Changes
in
Human
Mucosal
y6
T
Cell
Repertoire and Function
Associated
with
the
Disease Process in
Inflammatory
Bowel Disease
Laila
D.McVay,*
Baiqing Li,*
ReneeBiancaniello,*
Mary Anne
Creighton,* Dale
Bachwich,A
Gary
Lichtenstein,*
John L.
Rombeau,§
and
Simon R.
Carding*
*Department
of
Microbiology, tDivision of Gastroenterology, and
§Department
of Surgery,
University of Pennsylvania School of
Medicine,
Philadelphia, Pennsylvania 19104-6021, U.S.A.
ABSTRACT
Background: Although y6T cells are amajor compo-nentofthe human intestinal mucosa, it is not clear what role they play in mucosal inimunity or ifthey are in-volved in the disease process of inflammatory bowel disease (IBD).
Materials and Methods: Flow cytometry and reverse transcriptase-polymerase chain reaction (RT-PCR) as-says were used to identify quantitative and qualitative changesinthe repertoire of-y8Tcells presentinsurgical and/or biopsy samples of normal and inflamed colon from individual patients with ulcerative colitis (UC) or Crohn's disease (CD). Cytokineproductionand the abil-ity toadhere to and interact withcolonicfibroblasts were used tocompare the functionalproperties of yST cells isolated from the normal and diseased colonic mucosa. Results: Increased numbers of yS T cells localized in areasof inflammation andtissue injurywere found in
the majority of patients, irrespective of the typeofIBD present.This expansion was attributable to an increase in V61+ cells expressing a V5I-(D63)-J&1-encoded T cell receptorand was seen in patientswithseveredisease as well asthosewith newly diagnosedorlesssevereforms of IBD. Among T cells present in the inflamedmucosaof patientswith CD, y8Tcells, particularlyV81+cells, were a major source of the proinflammatory cytokine inter-feron--y and couldinteractwith colonic fibroblasts. Conclusions: Ourresults demonstrate thatthe chronic inflammatoryimmune responsecharacteristic of IBD is associatedwith distinctchanges in thenumber, distribu-tion, composidistribu-tion, and function of mucosal 'yS T cells. Through the production of cytokines andphysical inter-actionwithothercells, y8Tcells canperforman immu-noregulatory function and contributetothe pathophys-iology of IBDs.
INTRODUCTION
Inflammatory bowel disease (IBD) represents a complex group of idiopathic chronic inflamma-torydisorders of which Crohn's disease (CD) and ulcerative colitis (UC)arethe majorforms. While thereareseveralfeatures that distinguishCDand
Addresscorrespondence and reprint requests to:SimonR.
Carding,Department ofMicrobiology, 802FAbramson
Center, 34thStreet & Civic CenterBlvd.,Philadelphia, PA
19104, U.S.A.Phone:215-573-3022; Fax:215-573-9068;
E-mail: [email protected]
© 1997,THEPICOWERINSTITUTE PRESS. Allrightsreserved.
MolecularMedicine,Volume 3, Number 3, March 1997 183-203
UC, there aremanyepidemiological, clinical,
ra-diologic, endoscopic, and histologicfeatures that are shared between these two disorders.
Several immunologic and histopathologic
features of UC and CD can be explained as a
defect in mucosal immunoregulation, and as a direct or indirect consequence ofpersistent mu-cosal T cell activation (1-4). Experimental evi-dence for a direct role for T cells in IBD comes
from various animal models, especially "gene
knock-out" strains that spontaneously develop
bowel lesions. In particular, the presence of in-testinal inflammation in transgenic mice defi-cient inaC3Tcells (5) orTcell-derived cytokines
such as interleukin 2 (IL-2) (6) suggest that T
cells are responsible for the homeostatic
regula-tion of mucosal immune responses.
Inthehuman intestine,
y&'
cells areamajor component (30-40%) of theTcellsintheintra-epithelial spaces and the majority (>70%)
ex-press V6I-encoded receptors (7). Inview of the predominance of V61+ cells within the human
intestinal mucosa and the possibility that they have similar effector functions to their murine
counterparts, it seems likelythat they may play
an important role in the generation and/or
reg-ulation ofinflammatory responsesinthe
intesti-nal mucosa. The ability of yST cells to interact
with and influence the activity of a variety of
stromal cells (8-10), including epithelial cells
(8,1 1), and thefindingthat the severity ofIBDin Tcell receptor (TCR)-a/3 deficient miceis signif-icantly reduced when crossed withmice lacking ,y6 T cells (5) are all consistent with an immuno-regulatory role for mucosal 'y6T cells.
Whereas several groups of investigators have described an increase in the number of V81+
intraepithelial (IE) y8Tcells within the intestine of patients with coeliac disease (12-16), the
im-munohistochemical analyses of mucosal yB T
cellsinpatientswithIBDhave generated
contra-dictory results (16,17). Although one group has shown that thedistribution and relative number of TCR-5+ cells in sections of colonic mucosa
obtained fromIBDpatients was similar to thatin
sections of normal colonic mucosa (16), the re-sults of another study showed that there were fewer TCR-6+ cells in the inflamed mucosa of patients with IBD, and particularly, those with
UC (17). Thus, evidence for the involvement of
y8Tcells inIBDremains inconclusive. To inves-tigate furtherthe possible involvement of muco-saly6TcellsinIBD wehave useda combination ofsensitivecellular and molecular techniques to determine if thereareany qualitative or
quanti-tative changes in y8 populations in the colonic mucosa of patients with CD and UC.
The results of our studies identify changes in the number, distribution, and repertoire of
y6
T cells that are present within the lesions of the inflamedintestine inboth UC and CD. The findingthat,incontrastto
y8
Tcellspresentinthehealthy, noninvolvedmucosa, yScellsintheinflamedmu-cosaconstitutively produce theinflammatory cyto-kine interferon y (IEFN-,y) and selectively interact with colonic stromal (fibroblasts) cellssuggest that
mucosal
y6
Tcells can contribute to thedevelop-ment of thedisease processin IBD.
MATERIALS
AND
METHODS
ProcurementandProcessing of
Patient
Samples
Surgical resectioned segments of intact colon
were obtained from patients with UC, CD, or
diverticular disease (diverticulitis) atthe time of elective surgery. Histologically normal,
nonin-volved, mucosa was obtained from the
margins
of
surgical
resectioned colon that were atleast adistance of 15 cm from any lesions. Mucosal
biopsies
were obtained with the approval of theHumanInvestigations Committee of the
Univer-sity of Pennsylvania School of Medicine. The
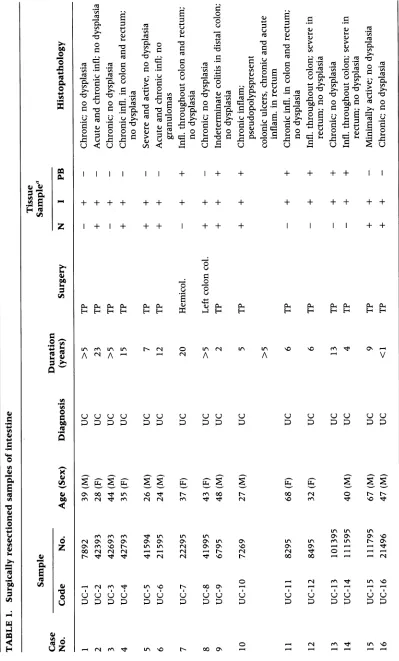
origin andnature of the patient samples used in this studyare compiled in Tables 1 and 2. When
available, peripheral blood mononuclear cells (PBMCs) from the same patient were used as
control samples. Patient-derived samples were
used for RNA extraction, reverse transcriptase-polymerasechain reaction (RT-PCR) analysis, in situ hybridization, and/or cell isolation for flow
cytometric andfunctional analyses.
CellIsolation
Peripheral blood mononuclear cellswereisolated from 5 to 10 ml of peripheral blood by density gradient centrifugation using Ficoll-Hypaque (Organo-Teknika, Durham, NC) and washedone totwotimes withPBS. Established human fibro-blast cell lines of skin (CRL-1509), lung
(CCD-1llLu), or colon (CCD-18Co) origin were ob-tained from the American Type Culture Collection (ATCC, Rockville,MD). Aprimary
co-lonic fibroblast cell line was derived from the involved mucosal tissue fragments of a patient with ileocolonic CD (CD-11, Table 1) as de-scribed previously (18). This linewasmaintained inDulbecco's modified Eagle's medium (DMEM) containing 10%fetal calfserum(FCS) and penicil-lin/streptomycin/bactrim/amphotericin B. Priorto
reachingconfluency, cells weretrypsinized and di-luted 1:5 in complete media and platedinto tis-sue culture-treated 20-mm petri dishes.
Adher-ence assays were performed with primary
fibroblasts that had beenpassaged twice.
IEand Lamina Propria (LP) T Cells
A modified version of a previously published procedure (19) was used to isolate IE and LP
lymphocytes (L). Briefly, after removal of all
as-sociatedmesentery, fat, and underlying
L. D. McVayet al.: y6TCells andInflammatoryBowel Disease
and cut into 2- to 3-cm pieces. IE lymphocytes
wereeluted by 3to4sequentialincubations with
Hank'sbalanced salt solution (HBSS) containing
0.1 mM EDTA at 37°C each for 15 min with
constantmagnetic stirring. The eluted cells were
then subjectedtotwo-stepdiscontinuous Percoll
(Pharmacia-LKB, Uppsala, Sweden) gradient centrifugationat 600 X gfor 20min using 70%
and 40% Percoll. IEL were removed from the
interface between the70% and40% Percoll
lay-ers. The remaining tissue fragments were incu-bated with HBSS containing collagenase typeIV
(Sigma, St. Louis, MO) at 370C for 4-6 hr with constant stirring. The recovered LP cell
suspen-sion was then subjected to Ficoll-Hypaque den-sity gradient centrifugation to isolate viable cells from which LP lymphocytes were obtained by
Percoll gradient centrifugation as described
above for IEL isolation. The yield of LP T cells
(CD3+ cells) from the noninvolved mucosa and
fromthe involved mucosa was calculatedas the
number of cells per gram of tissue used for
iso-lation, and this yield was routinely comparable to that reported by others (20,21).
