnt- prophage deleted for the imml region (SUSSKIND 1980). MS1582 is an immune (c2+ m d ) lysogen of prophage P22 16-amH1455 sieA44 Ap68tpfr49 in MS1363, used for the isolation of virulent mutants of P22 ( G R A ~ ~ A , YOUDERIAN and SUSSKIND 1985). Plasmids were constructed in E. coli strains C600 (APPLEYARD 1954) o r MM294 (ME-
SELSON and YUAN 1968) before transformation into Salmonella (LEDERBERG and COHEN 1974). F'lac episomes were introduced into MS1868 by conju al transfer from E. coli E5014 (F'lac+; SIGNER and WEIL 1968) and E. coli X90/F'lacl~1 (AMANN, BROSIUS and PTASHNE 1983).
Media, enzymes, and chemicals: Media and general phage techniques have been
described (LEVINE 1957; LEVINE and CURTIS 1961; SIGNER and WEIL 1968; SUSSKIND, WRIGHT and BOTSTEIN 1971; BOTSTEIN, CHAN and WADDELL 1972; WEINSTOCK, SUSSKIND and BOTSTEIN 1979; YOUDERIAN et al. 1983). Antibiotics were obtained from Sigma Chemical Company. Restriction endonucleases, E. coli DNA polymerase I large fragment, T 4 polynucleotide kinase and T 4 DNA ligase were purchased from New England Biolabs. Deoxyribonucleoside triphosphates and A T P were purchased from PL Biochemicals. C Y - ~ ~ P - ~ A T P (700 Ci/mmol), used in DNA sequence analyses, was from ICN Biomedicals, Inc. Synthetic XbaI (5' CTCTAGAG 3') and XhoI (5' CCTCGAGG 3') linkers were purchased from New England Biolabs. Synthetic Lac operator DNA (see Figure 3) was a generous gift from PONZY Lu. Synthetic XO,, DNA was a generous gift from ROBERT SAUER. Synthetic consensus, symmetric T r p and T e t operator DNAs were synthesized on a Systec Microsyn P1450A Automated DNA Synthesizer, using phosphoramidite substrates from ABN and Baker HPLC grade solvents (CARUTHERS
1982).
Plasmids: General procedures used in the construction, selection, and purification of
4 N. BENSON E T A L .
p PY 230 pMSlOO
ant‘
At1
I
Hlnf It x b o 1 linkers
1
0
FIGURE 2.-Construction of pPY 112. P22 DNA is indicated by thin lines; pBK322 DNA by wavy lines. P22 genes, or portions of genes (’) are indicated by thick lines. P , PstI; R, EcoKI; F,
H i n f f ; X , XbaI; 0, HpaI/PvuII junction. Only the relevant Hinfl site is indicated.
are derivatives o f pBR322 (BOLIVAR et al. 1977). pPY23O is a subclone of t h e Sau3AI a n t promoter fragment from P22 sieA44 mnt-tsl P,,,R1204 arc-amH1605 in t h e unique
BamH 1 restriction site o f plasmid plcIS99, oriented clockwise (YOUDERIAN, BOUVIER a n d
SUSSKIND 1982). T h e insert, flanked by EcoRI sites, was inverted by cleavage a n d
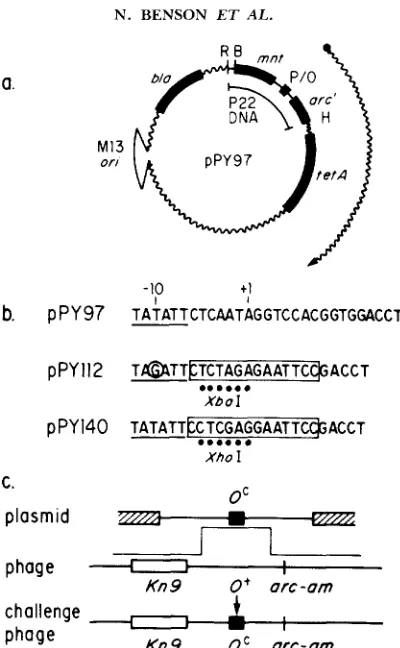
religation t o give pPY230. pMSlOO carries a n AvaII-HpaI fragment of P22 D N A sub- cloned into t h e EcoRI-PvuII “backbone” of plasmid pBR322 oriented in a clockwise direction (VERSHON et al. 1985a). pPY97 carries t h e EcoRII-Hind111 fragment o f P22 D N A carrying mnt, P,,, a n d t h e proximal two-thirds o f t h e a r c gene inserted in t h e EcoRI-Hind111 “backbone” of pZ150 (ZAGURSKY a n d BERMAN 1984), a derivative of pBR322 carrying t h e phage M 13 origin of replication a n d packaging initiation sites.
