Phage Genetic Sites Involved in
X
Growth Inhibition by the Escherichia coli
rap
Mutant
Plinio Guzman' and Gabriel Guarneros2
Departamento de Genitica y Biologia Molecular, Centro de Inuestigacibn y de Estudios Avanzados, Apartado Postal 14-740, Mixico 07000, D. F. Mixico
Manuscript received March 29, 1988 Accepted for publication November 17, 1988
ABSTRACT
T h e rap mutation of Escherichia coli prevents the growth of bacteriophage X. We have isolated phage mutants that compensate for the host deficiency. T h e mutations, named bar, were genetically located to three different loci of the X genome: b a d in the attP site, barll in the cIIZ ea10 region, and barIII within or very near the imm434 region. T h e level of X leftward transcription correlates with rap exclusion. Phage X mutants partially defective in the PL promoter or in pL-transcript antitermi- nation showed a Bar- phenotype. Conversely, mutants constitutive for transcription from the PI or pL promoters were excluded more stringently by rap bacteria. We conclude that rap exclusion depends on the magnitude of transcription through the wild type bar loci in the phage genome.
B
ACTERIOPHAGE X growth is inhibited in the Escherichia coli rap mutant (HENDERSON and WEIL 1976). This inhibition is affected by tempera- ture and phage immunity: the hybrid phage Ximm434cannot form plaques at 37 O or 42 O , whereas wild-type
X is inhibited at 42" but not at 37" (HENDERSON and WEIL 1976). Deletion or deletion-substitution mutants of one or both sides of the X attachment site (attp) overcome rap inhibition (HENDERSON and WEIL 1976). Mixed infection of rap bacteria with wild type and mutant phage resulted in a burst size reduction of the mutant phage whereas the burst of the wild type phage remained low (HENDERSON and WEIL
1976). This last result excludes explanations such as the production of a diffusible inhibitor by the wild type phage which is neutralized by the bar mutants, or the inactivation by the bar mutations of a site of action in X genome which overcomes the need for Rap function.
T h e nature of the E . coli Rap function and its role in phage X growth remains unclear. T h e rap mutation is recessive to rap' (GUARNEROS et al. 1987). This observation is consistent with the hypothesis that rap+ encodes a trans-acting product required for X growth. T h e rap site has been located at 26 min in the genetic
map of E. coli (GUARNEROS et al. 1987), very tightly
linked to the pth gene which encodes for peptidyl- tRNA-hydrolase (ATHERLY and MENNINCER 1972; HOVE-JENSEN 1983). At least one pth mutant tested showed the Rap- phenotype indicating that both mu- tations affect the same exclusion function (GUARNE-
' Present address: Department of Biology, Plant Sciences Institute, Uni-
' T o whom correspondence should be addressed. versity of Pennsylvania, Philadelphia, Pennsylvania 19104.
Genetics 121: 401-410 (March, 1989)
ROS et al. 1987). Whether the rap mutation affects Pth function has not been tested.
In this paper we report the isolation and character- ization of phage mutants which compensate for the lack of a good rap gene in the bacterial host. We name these new mutations bar for ''blanc0 de accion de
bar mutations. Known X mutants were also used to define further the essential properties of the Bar phenotype. In addition, we present genetic evidence implicating X transcription in rap exclusion.
- Rap." We report here the genetic location-of five X
MATERIALS AND METHODS
Bacteria, phage and plasmids: Escherichia coli K 12 and X
strains used in this study, their relevant genotypes, and their source o r reference are listed in Tables 1 and 2, respectively. Plasmids pPG 100, pPA 177, and pPA 150 are derivatives of pBR322 (BOLIVAR et al. 1977) which carry variable lengths of XattP DNA cloned into the BamHI site. In pPGl00 the HindIII-BamHI DNA segment of pBR322 has been substi- tuted for the 492-bp HindIII(27479)-BamHI(27972) frag- ment from XlacW205 red114 imm434 DNA which harbors the attP site. An equivalent plasmid has been constructed by other authors (Hsu, ROSS and LANDY 1980). Plasmids pPA177 and pPA150 harbor shorter XattP DNA segments (see Figure 6) (MIZUUCHI and MIZUUCHI 1980).
Isolation of X bar mutants: XlacW205 red 1 14 imm434 was the parental phage for the isolation of bar mutants; this phage does not form plaques on the rap host DH173 at 37' or 42". The bar mutants were selected by growth on DH173. The bar mutants were selected as rare plaques which plated on strain DH173 either from untreated or from hydroxylamine mutagenized phage stocks ( M O N T A ~ E Z
E . coli K12 strains (deletion mutants) were discarded (PARKINSON and HUSKEY 197 1).