IMMUNOMAGNETIC SELECTION OF MUCOSAL IEL AND LP
7y+
TCELLS. Anegative-immunomagneticselec-tionprocedure (22) was used to obtain
homog-enous populations of
TCR-,yS+
or TCR-aIB+ IELandLP cells from theinvolved and noninvolved
mucosa of colonic surgical specimens. Briefly,
total LP and IEL cell samples were sequentially
incubated with a mouse anti-human TCR-ac
(clone BW242/412; T CellDiagnostics, Woburn,
MA) or a mouse anti-human TCR--y6 antibody
(11F2, Becton Dickinson) and a rabbit
anti-mouseimmunoglobulin G (IgG) antibody
conju-gatedto supramagnetic particles (BioMagBeads,
Perseptive Diagnostics, Cambridge, MA). The
cells were then transferred to a 25-cm2 tissue
culture flask and placed on a plate magnet and
the "nonmagnetized" cells were removed. The
purity of eachT cellpopulation wasdetermined
by flow cytometry, and cell viability, by trypan blue dye exclusion.
Flow Cytometry
Aliquots (0.2-1.0 X 106) of bulkorfractionated
IEL and LP mononuclear cells were stainedwith
fluorochrome-conjugated mouse anti-human
CD3- (clone S4.1; Caltag Labs., San Francisco,
CA), CD8a- (3B5, Caltag), TCR-y8- (l1F2,
Bec-ton Dickinson), TCR-Vy9- (T Cell Diagnostics), TCR-V81- (TS8.2; T Cell Diagnostics),
TCR-V61-J31- (8TCS-1; T Cell Diagnostics) and/or
TCR-V52- (23) specificantibodies, fixed andanalyzed
on a FACScan (Becton Dickinson) using Lysis
IIor CellQuest softwareasdescribedpreviously (23).Fluorochrome-conjugated isotype-matched
mouse antibodies or irrelevant specificity (Caltag)
and anti-human CD45-specificantibodies (Dako
Corporation, Carpinteria, CA) wereusedto
con-trol for nonspecific reactivity and as fluoro-chrome-compensation controls, respectively.
RT-PCR Analysisof the TCR-VS
Repertoire
Total cellular RNAwas preparedfrom intestinal tissueor PBMC byCsCl density gradient centrif-ugation (24). RNA pellets were extracted with acidic phenol once, with phenol:chloroform
(1:1) once, and twice with chloroform prior to
ethanol precipitation. Approximately 10 ,tg of total cellular RNA wasused to synthesize cDNA
using random hexanucleotides as described
pre-viously (25). Anested PCR assay (24) was used
to analyze the
V8-J6
rearrangements expressed byy6
T cells in colonictissue and bloodmono-nuclear cell RNA/cDNA samples. Briefly, oligo-nucleotide primers specific for V81, V82, or V53
in combination with a CS gene segment primer
were used to amplify cDNA. The products of
amplification were electrophoresed and
TCR-PCR products gel purified and quantitated by
DNA fluorimetry. Equivalent amounts of each
cDNA were usedas a template for a second am-plificationusingthe same forward VS primer and
a reverse-primer specific for each of the four human JS genes. The products of amplification
were electrophoresed on 1.8% agarose gels, Southern blotted, and hybridized with
[y32p]
-ATP end-labeled VS1, V82, or V83 oligonucleo-tide probes containing sequence internal to the VA primers used for PCR. Individual V1I-JS1-4,V62-J61
-4, andV83-J61
-4receptor cDNAcloneswere usedto determine the optimum conditions for theproportional amplification ofVS-JS genes with no cross-priming. For analysis of mucosal biopsy specimens, 5 to 6 individual pieces of
tissue were pooled and total cellular RNA was
isolated as described above. After generating cDNA,aliquotswereanalyzedforV 1-J5
expres-sion by PCR amplification and Southern blot
analysis asdescribed for the surgical samples.
-4 0 u r-4 0 0 u 4--. 4-4
v P-1 P64
14 F-4 F-4
+ + + + + + -4 0 u .1.4 5
P-4 PL-4 P-4 P-4 0.4 P-( Qi
F-4 F- F-4 F-4 F-4 1-4 z F-4PL4 P-4F-4 F-4P-4 P-4H P-4F-4 P-4F-4
vlE m Ul ulE
A N A
-r- rq 0 ull. rq
-4 rq A
UII vl
A "lso\0 _-- It c7 V
_-4i 0 .P.4 4-) rA (10 4-. 0 0 (A qj W-4 04
5
(10 (A PC (U 0 0 .P.q 4-i u w rA 4i 1.4 P-4 P-4 tv w .P.4 to k :3 .A 1; w 0.4 m E-4..) u u u u u u u u u u u u u u u
.D .D .:) .D .D .D .D .D .D .D .D .D .D .D .:) .D
I--_0 ,I oo C) I'l r-_ _q W-1 r-m tn 0\ N rq rq W-oo -4 vl. C\ r-F--4 -4 F--4 LrN r..4I u -4 6 z qj "O 0 Q
N 0\ ol O\
oo N N N
r- "d Rt I'l ,o
o\ \o Ull (ON oo VUE as~
oo4
ai P-4 045
kV.A --4 N M "14 ul. \0 t- oo (ONi O-4 N lq
6 6 6 6
6 6
6
.D .:) .:) .D .D .D
u z 0 1- N m lqt vl 'IO
,--r- M t uE \0 - 00 0\ -4 -4 P-4 1-
1-,DO_ <, - E x;
~~~~~~~~~~-
-4 uE
++
++++ ++ + ++ +
++~~~~~~~4-
+ +++
-( .-
-- a
x
oSc
aD)
r. 7~ 0 (0
4-r,a r, 0 co ['t -A
S434EvE4 _4S- (a CL (-a ((a4
~~~~~~-
S-
ga I .( . . -[au
~~-4 )-4 --40
;-4~~00
~~~~.4--4;-4~~~~~~~~~~~~~~~~~~~~~~
a~~~~~VC)Q.
*4; j-4j;)~
(-a(-a
(-a ~ ~-4[I a-4 U00( (> (>>
4-~A>
4-HH
4-4+ + + + + + + + + + + +
+ + + + + + + +
4a)Q a)a a)-) a-)a)
u oD u u
U] (A UA VA
u u u u
9 9 P4 W
(0 0 *-4 a.) (ad P4
- - V- Utt N
N V A N
(. 0 -4 v] a)
P)
(0 0 v! Q.) A A S 0 0 u S 0 a-) U \O Vr AU U U U U U U U
zzPO kJ U UAA UU
NO ON t tIN UN
-- A A VV V
(0
a)
r~
r~
~ a) :Za) .~A
't
N O0
rs
o ON oN aON ON ON 0 ON
N O - N NO
m I'l It lt N
- N It [
U)UUU6 6
N 00 ON 0
--- - N N
00 't ON -4 \O NO O-ulE -CN ulE (N 00 \N 00 0 *-S:z -;-4 -4 OC) 0 00 V) *-S -a) .> m
(0
1-0 u 00 m 0 U (0 00 It ONON N N-00 CN4 NIINt~t t_- 0-0 00 U--Nrq N
2 2z
[INul NO N-00 OIN 0= - (r It
rq N ri (N(N (N rIrI rIcn INr
-TABLE 2. Biopsysamples ofintestinal tissue
Sample Tissue Sample
Case Duration
No. Code No. Age (Sex) Diagnosis (years) N I PB
1 Bl-UC 82494 26 (M) UC 15, recurrence + +
2 B2-UC 21495 25 (M) PanUC 12, recurrence + +
3 B3-UC 11295 37 (M) Nonspecific colitis 1 week + + +
4 B4-UC 72194 (M) UC 5, recurrence - +
5 B5-UC 12795 47 (F) Active UC 5-6, recurrence - +
6 B6-UC 9894 66(M) Noninvolved UC 13, recurrence +
7 B7-UC 3895 28 (M) UC 5, recurrence - + +
8 B8-CD 72794 (F) CD 2, recurrence - +
N,noninvolved; I, involved;PB,peripheralblood;UC, ulcerativecolitis;CD, Crohn'sdisease.
In Situ Hybridization
In situ hybridization was used to identify and localize populations of y8 T cells and IFN-,y-pro-ducing cells within frozensections ofintestine as
described previously (22,26,27). Briefly, 1- to
2-cm2 pieces of tissue from involved or
nonin-volved areas along the length of the resected intestine were snap frozen inTissue Tek (Miles, Elkhard, IN) and stored at -860C. Six-10 ,um
sections were cut from tissueblocks usinga cry-ostat, fixed in4% paraformaldehyde inPBS, pH 7.4,for20 min at roomtemperature, then stored
in 70% ethanol at 4°C in airtight containers.