5
type arc gene. These two fragments were purified by electrophoresis through and elution from a polyacrylamide gel, then mixed and ligated to give pPY 1 12. Derivatives of pPY112 carrying XOLl and Oloc were transformed into MS1363 and crossed with phage P22 sieA44 Pa,,RE56 arc-amH1605 to separate the Pa,, mutation and the operator substitution (YOUDERIAN, BOUVIER and SUSSKIND 1982). Progeny phage having the genotype P22 sieA44 P,',, 0' arc-am were selected as virulent recombinants on immune host MS1582. Recombinant phages were used to transduce plasmid pMS97 from per- missive host MS 1 363 to nonpermissive host MS 1868. Tetracycline-resistant transduc- tants were found to carry mutants of P,'n, plasmid pMS97 with the OLI and Olac substi- tutions (pPY 149 and pPY240, respectively). Plasmid pPY140 is otherwise isogenic with p P Y l l 2 , except that it carries an Xhol linker, in place of an XbaI linker, and the wild- type allele of P0,,RI2O4. pPY300 is a subclone of the T r p operator in pPY140. pPIlOOT is a subclone of the synthetic Tet operator in pPI39 (P. J. ISACKSON and K. P. BER- TRAND, unpublished results), a deletion derivative of pPY 140 lacking both the pBR322 DNA between the unique Hind111 and Sal1 sites and the M13 origin fragment. (The pBR322 deletion removes a copy of the pBR322 T e t operator.)
pMC7 (CALOS 1978) carries a HincII partial fragment with the lac14 gene inserted in the unique EcoRI site of tetracycline-resistant plasmid pMB9 (RODRIGUEZ et al. 1976). T h e plasmid source of X repressor, pLRB, carries an operon fusion of the P l , , W 5
promoter to both the c l gene and the pBR322 tetA gene (NELSON and SAUER 1985). T h e source of TrpR protein, pPY150, was constructed by digestion of plasmids pRPG47 (ARVIDSON, CAN and GUNSALUS 1985) and pZ152 (ZAGURSKY and BERMAN
1984) with EcoRI and PvuII and by ligation of the products. It carries the M I 3 origin from pZ152 and a fusion of the Pl,,W5 promoter to the trpR structural gene from pRPG47. T h e source of TetR protein is plasmid pBI501 (ISACKSON and BERTRAND
1985), a subclone of a HinclI fragment containing the TnlO tetR structural gene fused to the P1.,W5 promoter in vector pUC8 (VIEIRA and MESSING 1982).
DNA sequence analyses: Challenge phage DNA, single-stranded plasmid DNA or
double-stranded plasmid DNA was sequenced by the method of SANGER, NICKLEN and COULSON (1977), using a synthetic oligonucleotide complementary to the 5' end of the arc gene as primer (5' CGGCATTTTGCTCATTCC 3'). Single-stranded plasmid DNA was prepared by the method of ZAGURSKY and BERMAN (1 984).
Construction of challenge phages: Challenge phages were constructed by crosses
between plasmids and phages (YOUDERIAN et al. 1983). T h e operator sequences of challenge phages were determined either directly from phage DNAs or from the plas- mids used in the construction crosses.