35S-labeled RNAprobes (0.2 ng
RNA/,li;
specific activity >109 cpm/,tg) complimentary to thecoding (antisense) or noncoding (sense)
se-quences human IFN-,y (26) genes were used for detection of TCR- and
IFN--y-mRNA+
cells.IFN-y Assay
IFN-'y protein production was analyzed from
bulk, nonfractionated T cells and purified ac3 T
cells, and y5 T cells isolated from segments of noninvolved or involved mucosa from two
pa-tients with ileocolonic CD and one with colonic
CD. Two hundred thousand cells were cultured
in triplicate in microtiter plate wells containing either mediaalone orwells coated withan
anti-CD3 antibody (OKT3). After 96 hr, the culture supernatantwasharvestedandassayed for IFN-,y usingacommercial radioimmunoassay (RIA) kit (Centocor, Malvern, PA). The amount ofIFN-y present was determined by reference to a
stan-dard curve generated from samples containing knownamounts of rIFN-'y assayed inparallel.
Fibroblast Adherence Assay
T cells (3 X 106) from peripheral blood and/or
normal orinvolved colon from four patients with ileocolonic CD were allowed to adhere to
con-fluent monolayers of established or primary fi-broblast cell lines in20-mm petridishes at 370C
for30 min. Nonadherent cellswere removedby gentle washing with prewarmed media and
ad-herent cells removedby brief (3-5 min) trypsin
treatment. The number of adherent cells was
determined andtheTcell composition evaluated by flow cytometry. The degree of enrichment for
'y6Tcells after adherence tofibroblast
monolay-ers was determined by the number of
'y
T cells presentinthe inputand adherent cells. The pro-portionof mucosal V1 + y6Tcells abletoadhereto fibroblasts was determined from the number ofV61+ cells present in the input and adherent cell populations.
RESULTS
Insitu hybridization wasusedinitially to visual-ize and localize TCR-,y5+ cells infrozen sections of normal and diseased colonic mucosa.
Com-pared with sections of normal colonic mucosa,
sections ofdiseased tissue (CD) contained many
L. D. McVay etal.: y6 TCellsandInflammatoryBowel Disease
the muscularis mucosa and serosa (data not
shown). This distributionwasconsistent withthe
transmural pathology of CD. Inorderto analyze the distribution of -yS T cells in the normal and inflamed colon in more detail, flow cytometry
wasused to analyze cells directly after their
iso-lation from the colonicmucosa.
Changes in the Number, Distribution, and
Composition of Isolated Mucosal TCR-y6+
TCells in Noninvolved and Involved
Tissue from the SamePatient
Three-color flowcytometrywasusedtocompare
thenumber, distribution, and compositionof -y8
TcellsintheIE spaceandLPof the noninvolved
and involvedmucosa of individualIBD patients.
This approach was preferred to
immunohisto-chemical detection of T cells in intestinal tissue since this technique is more tedious and labor
intensive, it is difficult to perform, and it is a
relativelyinsensitivemeansof detectingantigens
such as the y6 TCR that are expressed at low
levels (28,29). For our study, surgically
resec-tioned segments of colon from a total of seven
patientswithcolonic CDorUCwereused for this
study. It should be noted that in contrast to the smallbowel which is rich in IELs, the colon has
relatively few IELs. Peripheral blood samples
from these patients were analyzed inparallel.
The cytofluorographs ofthe analysis of one
UC patient sample (Fig. LA) and the compiled analysis of allseven IBDpatients (Fig. 1B)
iden-tifydifferences in the distribution, number, and
composition of y6 T cells inthe blood and
intes-tine. In the blood, -y5 T cells comprised a small
proportion of T cells (3-4%), of which >95%
were CD4-, CD8- (data not shown) and
ex-pressedaTCRencodedbythe Vy9 andV52gene segments (Fig. iB). By contrast, inthecolon, ya
T cellsmade upalarger proportion of theTcells
presentinthe colonicmucosa,particularlyinthe
IE space (20-30%). Less than 10% of the IEL
population were CD19+, demonstrating that the
IELpreparationscontainedlow levels of
contam-inating LP lymphocytes. The 10-20% of
CD3-cellspresent inthe IEL preparationswereCD45
(datanotshown), whichis consistent with them
being contaminating epithelial cells.
In the involved mucosa of all of the IBD
samples analyzed, ySTcells comprisedamajorT
cell population in the LP of CD (mean, 43%;
range, 23-63%) and UC (mean, 64%; range,
45-83%) patients. Assumingthat the mucosalT
cell isolation procedure did not result in the
se-lective loss of ySTcells (seeMaterials and
Meth-ods), it was possible to determine the relative
number of y8Tcellspresentinthe involvedand
noninvolved mucosa. The averageyieldof T cells
(CD3+) recovered from the LP was 5.9 ± 0.4 X
1o6cells/g fromthe noninvolved colonic mucosa and 2.4 ± 0.6 X 107 cells/g from the involved mucosa, which is comparable to the values
re-ported previously (20,21). The proportion of
TCR-af3+ andTCR-,y8+ cells inthe noninvolved mucosa was 74% ± 10% and 26% ± 9%,
re-spectively (data not shown). Thus, noninvolved
colonicmucosasamples containedanaverage of
1.6 ± 0.3 X 106 -y Tcells/g tissue. Since in the
involved mucosa TCR-af3 and TCR-,yB repre-sented 54% ± 15% and 46% ± 22%, respec-tively, this tissue contained on average 6.9 ± 0.3 X 106 y6 cells/g tissue, representing an al-most 5-fold increaseover thatfound inthe
nor-mal mucosa.
Furthercharacterization of the mucosal y6T
cells in the normal and inflamed colon using
TCR-V,y/8-(J6)-specific antibodies revealed some interesting differences in the y6 T cell subsets present (Fig. iB). In the noninvolved colonic mucosa, V81+ ySTcells comprisedamajor
sub-set (43-52%) of IE 'y8 T cells. Of these cells,
V61-J8I-encoded TCR-8 chains predominated (63-76% of V81+ cells). By contrast, in the LP, V51+ y8T cells represented a minor subset (6-10%) of y8Tcells; as seen among IEL, VI+-J6I
wasthepredominant (43-58%)TCR-6 chain
ex-pressed by these cells. Thus, VSI-JSI-encoded receptorspredominate amongV61+ yS T cells in both the IE space and LP of the normal colonic
mucosa. Vy9/VS2+
y6
T cells that make up the largest-y5Tcell subsetinthe blood (Fig. IB) hadadistributioninthe normal colonic mucosa that
wasreciprocaltothat ofthe V61 subset. Whereas they representeda minor subset (13%) of
TCR-y75+
IEL, they were the majory6
T cell subset in theLP (67-79% ofTCR--y6+ cells).Incomparing they6Tcell subset distribution
in the involved and noninvolved mucosa, the
most striking difference was the increase in the proportion of VS1+ -y6 cells in the LP of the involved mucosa.Inboth theUCand CD patient groups, VS1+ cells represented the major (57-76%) population of yS T cells, of which most
(76-81%) expressed a V8I-J8I-encoded TCR.
By contrast, there was no significant difference
inthe proportion ofV 1-bearing y5Tcellsinthe
A
Non-Involved
I
Involved
IEL
0
7
.1 O l 1 l2 103 4
B
%CD3+ Cells %y8+Cells %V61+Cells
Patient
T cell source %CDl9+ 'l'+ V-9+/V82+ V81+ -J81+ CD8+
CD8-Colonic CD(n=3)
Blood ND 3 ± 2 78±8 10±8 ND ND ND
EEL N 5 4 21±10 13±7 43±13 76±12 75±6 21±4
I 10±3 28±13 27±12 67±21 74±11 33±4 68±7
LP N ND 5±4 67±11 10±5 58±8 42±6 63± 7
I ND 43 ±20 51±14 76±13 81±13 15± 3 86±5
TotalUC(n=4)
Blood ND 4 ±2 83±11 12 6 ND ND ND
EL N 4±3 22±9 13±10 52±14 63±19 83±5 13±4
I 11±4 26±8 18±9 61±23 60±14 40±2 62±4
LP N ND 15±7 79±16 6±4 43±10 ND ND
I ND 64±19 29±9 57±17 76±18 ND ND
IEL.inrepithelial lymphocytes;LP.laminapropria; N. non-involved colonic mucosa; I. involved colonic mucosa;
ND,notdetermined.
FIG. 1. Distribution of
'y6
Tcells inthenormalandinflamed intestinalmucosa(A) Representative analysis of y6Tcells present within the intraepithelialspace (IEL) and laminapropria (LP) of the colonic mucosa from apatient with total colitis. (B) CD8 expression andTCR-Vy/8usagebyIE- and
LP-y8
Tcells inthe blood, noninvolved (N) and involved (I) intestinalmucosa ofpatients withcolonic CD and totalcolitis. ND, not determined.
the intestinal mucosa ofIBD patients is
charac-terized, therefore, by changesinthe distribution of y6Tcellpopulations andby the accumulation of VS1+ cells in the LP. Although we do not
know if thisincrease in y8Tcells and V61+ cells represents expansion of resident mucosal cells or
cells infiltrating from the blood, differences in
the phenotype of V61+ cells in the LP (and IE
spaces) of normal colonic mucosa (majority
CD8+) versus involved mucosa (majority CD8 )
suggests they are mainlyofperipheral origin.
Bias inExpression of V81-encoded
Receptors by yST Cells in Surgical
Resected Tissue Specimens of IBD
Patients as Demonstrated by
RT-PCRAnalysis
Repertoireanalysis usingantibodies and flow cy-tometry is limited by the size of mucosal tissue samples, the number of cells that can be
recov-ered fromeachsample, andbytheavailability of TCR-V(J)-specific antibodies. Also, since the
LP
.
..4
I-
4 Iw..1-l-
.1
58
.
.....:.