RESULTS
Construction of challenge phages: Challenge phages carrying substituted binding sites w e r e constructed in two steps. First, sites w e r e synthesized and
cloned i n t o t h e unique restriction sites of plasmids pPY 1 12 or pPY 140, deriv- atives of plasmid p B R 3 2 2 carrying t h e P22 antirepressor control region. Sec- o n d , sites were exchanged for t h e M n t operator o n P 2 2 K n 9 arc-am phage by
homologous recombination between the plasmid a n d phage genomes.
As shown in Figure 3, p P Y l l 2 a n d pPY140 carry a subcloned region of
P 2 2 D N A that includes t h e mnt gene, t h e ant p r o m o t e r a n d a portion of t h e
6
a.
N. RENSON E T A L .
- I ? +I
b.
p PY 97 TATATTCTCAAT~GGTCCACGGTGGACCT~ P Y 112
T~TTLTCTAGAGAATTCCACCT
...
XboI
p PY 140 TATATT~CTCGAGGAATTCC@ACCT
...
C.
OC
plasmid ZZl--
2
L
I
Kn9 0’ afc-am
Phage
-
challenge
4
phage Kn9 0‘ afc-om
FIGURE 3.-Challenge phage construction. a , Structure of plasmid pPY97, a subclone of the wild-type mnt gene, ant promoter and Mnt operator in plasmid pZ150. Transcription of the tetA
gene, indicated by the arrow, originates from the ant promoter within the EcoRII (B) t o Hind111 (H) fragment of P22 DNA. b, Sequences of the wild-type promoter/operator region in pPY97, and the substitutions (boxed) within this region carried by otherwise isogenic plasmids pPY 1 12 and pPY140. Unique restriction sites within the subsitutions are designated; note that they are flanked on the right by EcoRI sites (5’ G A A T T C 3’) that are not present in pPY97. T h e PantK1204
mutation (-10, T:A + G:C) is circled. c, Diagram of the cross between plasmids and P22 Kn9 arc-am phage to construct challenge phage [P22 DNA (thin line); T n 9 0 3 (Kn9) DNA (open box); pBR322 DNA (hatched box); operator substitution (filled box)].
stranded oligonucleotides representing the new binding sites. T h e circular products of these reactions include plasmids with a new site positioned so that its first base pair aligns with the first base pair of the Mnt operator in the wild-type sequence, the startpoint of transcription of the ant promoter.
As illustrated in Figure 4, we have subcloned oligonucleotides corresponding to the natural binding sites for E. coli Lac repressor and phage h cI repressor, and synthetic, “idealized” binding sites for E. coli Trp and transposon T n l O Tet repressors into the unique restriction sites of these plasmids. These recom- binant plasmids were transformed into Salmonella host MSl883 and were crossed with P22 KnY arc-am phage. Recombinant challenge phages that had
CHALLENGE PHAGE SELECTION
7
a
OIOC
X0,l
’rrp”
“fer“
b.
7f.p”
f f..
0roH
fmQ
c.
7 G f #I
-1p
*1
tl?. t2p”CTAG AATTGTCAGCGGATAACAATT CTAGAGAA
-19 +I t10 t 2p
-I? t 1 ‘I? e 2 0
~TATATTJCCTCGA ACTAGT~AACTAGT
TCGAGG
AA [ T T ~ T C T A G +AT c A C C G ~ ~ AGTGGTA CTA GAG A A*I 2
P
‘2,O&%~iiiTcGp. CTCTATCATTGATAGAG TCGAGGAA
4
t I?TCGAACTAGTTAACTAGTTCGA
TCG AACTAGTTA ACTAGTBCGC
EGIACTAG&ACTAGTGCM
-1p +!