F..-:
0
L. D. McVay etal.: ySTCells andInflammatoryBowelDisease
ILEUM TERMINAL
VIl- J3J2 33 1123334
L.JLJW-i-
L.JL.AJ"L-NL-1 NL-2 NL-3
UC-I
UC-2
UC-3
UC-4
UC-5
uc-8
UC-9
uc-10
UC-12
UC-15
ASCENDING TRANSVERSE DESCENDING
COLON COLON COLON
Jt32J2 4 312J3 4, JilJ2 34
L JJL- _JL-JLJi L-LMJ
CD-1 CD-2
CD-4
CD-S
CD-7
CD- II CD-12
DIV-1
DIV-2
DIV-3
not d)NA
FIG. 2. RT-PCRanalysis ofV61-J6 TCR expressioninsurgical samples ofnormal,noninvolved, and
inflamed, involved intestine ofindividual patients withCD,UC, diverticulitis (DIV),ornormal (NL)
mucosafromvarious locations along thelength oftheintestine
Mucosal cDNAsampleswere amplifiedfirst with primersspecific for V81 and CS andtheamplified products were gelpurified then reamplifiedin asecondPCRreaction withthe sameV61 primer used for the first amplification
and witha reverseprimerspecific foreachof the four J6 genes. V51-J5PCRproducts wereelectrophoresed
through 2% agarose gels, blotted, andprobedwithaninternal 32P-labeledV81-specificoligonucleotide probe. Autoradiography wascarried out for 24 hr. Asterisksindicate V6-J$ profile ofnoninvolvedcolonicmucosa.
V61-J61-specific antibody may have some
reac-tivity with J6-encoded receptors other than J61 (30), it was important to corroborate the results obtained by flow cytometry using another
inde-pendent approach. A sensitive RT-PCR based
method thatwehavepreviouslyused for
analyz-ing TCR-,y5 repertoire analysis in human fetal tissues (24,25) was used for this purpose. The
assay uses oligonucleotide primers specific for
V61, V62,orV83,incombinationwithaCSgene
segmentprimertoamplifycDNAgeneratedfrom
mRNAsamples usingreversetranscriptase.After
gelelectrophoresis the products of amplification
were excised from the gel, and equivalent
amounts usedas a template for a second
ampli-fication using thesame forward VSprimeranda
reverse-primer specific for each of the four
hu-manJ8 genes. In establishingthisassay,
individ-ualV81-JT1-4, V82-J51-4, and V63-J61-4
recep-tor cDNA clones were used to determine the
optimum conditions for the proportional
ampli-fication ofV5-J5genes; therewas no
cross-prim-ing. The products of this second amplification
werethenelectrophoresedon 1.8%agarosegels,
Southern blotted, and hybridized with [y32p]_
ATP end-labeled V51, V62, or V83
oligonucleo-tide probes containing sequence internal to the
VS primers used for PCR.
This assay wasfirst used to analyze samples
of intestine from noninvolvedandinvolved
seg-SIGHOID/RECTUMX
JL J2J3 .
PERIPHERAL,
BIAWD J3 J2 3 4
L-JLJL-JL-i
INVOLVED NON-INVOLVED PERIPHERALBLOOD
V62- Jl J2 J334 JJ2334 J1 J2 J3 J4
t .,_ I__ LI L-J XtJ*.I
NL-lI NL-2
NL-3
UC-2
UC-4
UC-IO
UC-11
UC-'1
CD-I
CD-2
CD-4
CD-8
CD-7
CD-6
INVOLVED
V63- J3 J2 J3 J4
L.JLJiL- L-J
NL-I
NL-2
NL-3
UC4
-UC-II
-UC-12
-UC-IO
CD-I
CD-5
CD-12
CD-8
CD-7
CD-6
NON-INVOLVED PERIPHERALBLOOD
JI J2L_JL__J.J3 J4 J3 J2 J3 J4
I L- _JL_J LI.JLIJ
FIG. 3. Profile ofV62-J6 andV63-J6TCRexpression in surgical samples ofnoninvolved andinvolved intestine of individualpatients with CD, UC, or normal (NL) mucosa from colon cancer patients RT-PCR was carried out as described in Fig. 2-using 32P-labeled oligonucleotide probes complimentary toV62 or V63. Autoradiography wascarried out for 24 hr.
ments of tissue obtained from 20 patients with
IBD for Va-Jl receptor expression. Ten patients
diagnosed withUC (1-16inTable 1), sevenwith
CD (17-28 in Table 1), and three patients with
diverticular disease (diverticulitis, 29-31 in
Table 1) were used as a source of involved and noninvolved mucosalmRNA. In addition, tissue
from the normalmarginsofcolonicmucosafrom three colon cancer patients (32-34 in Table 1) were also used as noninvolved, "normal" con-trols. The results of this analysis shown inFig. 2
identify changes inthepatternofVS1
-J6
expres-sion in the involved mucosa compared to the noni-nvolved mucosa and blood of IBD patients.TheV8-J5profile of the noninvolved colonic mu-cosa (* profiles in Fig. 2) was characterized by
expression of receptors encoded by V81-J61, 2,
and 3, and by very low levels or an absence of
V61-J84-encoded receptor chains. This profile
was distinct from that of theperipheral bloodin
whichallfourJ6-genesegmentswereutilizedby
V61-encoded receptors. By contrast, a single Va1-encoded receptor predominated in the
in-volved colonic mucosa. Regardless of whether the involved intestinal tissue was derived from
patients with UC, CD, or diverticulitis, V61-J61
receptor expression predominatedand itwasthe
only V81-encoded receptor detected in 13/17 IBD patients and 3/3 patients with diverticular disease. Importantly, thisfinding agreeswith the results obtained by flow cytometry in which
V51-J61 cells were the major
V61'
yb T cellpopulation in the involved colonic mucosa
(Fig. 1), and it also demonstrates the utility of this procedure for TCR--ya repertoire analysis.
DNA sequence analysis of randomly selected
V1-Ca cDNAs from involved and noninvolved
mucosal mRNAsamples from a patient with UC did not identify any V51 receptors that were uniquetothe inflamed colonic mucosa (datanot
shown). All of the Val-TCR-cDNAs encoded
functional, in-frame receptors consistent with the receptor cDNAs detected by RT-PCRanalysis being derived from productively rearranged
genes. The V51-Ca cDNAs from noninvolved
mu-L. D. McVayetal.: ySTCellsandInflammatory Bowel Disease
INVOLVED
V61- JI J2 J3 J4
L-)LJ j
NON-INVOLVED
JI J2 J3 J4
LA *_jI DURATION
15y
12y
1wk
5y
V52-J41 waspredominantor wastheonly
recep-torexpressed. Likewise,inraresamples (i.e.,
UC-10) a bias in the V63-J8 expression was also evident. Taken together, our analysis of the
TCR-V6 repertoire in the colonic mucosa
dem-onstratesthata single VSreceptor, V 1-J$1,
pre-dominates in the inflamed mucosa of patients
with CD and UC.
5-6y
13y
5y
2v
FIG. 4. Profile ofVei-Je TCRexpression in
mucosalbiopsysamples of noninvolved and
involved intestine ofindividualpatientswith colonicUC
PCRwas carried out asdescribed in Fig. 2.
Autora-diography
was carried outfor24 hr.cosa utilized the J61 gene segment, which is consistent with the results fromboth flow cyto-metricandRT-PCRanalyses.Inaddition,asingle
D-gene segment, D63, was exclusively used by
all of the VSI-receptor chains expressed in the noninvolved (33/33) and involved (22/22) mu-cosa.
Lack ofBias inthe Expressionof V62- and
Ve3-encoded Receptorsin Surgical
Resected Tissue Specimens of
Patients with IBD
Wewereinterestedinknowing whether the V82
and V63 receptors, which are similar to V81-encoded receptors, were also biased in JS gene
usage. Figure 3 shows the V82/V83 repertoire
analysis of 10 IBD patients representative of a
total of20patient samples analyzed. Inthe
nor-mal intestine, V62 receptors primarily utilized J61 and J63 whereas V83 receptors utilized all fourJ6 genesegments.Thisprofile ofV62-J6 and V63-J6 expressiondidnotchange significantly in the noninvolved intestine or in tissue samples
obtained from different regions of the inflamed intestine (data not shown). In2/16 CD samples
and 3/35 UC samplesfromtheinflamedmucosa,
Bias inExpression ofV61-encoded
Receptors by yST CellsinBiopsy
Tissue Specimens
TheRT-PCRassaywasnextappliedtotheanalysis
of colonic mucosal tissuebiopsy samples to
deter-mine ifchanges in yST cell repertoire detected in patients with severe disease were present in
pa-tients with newly diagnosed and/or less severe
forms ofIBD (Table 2).Becauseofthe small size of the mucosal tissuesamples procured by
sigmoidos-copyandcolonoscopy, itisnotnormallypossibleto
isolatesufficient numbers ofcellsfor
fluorescence-activatedcellsorting (FACS)analysis.Thesamples
obtained from eight UC patients (Table 2) were
therefore processed forRNAisolationandRT-PCR
analysis of TCR-V6-JT expression. The results of this analysis shown in Fig. 4 clearly identify V61-J61 receptorexpression asbeing the predominant
receptorexpressed inallof thepatientsamples.In
five of eight patient samples itwastheonlyV61-J6
TCR-8chainexpressed. Interestingly, this receptor was also predominant in the mucosa of a newly
diagnosed UC patient who had symptoms of
dis-easeforless than 2weeks. Theexpressionofother
V81-Jo-encodedreceptorsin noninvolved samples strongly suggests that the predominance of
V81-J61 receptor expression in the involved mucosa
was notdueto a PCR-relatedartifact.
In summary, our cellular and molecular
analyses of y6 T cells in the colonic mucosa of
patients with IBD has shown that these cells
accumulate in large numbers at the site of
in-flammation and tissue injury and canbe
distin-guishedbyexpression ofaTCR-8chainencoded
by V61-J61 gene segments. These changes were
apparent in all the IBD patients we analyzed,
irrespective of the form orstage and severity of
disease. To begin to understand the significance ofthese changes in the mucosal y8 T cell
reper-toire for the disease process inIBD, subsequent
experiments were designedtoanalyzethe effec-torfunction of mucosaly5T cells.