TCGIACJCTTTAFSAGTACAA
-2p -I,O
-y
-3,OY
tIp
CTCTATCATTGATAGAGFIGURE 4.-Design of operator substitutions on challenge phages. a , Positions of cloned asym- metric wild-type Lac and OLI operators, and idealized, symmetric T r p (“trp”) and Tet (“tet”) op-
erators with respect to the ant promoter. “+1” designates the base pair corresponding to the wild- type P,,, startpoint. The conserved Pant -10 hexamer is boxed. b, Comparison of the consensus T r p operator (“trp”) with the three naturally occurring T r p operators controlling the trp, aroH
and trpR operons (adapted from GUNSALUS and YANOFSKY 1980). The positions of each operator with respect to the promoters they control are indicated. Differences from the idealized sequence are underlined. c, Comparison of the consensus Tet operator with the two operators controlling the tetA promoter (see DANIELS and BERTRAND 1985).
virulent progeny able to form plaques on an immune (P22 c2+ mnt+) lysogen. To distinguish recombinant challenge phages from mutants of the parental P22 K n 9 arc-am phage that had acquired mutations in the Mnt operator ren- dering the synthesis of antirepressor constitutive, challenge phage DNA was digested with endonuclease EcoRI to verify the presence of the additional
EcoRI site within the operator substitution. T h e precise structures of the op- erator substitutions were confirmed by DNA sequence analyses.
8 N. BENSON E T AL. TABLE I
Substitution of the Lac operator for the Mnt operator confers new immunity properties on P22
Bacterial host
MS1363 MSl367 MSl582 MS1367/pMB9 MS13671pMC7
Phage (none) ( C Z + ) (c2+mnt+) (c2') (cZ+lacI+)
KnY Om,, arc-am 1
.o
0.9 1.o
1.1K n 9 01,, arc-am 1
.o
1.o
0.7 0.9 4 0 - 5Phage suspensions were diluted and plated on the indicated bacterial strains. Plaques were scored after incubating for 12 hr at 37". Efficiencies of plating on each bacterial strain are the titers on each indicator divided by the titer on MS1363. All bacterial hosts carry the supE40 amber suppressor allele and produce the repressors indicated below the strain designations.
TABLE 2
Survival upon challenge phage infection depends on the amount and activity of repressor
Concentration of IPT G (M)
Bacterial host 0 I 0-5 10-4 10-3
MS 1868 -a 0-7 <lo-?
MSl868/F'lacI+ 2 x 1 0 - ~ 2 x 10-5 4 0 - 7 <io+
MS1868/F'lacP1 0.5 0.02 3 x 10-7 2 x
MS1868/pMC7 0.5 0.5 0.5 0.2
Cells were grown in LB medium at 37" to a density of 2 X 108/ml, diluted fourfold, grown two generations in the presence of the indicated amounts of IPTG (isopropy~-@D-galactopyrano- side) and infected with P22 KnY Ole, arc-am phage at a multiplicity of infection of 20. After 20- min adsorption at 2 5 " , infected cells were diluted and spread on plates containing the indicated amounts of IPTG. Entries in the table are the efficiencies of survival of the hosts following challenge phage infection, defined as the titer of each infected cell culture assayed on green/kan plates divided by the titer of input cells assayed on green plates.
superinfecting phage must synthesize antirepressor and inactivate c2 repressor produced by the prophage in order to grow (BOTSTEIN et al. 1975). However, the parental phage cannot form plaques with normal efficiency on MS1582, a lysogen that produces both c2 and Mnt repressors ( G R A ~ A , YOUDERIAN and SUSSKIND 1985). Likewise, the challenge phage with the Lac operator cannot form plaques efficiently on MS1367 carrying a derivative of pMB9 that pro- duces Lac repressor, pMC7 (CALOS 1978). These hosts are homoimmune to phages with the binding sites corresponding to the repressors they produce. Consistent with this idea, mutants of P22 Kn9 Olac arc-am phage that form plaques efficiently on the "Lac-immune" Salmonella host are found to carry constitutive mutations that inactivate the Lac operator (data not shown).
Selective recovery of cells that produce Lac repressor: When a sensitive
TABLE 3
Cells that produce specific DNA-binding activities may be recovered from
mixed populations using the challenge phage selection
~
~~ ~~ ~-
~~
Relative concentration
2 x 10-6 2 x IO-* 2 x 10-5
Expected survivors (520) (265) (27)
Efficiency of recovery 0.3 0.2 0.2
Observed survivors 165 57 5
Cultures of MS1868/pMC7 (lac@ and MS1868/pMB9 (lacl-) were grown to densities of 3 X 10’ cells/ml in LB/tet medium at 37”. The MS1868/pMC7 culture was serially diluted and added to aliquots of the undiluted MS1868/ pMB9 culture to give mixed cultures with the indicated relative concentration of MS1868/pMC7. Aliquots of the mixed cultures were infected with P22 Kn9 Ole, arc-am phage at a multiplicity of infection of 20, incubated at 25” for 20 min to ensure efficient adsorption and then distributed to green/kan/tet plates. Colonies representing observed challenge phage survivors were scored after incubating for 24 hr at 37”. The number of expected survivors was calculated from the titer of input MS1868/pMC7 cells. All five survivors of the most dilute mix were determined to carry pMC7 upon further analysis.
duces Lac repressor to survive challenge phage infection is dependent on both the amount and activity of the Lac repressor produced by the host. As shown in Table 2, a host carrying the high copy-number plasmid pMC7 that produces Lac repressor from the strong lacZQ promoter is lysogenized by the challenge phage even at high concentrations of inducer. In contrast, when Lac repressor is produced from a single-copy lade’ promoter carried on an F’ episome, a high concentration of inducer prevents lysogeny of the challenge phage. T h e amount of repressor produced from the wild-type l a d promoter in single copy (F’lac+) is sufficient to permit lysogeny with reduced efficiency in the absence of inducer.
How efficiently may the challenge phage selection be used to recover rare clones that produce Lac repressor from a mixed population of cells? A recon- struction experiment was performed to answer this question. We mixed various proportions of supo cells carrying pMB9 with otherwise isogenic cells carrying pMC7. As shown in Table 3, infection of mixed populations with challenge phages permits the selective recovery of cells carrying pMC7 ( l a d + ) us. cells carrying pMB9 (lacZ-) at frequencies as low as one in a million on a single plate. It follows that the challenge phage selection may be used to recover clones that produce a specific DNA-binding activity efficiently from large, mixed populations of cells; that is, from plasmid o r phage expression libraries that include rare clones producing specific DNA-binding activities.
10 N. BENSON E T A L . TABLE 4
The challenge phage selection works for a wide variety of DNA-binding proteins
Challenge phage operator
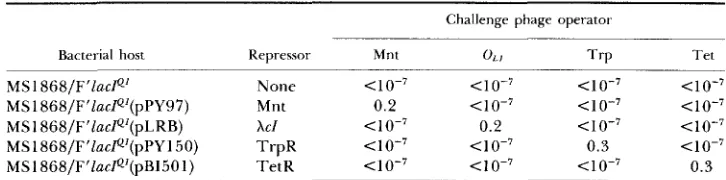
Bacterial host Repressor Mnt 0 1 1 T r p T e t
MSl868/F’ZacP’ None 4 0 - 7 4 0 - 7 < I 0-7 <io-’
MSl868/F’lacP’(pPY97) Mnt 0.2 4 0 - 7 <IO-’ < 10-7
MS 1868/F’lacP‘(pLRB) X C I <io-’ 0.2 < i o - 7 < 10-7
MSl868/F’Zac~’(pPY150) T r p R 4 0 - 7 <IO-’ 0.3 <IO-’
MS1868/F’Iac~’(pB1501) TetR 4 0 - 7 <IO-’ 4 0 - 7 0 . 3
Cultures of MSl 868/F’lacP1, MSl868/F’Zac~’ (pPY97), MSl 868/F’ZacP1 (pLRB), MS1868/ F’lacP’ (pPY150) and MS1868/F‘ZacPL (pB1501) were grown at 37” in LB medium to a density of 5 X 108/ni1, diluted fourfold, grown two generations in the presence of M IPTG (to induce the synthesis of the particular plasmid-coded repressor protein) and infected with a multiplicity of
20 of each challenge phage. After 20-min adsorption at 2 5 ” , infected cells were diluted and spread on green/kan plates containing M IPTG. Numbers in the table are the efficiencies of survival
of host cells following challenge phage infection, defined as the titer of each infected cell culture assayed on green/kan plates divided by the titer of input cells assayed on green plates.
and TnlO Tet operators selectively lysogenize hosts that produce their respec- tive repressors. It is clear that the challenge phage selection can be applied to a wide variety of specific DNA binding site/binding protein interactions.