B-I
B-2
B-3
B4
B-5
B-6
B-7
B-8
A l 5B | ihi i
,... ..,.~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~...
, 1_
M_
~
~
~
~
~
~
~
~
M
FIG.5.In situ hybridizationanalysis ofIFN-ygene expressionin sections ofnoninvolved. r Fed
areasof colon from CD patient, CD-i
Lo
_;1L5...
^_S
7:t.;..':'4'~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.
.
....
II s;4sAl~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~31r -5-':::.:
w --~~~~d..._
(A). 5Nonsinvolved issu sen Rn probe. (B Noninvole d tiss, aontinsenseiRnA probe(ninvolved tiss.e,
sense RNAprobe. (D-F) Involvedtissue, anti-sense RNAprobe. L, lumen;M, mucosa; S, submucosa; By, blood
L. D. McVayetal.: ySTCells andInflammatoryBowel Disease
TABLE 3. IFN-,yproductionbylaminapropriaTcells in the normal and diseased colon
U/ml TCell
Populationa %mRNA+ Cellsb Media anti-CD3
CDPatient #1 (Ileocolonic)
Total N 12 ±4 <5 39 ±8
I 39 ± 5 5 99± 10
a,+ T cells N 10±2 10 ±2 38 10
I 46± 10 9±3 53± 14
-y5+ T cells
(9%VS1+) N 16±4 7 ± 6 19 ±2
(35% V$1+) I 39 ± 5 12 ±4 109± 5
CDPatient#2(Ileocolonic)
Total N 15 ±3 8 ±9 13 ±2
I 61 ±6 7 ± 7 78 ±9
a,l3Tcells N 11±5 6±6 23±5
I 56 ±8 13 11 57 ±8
,yS+ Tcells
(12% V81+) N 17 9 11 8 18± 13
(53% V61+) I 38 7 24 10 165± 11
CDPatient#3 (Colonic CD)
Total N 11 6 <5 59 ± 17
I 35 10 43 19 78± 21
ap+
Tcells N 29±5 18 9 39± 6I 39 7 33 11 54± 10
,yb+ Tcells
(15%V61+) N 9±3 12±8 35± 12
(43% V$1+) I 66 8 34 15 177 ± 21
aNonfractionated (total)Tcells andpurified populations of
af3+
and-yS+ cells were isolated from segments of noninvolved (N)and involved(I) colonicmucosa.Frequency ofV8I-expressing cellsin y6Tcell preparations is shown in parentheses.
bFrequencyof cellsthat afterhybridizationinsituwith anti-sense probe were overlayed with more than 5 grains.
Mucosal ySiT Cells Area Major Sourceof IFN-,yProduction in IBD
Since IFN-y production appearstobe an
impor-tant mediator of the inflammatory process and tissue injury in patients with IBD (31) and is involvedinthepathogenesis of intestinal
inflam-mation inanimal models ofIBD (32), we inves-tigated whether
y8
T cells in the intestinal mu-cosaof patients withIBD were a sourceof IFN-,y.In situ hybridization analysis of
IFN-'y
mRNAexpression in tissue sections from involved or
normal intestinal mucosaltissue (Fig. 5) showed that the
IFN-'y
genewasconstitutively expressedatlowlevelsby cellsinthenormal, noninvolved,
mucosa. By contrast, in sections ofinvolved co-lonic tissue froma CD patient (CD-1), the num-berof IFN-'y-mRNA+ cells wasgreatly increased (Fig. SD). Nohybridization signal was detected in
sectionsprobed withacontrol senseprobe (Fig.5A
andC). The distribution of IFN-,ymRNA+ cells in
diseased mucosa (Fig. SD) was similar to that of
TCR-S+ mRNA-expressing cells (data not shown), suggestingT (y6) cells as alikely source of IFN-,y production. Inaddition, the identification of IFN-,y
rnRNA+ cells in blood vessels in the submucosa (Fig. SE) and muscle (Fig. SF) was consistent with
A
r-
F-c_ 0
._
0
0
B
0 0
-._
0
E
0
._ 3
Po
Colon Skin Lung
Fibroblast Cell Lines
N I N I N I N I
CD7-ML CD8-ML CD9-ML CD10-ML
FIG. 6. Enrichment ofy6Tcells amongperipheral blood (PB) and colonic mucosal (ML)Tcellsby adherence tofibroblast cell lines
(A) Selectiveadherenceofy6Tcellstofibroblast celllinesof intestinalorigin. Tcells isolated from the blood of normal healthy donors (N-PB) andCDpatients (CD-PB) and from segments of inflamed(involved) colon fromCDpatients
(CD-ML) wereco-cultured with different established fibroblast cell lines for30minat370C.The fold-enrichment ofys
Tcells among theadherentcells was determinedby comparing the number ofySTcellsintheinputandadherent frac-tion, the values for whichwereobtainedbystaining with anti-TCR antibodies and flow cytometry. (B) Preferential ad-herence of mucosal 'ySTcellstocolonic fibroblasts. ColonicTcellsisolated from segments of involved (I)and nonin-volved (N) colon from thesame CDpatient werecultured withthe colonic fibroblast cellline,CCD-18Co,for30min. Fold-enrichment of adherenta(3and ySTcellswasdetermined byflow cytometry.
the transmural distribution of
TCR-8-mRNA'
cells (datanotshown).To identify the cellular source(s) of IFN-,y production in the colonic mucosa, IFN-y-gene expressionand protein secretionwasanalyzedin
isolated populations of
y6
and aj3 LP T cells. These T cell populations were isolated from the mononuclear cell fraction of noninvolved and involved mucosa from three patients with CD using immunomagnetic selection. The isolated populationswereroutinely >95% CD3+and the degree of contamination of they8
fraction byaI3+ cells and the af3 fraction by y6+ cells was routinely <10% (data not shown). As seen in ourpreviousflow cytometricanalyses of mucosal
Tcells (Fig. IA), VS1 + cells were the major y6T
cell subset (67 ± 8%) in the involved mucosa compared with the noninvolved mucosa (12 ±
2%).Theincrease inthe number ofTcellsinthe
LPof involvedmucosa(seeabove)wasparalleled by an increase in the frequency of cells that
constitutively expressed IFN-,y-mRNA (Table 3). Thiswasparticularly apparent for
y6
Tcells that contained a higher proportion of mRNA+ cells than af3 T cells. The analysis ofIFN-'y
secretionwas consistent with the mRNA analysis and
demonstrated that
y8
T cells, including V81 +cells, were a major source of this
proinflamma-tory cytokine in the involved mucosal CD
sam-ples analyzed.
Selective Interaction of 'y6 T Cells with Colonic Fibroblasts
Inviewofthe accumulation of y6Tcells, partic-ularly VS1+ cells, in the LP of the involved
in-testinal mucosa of CD patients and results from previous studies demonstrating that y8 T cells
can interact with stromal cells in other tissues
suchasskin (9,33),wedetermined if mucosal y6
Tcells, particularly VS1+cells, from patients with
IBD could adhere to and interact with colonic stromal cells, and fibroblasts in particular. To accomplish thiswe took advantage of the
avail-ability of established and well-characterized fi-broblast cell lines of different mucosaltissue
or-igin and the y8 T cell-skin fibroblast adherence assaydescribedby White and co-workers (9).To
date, the possibility that
y6
T cells can interact with colonic stromal cells suchas fibroblasts and how this interaction might be affected duringIBDhasnotbeeninvestigated. Bulkpopulations of colonicT cells isolated from segments of
nor-mal and involved tissue of individual IBD
pa-tients were briefly incubated with confluent
L. D. McVay et al.: ya TCells and InflammatoryBowel Disease
B
U,
.-A (U
0
WI-U,
.0
80
60
40
-20
0-CD7-ML CD8-ML CD9-ML CD10-ML CD-11-ML
FIG. 7. Predominance of fibroblast-adherent VS1I 'yi Tcells in the inflamed colonic mucosa
Colonic Tcells isolated from segments of involved and noninvolved colon of CD patientswereco-cultured for 30 minwith the established colonic fibroblast cell lines CCD-18Co (A) ora primary colonicfibroblast cell line ob-tainied frorn involved mucosa of CD Patient 11 (B). Theproportion ofV61+ fibroblast-adhering cells was deter-mined from the nluLmber of V6l+ cells in the inpLutand adherent cell populations by flow cytometry.
and phenotype of adherenit and nonadherent
cells determiined by flowcytometry.
As seein in Fig. 6A, mucosal y6 T cells from
one of two patienits with CD (CD8) were
consid-erablyenriched for after co-culture with colonic fibroblasts. Whereas co-culture with skin- or
luIng-derived libroblasts produced between 1.5-and 3.5-fold enlrichment of -y6 T cells, culture
with the colonic fibroblast cell line CCD-18Co resulted in an almost 10-fold enrichment of y6T
cells. A similar level of y6 Tcell enrichment was
also seenusinganothercolonicfibroblast cell line
(CCD- 12Con) (data not shown), excluding the
possibility that the interaction with the CCD-18Co cell line was due tosomepeculiarproperty
of this cell line. Analysis of nonadherent T cells from cultures with colonic fibroblasts showed a
reciprocal decrease in the y6 T cells (data not
shown), excluding the trivial explanation that theirenrichmeintwas dueto preferential survival duringorafter culture.Thespecificityof mucosal
-y T cell-fibroblast interaction was also
demon-strated by the failure to see a similar enrichment
of y6 T cells tisinlg peripheral blood from two of
thesame CDpatients, orusingfibroblasts derived
from sources other than the colon. Enrichment
of o43 T cells am-ong adherent cells after
co-cul-ture with colonic fibroblasts, or fibroblasts of other tisstic origin (data not shown), was 2-lold or less (Fig. 6B), which suggests that increased
adhesiveness of -y T cells reflects an inherent
p)roperty of these cells rather than a function of
the fibroblastmonolayer. Slightly higherlevelsof
-y6 T cell enrichment were seen usinlg
lympho-cytesfrom the involvedversus noninvolved
mu-cosa with three of the four patienits with CD
(Fig. 6B).