Moreover, synthetic, symmetric versions of the T r p and Tet operators be- have as natural operators should. Together with their cognate repressors, these “idealized” operators can effect repression of the ant operon. These findings are consistent with those of SADLER, SASMOR and BETZ (1983), who demon- strated that a symmetric version of the Lac operator binds wild-type repressor with tenfold higher affinity than does the asymmetric, wild-type operator. Re- cently, we have determined that the operator of SADLER, SASMOR and BETZ (1983), a symmetric version of the X operator, and an “idealized,” symmetric version of one of the E. coli Gal operators are sensitive to each of their cognate repressors, as determined by the challenge phage selection (data not shown). Therefore, we expect that the study of many repressor/operator interactions may be modeled using symmetric or nearly symmetric operator sites.
DISCUSSION
T h e majority of prokaryotic repressors that recognize specific sequences of D N A are thought to act by binding to operator sites that overlap the RNA polymerase binding sites (promoters) they control. Presumably, when a repres- sor binds its operator, it sterically excludes the binding of polymerase to the promoter; that is, repressors are competitive inhibitors of R N A polymerase binding. Consistent with this idea, we have shown that five different operators can function at the same site within the P22 ant operon (the startpoint of transcription).
1985). We have subcloned both the Lac and Tet operators at the startpoint of ant promoter transcription, at positions analogous to those within their original operons, and have shown that these operators function when paired with the heterologous ant promoter. T h e locations of the T r p and XOL, op- erators within their wild-type operons are different from the location of these operators on challenge phages (with respect to conserved promoter sequences). In both cases, the site still functions as an operator when placed at the start- point of transcription. T h e operons controlled by X and T e t repressors have reiterated operators; in both cases, we have shown that a single, minimally defined operator site suffices for repression of an operon.
In previous reports (YOUDERIAN et al. 1983; VERSHON et al. 1985b), we demonstrated that, starting with a symmetric mutant operator unable to bind wild-type P22 Mnt repressor, we can select mutant repressors that recognize this mutant operator. This simple genetic strategy for selecting mutant repres- sors with altered specificities of binding may be applied systematically and exhaustively to a single symmetric operator. One can synthesize all possible symmetric variants of a reference, symmetric operator; introduce this set of sites into a challenge phage vector; identify the subset of mutant operators that fail to bind wild-type repressors; and isolate mutant repressors that bind each mutant operator.
In this paper, we have established that challenge phages with idealized, sym- metric versions of the T r p and Tet operators can be used to select for binding of the wild-type T r p and Tet repressors (see Figure 4). Experiments are in progress to identify the genetic determinants of each of these DNA binding interactions in a thorough manner. More recently, we have shown that ideal- ized, symmetric versions of the X and Lac operators may also be used as starting points for the analyses of these two interactions (data not shown).
We have also set the stage for the systematic isolation of eukaryotic DNA- binding proteins from expression libraries, starting with the sites they recog- nize. Eukaryotic binding sites can be introduced onto challenge phages with relative ease. Provided that their corresponding binding proteins can be ex- pressed in sufficient amounts and activities in Salmonella hosts, we can select bacterial clones that produce these proteins from large, mixed populations of cells using challenge phages.
12 N . BENSON E T A L .
determine precisely how induction ratio varies as a function of increasing distance between a promoter and an operator site placed from one to 17 base pairs 3’ relative to the transcription startpoint.
In summary, we have shown that the challenge phage selection may be applied to a wide variety of specific, prokaryotic DNA-binding interactions and have laid the foundation for extending the selection to the isolation of eukar- yotic DNA-binding proteins.