Analysis of the fibroblast-adherent -y5Tcells
showed that the majority of those isolated
from the noninvolved (75%0/ + 8%) and
in-volved (84%±+ 9%) mucosa of four patients
with CD (#7-10; Fig. 6B) were V61+. By
com-paring thenumberofV61+ cellsinthe input and adherent cell populations it was possible to
de-termine what proportion of these cells in the
noninvolvedversusinvolvedmucosaofthesame
patient were fibroblast-adherent (Fig. 7A).
Whereas in the noninvolved mucosa
fibroblast-adherent V61 cells represented <40% (mean of 310% with a range of 24% to 390/o) they
repre-senited almost 80% (mnean of 78% with a range
of 63% to 92%) of V61+ cells in the involved
mucosa. Thus, the accumulation of V61+ -y T
cells in theinflamedmucosaofpatientswithIBD
wasassociated withan increase intheirabilityto
interactand adheretocolonicfibroblasts. Usinga
primary colonic fibroblast cell line derived from mucosal tissue sample from a patient with pri-mary colonic CD (Fig. 7B), we have reproduced
the resLilts obtained using established colonic fi-A
100
80
60
40
20
cn
0
-W
U,
0
O Noninvolved * Involved
CD12-ML
nn
broblast cell lines. In particular, using either do-nor-matched or allogeneic LP lymphocytes from noninvolved or involved mucosa as input cells, a higher proportion of V81+ cells from the in-volved mucosa (88% ± 6%) were fibroblast-adherent compared with those from the nonin-volved mucosa (47% ±- 5%).
DISCUSSION
We have compared the phenotypic and func-tional characteristics of ySTcells within the nor-mal and inflamed colonic mucosa of a large number of patients with IBD. Our results
dem-onstrate that the chronic inflammatory immune responsethat occurs inthecolon of IBD patients is associated with distinct changes in the
num-ber, distribution, composition, and function of mucosal y6 T cells. The size of the y6 T cell
population in the diseased mucosa was signifi-cantly increased and could be attributed to an
increase in V81+ y8 T cells, among which, cells thatexpressedaTCR8-chain encodedbyV1-J81 gene segments predominated. These changes in
mucosal ySTcells were notrestrictedto individ-ual patientsor aparticulargroupof patientswith
IBDbutwerepresentinthe majorityof UC, CD, and diverticulitis patient samples that we
ana-lyzed. The predominance of V81-J81
receptor-bearingcells was seen inpatientswith advanced forms of disease who required surgery and in
biopsy samples obtained from patients with less
severe forms of IBD, as well as in apatient that had clinical signs of disease for less than2weeks. AmongTcells presentintheinflamedmucosaof patients with CD, y6Tcells were a major source
of theproinflammatory cytokineIFN-,yandwere
enriched for V81+ yBTcells that couldselectively
interactwithcolonicfibroblasts. Althoughit was notpossible to determine ifthese changes inthe properties of mucosal
y6
cells were primary ab-normalities of these disorders, they areconsis-tent with, and suggest that they contribute to,
thepathophysiology of IBD.
The results of our studies contrast with those obtained previously from the immunohisto-chemical analysis of y8 T cells in colonic tissue sections obtained from IBD patients in which a
decrease (17) ornochange (16)inthenumber of
TCR-6+
cells was reported. A likely explanationfor the apparent discrepancy between thesetwo
studies and our own is the different techniques used. The immunohistochemical detection of T
cells inintestinal tissueisdifficult to perform and is a relatively insensitive means of detecting an-tigens such as the yS TCR that are expressed at
low levels (28,29). In addition, since therelative number ofy8Tcells within the intestinal mucosa has been shown to vary greatly, not only be-tween different sections of the same tissue but also between sections of the samesiteoriginating from different subjects (29), estimates of their relativenumberbased upon this technique alone
are unlikely to be accurate. Finally, since the
degree ofinflammatory activity may also
signif-icantly influence the distribution of y6T cells in
the diseased colon, variation among individual patients withIBD may also accountfor some of the contrasting results obtained fromthe
differ-ent studies.
The increase in
V61-expressing y8
T cells intheLPof the inflamed colonofIBDpatients may represent the expansion ofresident and/or infil-trating cells fromthe blood. Since there are
cur-rently no phenotypic markers, including CD8, that can unambiguously discriminate between intestinal and blood-derived V1
y8
T cells, itis not possible to definitivelyidentify the origin of V61+ cells within the LP of patients with IBD.However, several pieces of evidence suggest that
alarge number of these cellsarederived from the
peripheral blood. The phenotype of LP V61+
(CD8 ) cells in IBD patients is similar to that of those normally found in theblood (34) and
re-sembles the phenotype of blood-derived
y$
Tcells that possess transendothelial migratory properties andaccumulate within tissuesatsites of inflammation (10). Interestingly, Chowers andcolleagues (3 5) haverecentlyshownthat,on
the basis of the presence of unique junctional
sequences of TCRs expressed by intestinal
y6
Tcells, it is possible to distinguish between V81+
cells normally resident in the blood and intes-tine.
Irrespective of the origin of the V81+
-y6
Tcells that accumulate in the colonic mucosa of
IBDpatients, animportantquestionthatremains tobeaddressedis thenature of thestimulus that drives their expansion. The oligoclonal nature
and limited structural diversity of the intestinal
VMI
repertoire in the normal intestine (35) sug-gestthat thearrayofligands recognized bythese receptors is limited. There are three possiblesources ofthese ligandsinthe inflamed colon of
IBD patients. The first is that they are enteric
(bacterial and food) antigens that mucosal V81+
y8
Tcells could beexposedtoas aresult of themL. D. McVayet al.: y6TCellsandInflammatory Bowel Disease
intestinal wall, which is disrupted as part ofthe
disease process in IBD. The recent observation
that E. coli and P.
aeruginosa,
which normally inhabit the gastrointestinal tract, can stimulate V51-bearing peripheral blood y6 T cells (36) isconsistentwith thispossibility. Furthermore, the
finding that gut T cells from patients with IBD
proliferate vigorously to preparations of their ownflora (37) suggests that abreakdown inthe mechanismsthatnormally operateto maintain T
cellhyporesponsiveness ("oral tolerance")to en-teric antigens is an important pathogenetic mechanism of IBD. A second possibility is that the antigens are endogenously derived and are
expressed by stressed ordamaged epithelial cells. The selective accumulation of V81+ y6 Tcells in
the IE spaces of the intestine has long been
in-terpreted as evidence for an immune surveil-lance role for thesecellsinwhich they recognize and react with a common antigen, such as heat shock proteins (HSP), expressed by epithelial cellsas aresult ofinfection, damage, or transfor-mation (38). To date, however, recognition of
self-antigens such as HSP by y6 T cells has only been demonstrated for cells that utilize a
recep-torencoded by the V82 and Vy9 gene segments (39,40). Extensive epithelial cell damage could, however, enable IEL such asVMI+ cells to"leak" intothe LP, thus contributing to the increase in the number of V81+ cells seen inthe LP of IBD patients. The third possibility is that the
activa-tionofV61-expressing ySTcells is antigen-inde-pendent and does not involve the TCR/CD3
complex. The existence of alternative, antigen-independent pathways of mucosal T cell
activa-tion is suggested by the finding that LP T cells, unlikeperipheral bloodTcells, from normal and
IBD patients exhibit vigorous proliferative re-sponses via the CD2/CD28 signaling pathway (41-45).The presenceinthe inflamed mucosa of
large numbers of activated T cells that produce
cytokines (46-49)couldalso serve to activate,in a"bystander" manner, V61+cells (50), as well as
tomaintain apopulation of chronically activated
T cells within the mucosa.
The
y8
T cells that accumulate in the in-flamed mucosa of patients with CD possess two functional characteristics that are relevant to thepathophysiologyof thisIBD. The first is that they
are a majorsource oftheproinflammatory cyto-kine IFN-y, which has long been shown to me-diate epithelial cell injury (51). Although the
constitutive production of
IFN-'y
by colonic lym-phocytes from IBDpatients has been previously reported (44,49,51), our analysis identifies yS Tcells, including V61 + cells, as a source of this
cytokine in situ in the inflamed mucosa, and
suggests a meansby which they might contribute
to the chronic inflammatory immune reaction andtissueinjuryin CD.Thesecondcharacteristic
isthe selective adherenceand interaction of
mu-cosal y6 T cells, particularly VS1+ cells, with
in-testinal-derived fibroblasts. Although uncon-trolled fibroblastproliferation, excessivecollagen secretion,and fibrosis cause severe tissuedamage
and are important complications of CD (52,53),
theprocesses regulating normal fibroblast
activ-ity arepoorly understood. Onewayinwhichthe
activity of fibroblasts and other stromal cells could be regulated and coordinated is through direct interactionswith othercells, such asVS1+
y6 Tcells. Theproduction by -yBT cells of tumor necrosis factora (TNF-a), ILI-f, and
transform-ing growth factor ,B (TGF-,B) (54) that can pro-mote fibroblast growth and collagen synthesis (55), and the observation that
epithelial-associ-ated y8 T cells in the skin can influence the
activity of keratinocytes by the production of
keratinocyte cellgrowth factor (8) areall consis-tent with this hypothesis. This immunoregula-torypropertyofySTcellsisfurther substantiated by the results fromavarietyof animal models of infectious disease in which the ability of -yS T cells toinfluence the activity of other cell pop-ulations and the outcome of inflammatory im-mune responses has been demonstrated (56-61). Since ithaspreviously been shown that fibroblasts can prolong the survival ofT cells in
culture after the removal of IL-2 and antigen (62), fibroblast adherence may also provide a
means of localizing or retaining activated, pe-ripheral blood-derived, V51+ cells in the in-flamed colon. Although we do not yet have an explanation for the differences in 'y$ T cell
sub-sets adhering to colonic fibroblasts, it could be related to quantitative and/or qualitative
changes in the expression of adhesion/integrin
molecules such as LFA-1 and VLA-5 by V81+
cells (9,33,63) and their reciprocal receptors,
ICAM-1 and fibronectin, respectively, by fibro-blasts (9,33). The preferential association of VS1+ y8Tcells withfibroblasts provides a possi-ble explanation for their accumulation in the lungs (64) and joints (65-69) insarcoidosis and rheumatoid arthritis patients, respectively. Inter-estingly, inflammatory disorders of the lung, joints,skin,andeyeareoften presentas
where, asa result of theirabilityto interactwith fibroblasts or other connective tissue elements,
they accumulate and can contribute to the
in-flammatory immune reaction and inflammation atthese sites.
Insummary, we have demonstrated that the chronic inflammatory reaction and tissue injury that occurs in the colon of patients with IBD is associated with distinct changes in the number, distribution, composition, and effector function of mucosal
'y
Tcells,particularlyV81-expressing cells. These findings suggest that these cellscon-tribute to at least some of the immunological and
pathological changes presentin the inflamed
mu-cosa of patients with IBD, andparticularly CD.
ACKNOWLEDGMENTS
We aregrateful to Ms. Linda Hurd for compiling
pertinentdataon IBDandnormal patientsinthis study. We thank William G. Crowley for expert technical assistance.This work wassupportedby grants from the American Cancer Society (JFRA 399) and the National Institutes of Health (Al 31972) toS.R.C. L.D.M. wassupported in part by
a National Institutes of Health Training Grant (5-T32-CA-09140-20) and a Crohn's and Colitis of America (CCFA) Fellowship.
REFERENCES
1. Strober W, James SP. (1986) The immuno-logical basis of inflammatory bowel disease.
J. Clin. Immunol. 6: 415-433.
2. Shanahan F, Targan S. (1989) In: Shaffer E, Thompson ABR, (ed). Modern Concepts in Gas-troenterology. Plenum, New York, pp. 291-310.
3. Strober W, Ehrhardt RO. (1993) Chronic
in-testinal inflammation: An unexpected out-come in cytokine or T cell receptor mutant mice. Cell 75: 203-205.
4. MacDonald TT, Spencer J. (1988) Evidence that activated mucosal T cells play a role in the pathogenesis of enterpathy in human smallintestine.J. Exp. Med. 167: 1341-1349.
5. MombaertsP,Mizoguchi E, Grusby MJ,etal.
(1993) Spontaneousdevelopment of inflam-matory bowel disease inT cell receptor mu-tant mice. Cell 75: 275-282.
6. Sadlack B, Merz H, Schorle H, et al. (1993) Ulcerative colitis-like disease inmice with a
disrupted interleukin 2 gene. Cell 75: 253-261.
7. Deusch K, Luling F, Reich K, etal. (1991) A major fraction of humanintraepithelial lym-phocytes simultaneously express the y6 T cell receptor, the CD8 accessory molecule
and preferentially uses the VS1 gene seg-ment. Eur. J. Immunol. 21: 1053-1059. 8. Boismenu R, Havran WL. (1994)
Modula-tion ofepithelial cell growth by intraepithe-lial y6 T cells. Science266: 1253-1255. 9. White B, Korn JH, Piela-Smith TH. (1994)
Preferential adherence of human y8, CD8+ andmemoryTcellstofibroblasts.J.Immunol. 152: 4912-4918.
10. Galea P, Brezinschek R, Lipsky PE, Oppen-heimer-Marks N. (1994) Phenotypic charac-terization ofCD4U/a TCR+ and y8 TCR+ T
cells with a transendothelial migratory
ca-pacity. J. Immunol. 153: 529-542.
11. Kagnoff MF. (1996) Mucosal immunology:
Newfrontiers. Immunol. Today 17: 57-59.
12. Halstensen TS, Scott H,BrandtzaegP. (1989)
Intraepithelial Tcells of the TCR
y6+
CD8and V61/J81+ phenotypes are increased in
coeliac disease. Scand. J. Immunol. 30: 665-672.
13. Spencer J, Isaacson PG, Diss TC, MacDonald
TT. (1989) Expression of disulfide-linked
and non-disulfide-linked forms of the T cell receptor y8 heterodimerinhumanintestinal intraepithelial lymphocytes.Eur. J. Immunol.
19: 1335-1338.
14. Spencer J, Isaacson PG, MacDonald TT, Thomas AJ, Walker-Smith JA. (1991) Gam-ma/delta Tcells andthe diagnosis of coeliac disease. Clin. Exp. Med. 85: 109-113.
15. Rust C, PenaS,KluinP, KoningF. (1990) y8
T cells in coeliac disease. Res. Immunol. 141:
668-671.
16. Trejdosiewicz LK, Calabrese A, Smart CJ, Oakes DJ, Howdle PD, Crabtree JE, Losowsky MS, Lancaster F, Boylston AW.
(1991)
y6
Tcell receptor-positive cells of the human gastrointestinal mucosa: Occurranceand Vregion gene expression inHeliobacter pylori-associated gastrtis, coeliac disease and
inflammatoryboweldisease. Immunology84:
440-444.
17. Fukashima K, Masuda T, Ohtani H, Sasaki I, Funayama Y, Matsuno S, Nagura H. (1991) Immunohistochemical characterization,
dis-tribution, andultrastructure oflymphocytes bearing T-cell receptor y/8in inflammatory
L. D. McVay et al.: y6TCells and Inflammatory Bowel Disease 201
18. Stallmach A, Schuppan D, Riese HH,
Mat-thes H, RieckenEO. (1992) Increased colla-gen type III synthesis by fibroblasts isolated from strictures of patients with Crohn's
dis-ease. Gastroenterology 102: 1920-1929. 19. Bull DM, Bookman MA. (1977) Isolation
and functional characterization of human intestinal mucosallymphoid cells. J. Clin. In-vest. 59: 966-974.
20. Bookman MA, Bull DM. (1979)
Character-istics of isolatedintestinal mucosalymphoid
cells in inflammatory bowel disease.
Gastro-enterology 77: 513-520.
21. Selby WS, Janossy G, Bofill M, Jewell DP. (1984) Intestinal lymphocyte
subpopula-tions in inflammatory bowel disease: An
analysis byimmunohistological and cell
iso-lation techniques. Gut 25: 32-40.
22. Carding SR, Kyes S,Jenkinson EJ, Kingston R, Bottomly K, Owen JJT, Hayday AC. (1990) Developmentally regulated fetal
thy-micandextrathymic T-cell receptor
y6
gene expression. Genes Dev. 4: 1304-1315. 23. Li B, Rossman MD, Imir T, Oner-EyubogluAF, Lee CW, Biancaniello R, Carding SR. (1996) Disease-specific changes in
-yS
T cellrepertoireandfunctioninpatientswith
pul-monarytuberculosis. J.Immunol. 157: 4222-4229.
24. McVay LD, Carding SR. (1996) Extrathymic
generation of
y8
T cells bearing invariant antigen receptors during fetal development.J. Immunol. 157: 2873-2882.
25. McVay LD, Carding SR, BottomlyK,Hayday
AC. (1991) Regulated expression and struc-ture ofTCR gamma delta transcripts in
hu-manthymic ontogeny. EMBOJ. 10: 83-91. 26. Buschle M, Campana D, Carding SR,
Rich-ards C, Hoffbrand AV, Brenner MK. (1993)
Interferonyinhibitsapoptoticdeathin B cell chronic lymphocytic leukemia. J. Exp. Med.
177: 213-218.
27. CardingSR, McNamara JG, Pan M, Bottomly
K. (1990) Characterization of
y6
Tcellclones isolated from human fetal liver andthymus.Eur. J. Immunol. 20: 1327-1335.
28. Borst J, van Dongen JJM, Bolhuis RLH, Pe-ters PJ, Hafler DA, deVries E, van de Griend
RJ. (1988) Distinct molecular forms of
hu-man Tcell receptor
y/6
detectedonviableT cells byamonoclonal antibody. J. Exp.Med.167: 1625-1644.
29. Vroom T, Scholte G, Ossendorp F, Borst J. (1991) Tissue distribution of human y5 T
cells: No evidence forgeneral epithelial tro-pism. J. Clin. Pathol. 44: 1012-1017.
30. Porcelli S, Brenner MB, Band H. (1991) Bi-ology of the human y6 T-cell receptor.
Im-munol. Rev. 120: 137-183.
31. Fiocchi C. (1994) Cytokines. In: Targan SR,
Shanahan F (eds). Inflammatory Bowel Dis-ease, from Bench to Bedside. Williams and
Wilkins, Baltimore, pp. 107-122.
32. Powrie F, Leach MW, Mauze S, Menon S,
CaddleLB, CoffmanRL. (1994) Inhibitionof
Thl responses preventsinflammatory bowel disease in scid mice reconstituted with
CD45RBhi CD4+ T cells. Immunity 1: 553-562.