We a r e especially indebted to ROBERT GUNSALUS for providing us with plasmid pRPG47, and t o PAUL ISACKSON and KEVIN BERTRAND for providing us with pBI501 and pPIlOOT before publication. We a r e also grateful to RICHARD DEONIER, DAVID GALAS, PONZY Lu and ROBERT SAUER for gifts of synthetic oligonucleotides, plasmids and bacterial hosts. We thank PETER BERGET a n d MIRIAM SUSSKIND for their detailed criticisms of the manuscript. This research was supported by National Institutes of Health grant GM34150. P.S. WAS the recipient of a 1984 summer fellow-
ship from the Student Research Associates Prograni of the California Heart Association.
LITERATURE CITED
AMANN, E., J. BROSIUS and M. PTASHNE, 1983 Vectors bearing a hybrid trp-lac promoter useful
APPLEYARD, R. K., 1954 Segregation of new lysogenic types during growth of a doubly lysogenic
ARVIDSON, D. N., B. CAN and R. P. GUNSALUS, 1985 Interaction of the Escherichia coli t r p aporepressor with its ligand L-tryptophan. J. Biol. Chem. 261: 238-243.
BOLIVAR, F., R. L. RODRIGUEZ, M. C. BETLACH and H . W. BOYER, 1977 Construction and characterization of new cloning vehicles. I. Ampicillin-resistant derivatives of the plasmid pMB9. Gene 2: 75-93.
for regulated expression of cloned genes in Escherichia coli. Gene 25: 167-178.
strain derived from Escherichia coli K12. Genetics 39: 440-452.
BOTSTEIN, D., R. K. CHAN and C. H. WADDELL, 1972 Genetics of bacteriophage P22. 11. Gene order and gene function. Virology 49: 268-282.
B o r s r E m , D., K. K. LEW, V. JARVIK and C. A. SWANSON, JR., 1975 Role of antirepressor in the bipartite control of repression and immunity by bacteriophage P22. J . Mol. Biol. 91: 439- 462.
Virulent mutants of bacteriophage P22. I. Isolation and BRONSON, M. J. and M. LEVINE, 1971
genetic analysis. J. Virol. 7: 559-568.
DNA sequence for a low-level promoter of the lac repressor gene and a n “up” promoter mutation. Nature 274: 762-765.
Chemical synthesis of oligodeoxyribonucleotides using the phosphite triester intermediates. pp. 7 1-79. In: Chemiral and Enzymatic Synthesis of Gene Fragments, Edited by H. G. GASSEN and A. LANG. Verlag Chemie, Weinheim, FRG.
Promoter mutations affecting divergent transcription in the T n l O tetracycline resistance determinant. J. Mol. Biol. 184: 599-610.
(;RAGA, D., P. YOUDERIAN and M. M. SUSSKIND, 1985 Mutations that iniprove the ant promoter of Salmonella phage P22. Genetics 110: 1-16.
Nucleotide sequence and expression of Escherichia coli trpR, the structural gene for the trp aporepressor. Proc. Natl. Acad. Sci. USA 77: 71 17-7121. Activation and repression of the XPRF promoter by cI1 protein. pp. 229-238. In: Sequence Speczficity i n Transcription and Translation, Edited by R. CALENDAR and L. GOLD. Alan Liss, New, York.
Dominant negative mutations in the T n l O tet re- CALOS, M., 1978
CARUTHERS, M. H., 1982
DANIELS, D. W. and K. P. BERTRAND, 1985
GUNSALUS, R. P. and C. YANOFSKY, 1980
GUSSIN, G . , J.-J. HWANG, K. M A n , M . ZURER and D. COURT, 1985
pressor: evidence for use of the helix-turn-helix motif in DNA binding. Proc. Natl. Acad. Sci.
Transformation of Salmonella typhimurium by plasmid
Mutations in the temperate phage P22 and lysogeny in Salmonella. Virology
Genetic fine structure of the C region and the linkage map
Dual control of lysogeny by bacteriophage P22: an antirepressor locus and its controlling elements. J. Mol. Biol. 91:
DNA restriction enzyme from E. coli. Nature 217: 1110-
X-repressor mutations that increase the affinity and
Control of transcription by the bacteriophage P22 re-
Operator sequences of bacteri-
A mechanism for USA 82: 6226-6230.