33. NakajimaS, RoswitWT, LookDC, Holtzman
MJ. (1995) A hierarchy for integrin expres-sion and adhesiveness among T cell subsets thatislinkedtoTCR geneusageand
empha-sizes V81+ -y5 T cell adherence and tissue retention. J. Immunol. 155: 1117-1131. 34. GrohV, PorcelliS,FabbiM,Lanier LL,Picker
J, Anderson T, Warnke RA, Bhan AK,
Strominger JL, Brenner M. (1989) Human
lymphocytes bearing T cell receptor y/1 are
phenotypically diverse and evenly distrib-uted throughout the lymphoid system. J. Exp. Med. 169: 1277-1294.
35. Chowers Y, Holtmeier W, HarwoodJ,
Mor-zycka-Wroblewska E, Kagnoff MK. (1994)
The VS1 receptorrepertoire in human small
intestine and colon. J. Exp. Med. 180: 183-190.
36. KerstenCM, McCluskey RT, BoyleLA,
Kur-nickJT. (1996)Escherichia coli and
Pseudomo-nasaeruginosa induce expansion ofV82 cells
inadult peripheral blood, but of V81 cells in
cord blood. J. Immunol. 157: 1613-1619. 37. Duchman R, Kaiser E, Hermann E, Mayet
W, Ewe K, Meyer-Zum K-H. (1995)
Toler-ance exists towards resident intestinal flora but is broken in active inflammatory bowel disease. Clin. Exp. Immunol. 102: 448-455. 38. Janeway CA, Jr, Jones B, Hayday A. (1988)
Specificityandfunction ofTcellsbearing
y6
T cellreceptors. Immunol. Today9: 73-77. 39. Munk ME, Schoel B, Modrow S, Karr RW,
Young RA, Kaufmann SHE. (1989) T
lym-phocytes from healthy individuals with specificity to self-epitopes shared by
myco-bacterial and human 65-kilodalton heat shock protein. J. Immunol. 143: 2844-2849. 40. Haregewoin A, Soman G, Hom RC, Finberg
mycobacterial heat-shock protein. Nature 340: 309-312.
41. Ebert EC. (1989) Proliferative responses of
human intraepithelial lymphocytes to vari-ous Tcell stimuli. Gastroenterology978: 1372-1381.
42. Pirzer UC, Schurmann G, Post S, Betzler M, Meuer S. (1990) Differential responsiveness
to CD3Ti vs. CD2-dependent activation of
human intestinalT lymphocytes. Eur. J.
Im-munol. 20: 2339-2342.
43. Qiao L, Schurmann G, Betzler M, Meuer S. (1991)Activationandsignalingstatusof hu-man lamina propria T lymphocytes.
Gastro-enterology 101: 1529-1536.
44. De Maria R, Fais S, Silvestri M, Frati L, Pal-lone F, Santoni A, Testi R. (1993) Continu-ous invivo activation and transient hypore-sponsiveness to TCR/CD3 triggering of human gut lamina propria lymphocytes.
Eur. J. Immunol. 23: 3104-3108.
45. Targan SR, DeemRL, LiuM,WangS, NelA. (1995) Definition of a lamina propriaT cell
responsive state:Enhanced cytokine respon-siveness of T cells stimulated through the
CD2 pathway. J. Immunol. 154: 664-675. 46. Fais S, CapobianchiMR,PalloneF, Di Marco
P,BoirivantM,DianzaniF,TorsoliA. (1991) Spontaneous release of interferon-y by
in-testinal lamina propria lymphocytes in
Crohn'sdisease. Kineticsofinvitroresponse to interferon 'yinducers. Gut 32: 103-107. 47. Mullin GE, Lazenby AJ, Harris ML, Bayless
TM, JamesSP. (1992)Increased IL2-messen-gerRNA in the intestinal mucosal lesions of Crohn's disease but not ulcerative colitis. Gastroenterology 102: 1620-1627.
48. Brynskov J, Tvede N, Andersen CB, Vrlien
M. (1992) Increased concentrationsof
inter-leukin 1,3, interleukin 2 and soluble
inter-leukin 2 receptors in endoscopical mucosal
biopsy specimens with active inflammatory
bowel disease. Gut 33: 55-58.
49. Fuss IJ, Neurath M, Boirivant M, Klein JS,
de laMotte C, Strong SA, FiocchiC, Strober
W. (1996) Disparate CD4+ lamina propria (LP) lymphokine secretion profiles in
in-flammatory bowel disease: Crohn's disease
LP cells manifest increased secretion of
IFN-,y, whereas ulcerative colitis LP cells manifest increased secretion of IL-5. J.
Im-munol. 157: 1261-1270.
50. BouillierS, CochetM,Poccia F, GougeonML. (1995) CDR3-independent y5 V81+ Tcell
ex-pansion in the peripheral blood of HIV-in-fected persons. J. Immunol. 154: 1418-1431.
51. Deem RL, Shanahan F, Targan SR. (1991)
Triggeredhumanmucosal Tcells release
tu-mor necrosis factor-alpha and interferon-gamma which kill human colonic epithelial cells. Clin. Exp. Med. 83: 79-84.
52. Graham MF, Diegelmana RF, Elson CO,
Lindblad WJ, Gotschalk N, Gay R. (1988) Collagencontentand typesinthe intestineal
stricturesof Crohn's Disease. Gastroenterology 94: 257-265.
53. Mathew RC, Weinstock RJ. (1994)
Granu-loma formation. In: Targan SR, Shanahan F
(eds). Inflammatory Bowel Disease, from Bench to Bedside. Williams and Wilkins, Baltimore, pp. 151-170.
54. Haas W, PereiraP,TonegawaS. (1993) y/8T
cells.Annu. Rev. Immunol. 11: 637-685. 55. Sartor, RB. (1994) Cytokines in intestinal
inflammation: Pathophysiological and clini-cal considerations. Gastroenterology 106: 533-549.
56. DohertyPC, Allan W,Eichelberger M, Card-ing SR. (1992) Roles of
af3
and yS T cell subsets inviral immunity. Annu. Rev.Immu-nol. 10: 123-151.
57. Hiromatsu K, Yoshikai Y, Matsuzaki G, Ohga S,MuramoriK,MatsumotoK,Bluestone JA,
Nomoto K. (1992) Aprotectiverole of y/8T
cells inprimaryinfection with Listeria mono-cytogenes in mice. J. Exp. Med. 175: 49-56.
58. RosatJP,MacDonald HR,LouisJA. (1993)A
role for y8+ T cells during experimental in-fection ofmicewith Leishmania major. J. Im-munol. 150: 550-555.
59. Skeen MJ, Zeigler HK. (1993) Induction of
murineperitoneal
y6
Tcells and their role in resistance to bacterialinfection. J. Exp. Med.178: 971-984.
60. Tsuji M, Mombaerts P, LeFrancois L, Nus-senzweig RS, Zavala F, Tonegawa S. (1993)
-y6Tcellscontribute to immunity against the liver stages of malaria in aB3 T cell deficient
mice. Proc.Natl. Acad. Sci. U.S.A. 91: 345-349. 61. Carding SR, Allan W, McMickle A, Doherty
PC. (1993) Activation of cytokine genes in T cells during primary and secondary murine influenza pneumonia.J. Exp.Med. 177:
475-482.
62. Scott S, Pandolfi F, Kurnick JT. (1990)
Fi-broblasts mediateTcell survival:Aproposed mechanism forretentionofprimedTcells.J.
Exp. Med. 172: 1873-1876.
Ad-L. D.McVay etal.: y6TCells and Inflammatory BowelDisease 203
hesion and costimulation of proliferative
re-sponses ofhumany6Tcellsby interaction of
VLA-4and VLA-5 with fibronectin. Immunol.
Lett. 35: 101-105.
64. Forrester JM, Newman LS, Wang Y, King
TEJ, Kotzin BL. (1993) Clonal expansion of lung V81+ T cells inpulmonary sarcoidosis.
J. Clin. Invest. 91: 4291-4302.
65. Soderstrom K,Halapi E, Nilsson E, Gronberg A, van Embden J, Klareskog L, Kiessling R.
(1990) Synovial cells responding to a 65-kD
mycobacterial heat shock protein have a
high proportionof a TCRy8 subtype
uncom-mon inperipheral blood. Scand. J. Immunol.
32: 503-515.
66. Smith MD, Broker B, Moretta L, Ciccone E,
Grossi CE, Edwards JCW, Yuksel F, Colaco
B, Worman C, MacKenzie L, Kinne R,
Weseloh G, Gluckert K,LydyardPM. (1990)
T y6 cells and their subsets in blood and synovialtissuefrom rheumatoid arthritis
pa-tients. Scand. J. Immunol. 32: 585-593.
67. Sioud M, K7jeldsen-Kragh J, Quayle A,
Kal-venes C, Waalen K, Forre 0, Natvig JB. (1990) The VS gene usage by freshly isolated
Tlymphocytes fromsynovial fluids in rheu-matoid synovitis: A preliminary report. Scand. J. Immunol. 31: 415-421.
68. Kjeldsen-Kragh J, Quayle A, Kalvenes C, Forre 0, Sorskaar D, Vinje 0, Thoen J, NatvigJB. (1990)Ty5cells injuvenile
rheu-matoid arthritis and rheumatoid arthritis. ScandJ. Immunol. 32: 651-659.
69. Bucht A, Soderstrom K, Hultman T, Uhlen
M, Nilsson E, Kiessling R, Groenberg A.
(1992) Tcell receptor diversity and
activa-tion markers in the V delta 1 subset of rheumatoid synovial fluid and peripheral blood T lymphocytes. Eur. J. Immunol. 22: 567-574.
70. Spiro HM. (1993) Complications of colitis: Extracolonic complications. In: Spiro HM (ed). Clinical Gastroenterology. McGraw-Hill,
New York, pp. 746-759.