LEDERBERG, E. M. and S. N. COHEN, 1974
deoxyribonucleic acid. J. Bacteriol. 119: 1072-1074.
LEVINE, M., 1957 3: 22-41.
LEVINE, M. and R. CURTISS, 1961 of phage P22. Genetics 46: 1573-1580.
LEVINE, M., S. TRUESDELL, T. RAMAKRISHNAN and M. J. BRONSON, 1975
42 1-438.
MESELSON, M. and R. YUAN, 1968 11 14.
NELSON, H. C. M. and R. T. SAUER, 1985 specificity of operator binding. Cell 42: 549-558.
pressor. J. Mol. Biol. 157: 21-48.
ophages P22 and 21. J. Mol. Biol. 137: 81-91.
repressor action. J. Mol. Biol. 43: 201-213.
RODRIGUEZ, R. L., F. BOLIVAR, H. M. GOODMAN, H. W. BOYER and M. BETLACH, 1976 Construction and characterization of cloning vehicles. pp. 47 1-477. In: Molecular Mech- anisms in Control of Gene Expression, Edited by D. P. NIERLICH, W. J. RUTTER and C. F. Fox. Academic Press, New York.
A perfectly symmetric lac operator binds the lac
DNA sequencing with chain-terminating inhibi- POTEETE, A. R. and M. PTASHNE, 1982
POTEETE, A. R., M. PTASHNE, M. BALLIVET and H. EISEN, 1980
REZNIKOFF, W. S., J. H. MILLER, J. G. SCAIFE and J. R. BECKWITH, 1969
SADLER, J. R., H. SASMOR and J. L. BETZ, 1983
SANGER, F., S. NICKLEN and A. COULSON, 1977
repressor very tightly. Proc. Natl. Acad. Sci. USA 8 0 6785-6789.
tors. Proc. Natl. Acad. Sci. USA 7 4 5463-5476.
SAUER, R. T . , W. KROVATIN, J. DEANDA, P. YOUDERIAN and M. M. SUSSKIND, 1983 Primary
Recombination in bacteriophage A. I. Mutants deficient in
A new gene of bacteriophage P22 which regulates synthesis of antire-
Mechanism of action of Salmonella phage P22 antire-
Superinfection exclusion by P22 prophage in lysogens of Salmonella typhimurium. 11. Genetic evidence for two exclusion systems. Virology
The bacteriophage P22 Arc and Mnt repressors: overproduction, purification, and properties. J. Biol. Chem. 2 6 0 121 24-12129.
structure of the imml immunity region of bacteriophage P22. J. Mol. Biol. 168: 699-713. SIGNER, E. R. and J. WEIL, 1968
general recombination. J. Mol. Biol. 34: 261-271.
pressor. J. Mol. Biol. 138: 685-713.
pressor. J. Mol. Biol. 98: 413-424. SUSSKIND, M. M., 1980
SUSSKIND, M . M. and D. BOTSTEIN, 1975
SUSSKIND, M. M., A. WRIGHT and D. BOTSTEIN, 1971
45: 638-652.
VERSHON, A. K., P. YOUDERIAN, M. M. SUSSKIND and R. T. SAUER, 1985a
14 N . BENSON E T A L .
VIEIRA, J. and J. MESSING, 1982 T h e pUC plasmids, an M13mp7-derived system for insertion mutagenesis and sequencing with synthetic universal primers. Gene 19: 259-268.
WEINSTOCK, G., M. M. SUSSKIND and D. BOTSTEIN, 1979 Regional specificity of illegitimate recombination by the translocatable ampicillin-resistance element T n l in the genome of phage P22. Genetics 92: 685-710.
Sequence determinants of promoter
Changing the
YOUDERIAN, P., S. BOUVIER and M. M. SUSSKIND, 1982 activity. Cell 30: 843-853.
DNA-binding specificity of a repressor. Cell 35: 777-783.
YOUDERIAN, P., A . VERSHON, S. BOUVIER, R. T. SAUER and M. M. SUSSKIND, 1983
ZAGURSKY, R. J. and M. L. BERMAN, 1984 Cloning vectors that yield high levels of single-stranded