C600
DH 173 S60 1 K936 N2668
GG95 GG191 GGl98 GG240 GG228 GG289 GG609
thrl leuB6 thil lacy1 supE44 tonA21
C600 lac amyl4 rap
trp gal recA himA42 lig7(ts)
S601(h)
supF Alac gal (P2) supF Alac gal recA13
S 165 lac recA" D H l 7 3 recA" GG198 (Ximm434) GGl9l(X)
Strain Relevant Source
designation genotype or reference
Media: Cells were grown on LB (10 g tryptone, 5 g yeast extract and 5 g NaCl per liter, pH 7.2) or TB (10 g tryptone and 5 g NaC1, pH 7.2) supplemented with 0.2% of D-
APPLEYARD (1954) maltose. T h e differential medium for lac or nul transducing
HENDERSON and WEIL (1976)
SMITH (1 975)
MILLER and FRIEDMAN (1980)
GOTTESMAN, HICKS and
Our collection
Our collection Our collection Our collection Spontaneous lac This work
GELLERT (1 973)
phage was tetrazolium lactose and tetrazoiium galactose, respectively (OHLSSON, STRICINI and BECKWITH 1968). Cells harboring pBR322 or derivatives of this plasmid were grown in media supplemented with ampicillin at 50 pg/ml.
Phage crosses: T h e procedure for lytic crosses has been reported elsewhere (CUARNEROS and ECHOLS 1973). T h e crosses in Figure 1 were done in strain GG 198 at 34 O , other crosses were done in strains K936 or C600 at 40" to reduce site specific recombination which depends on the naturally temperature sensitive Xint gene product (GUARNEROS and
ECHOLS 1973) and on the bacterial himA gene product
(MILLER and FRIEDMAN. 1980).
O u r collection Testing for Bar phenotype: T h e Bar phenotype of the
sr1::TnlO derivative of strain N5130 (M. GOTTESMAN) as donor. coli rap at 40". The efficiency of plating (eop, plaques on strain DH173 divided by plaques on strain C600) for X bur stocks. The mutant bur205 arose spontaneously. All the mutants is close to 1, at least 10,000-fold higher than that mutants yielded about equal number of plaques on strain for the parental phage XlucW205 red114 imm434. T o ex- C600 and DH173. Since we were interested in point muta- amine the activity of bur in the uttP DNA segment cloned tions, those X bur mutants selected on DH173 that were in pBR322 derivatives (see Figure 6), phasmids were ob- Constructed by p1 mediated cotransduction of recA and phage was assayed by measuring their ability to plate On E'
TABLE 2
Bacteriophage X strains
Genotype
lacW205 red1 14 imm434 lacW205 A53 imm434 lacW205 A56 imm434 gal8 bioll imm434 cIIam4l gal8 imm434
biol I imm434 cIIam41 Jam6 int2 red 114 imm434
baa256 cI857 As-s cI857 ninL32 cI857 byp Aam32 gal8 biolO clam34 imm434 cI Oam29 cI857
Ab2 cI857 cI857 cro27 Ab2 cI857 0-027 intC226 cI857 Ab2 intC226 cI857 sex1 cI857 nutL44 imm434 Aint-xis3080 cI857
lacW205 bio936 red3 imm434
ninL63 imm434
lacW205 As-x imm434
lacW205 intam29 red10 imm434
Source, reference or derivation"
GUARNEROS et al. (1 987)
OPPENHEIM, GOTTESMAN and GOTTESMAN (1982)
OPPENHEIM, GOTTESMAN and GOTTESMAN (1 982)
Our collection Our collection Our collection Our collection NIH collection
D. COURT
J. SALSTROM
Our collection O u r collection O u r collection Our collection
EISEN et al. '( 1970)
O u r collection
SHIMADA and CAMPBELL (1974) via NIH collection
Our collection
M . GOTTESMAN
Our collection
R. WEISBERC
Xga149 bio936 red3 cI857
X
XlacW205 bioll imm434 Xbio7-20 ninL63 imm434
X
Ximm2l Oam29 hbio7-20 A.-x cI857
X
XlacW205 red114 imm434
XDaml5 b515 b519 intam29 red10 imm21
X
XlacW205 bioll imm434 ~
rap Inhibition of Phage Growth
a
X
b
-7, redf/4 lim, 434
3
5
4
1o-H &io IfI
I h m e n m 4 110 f
i i 4cEm41 a/ 8 imU34
FIGURE 1.-Location of X bar mutations by Int mediated crosses. The lacW205 red114 imm434 phage (heavy line a) was either the barl02,
102, 203, 205 or 3 0 4 mutant. The lacW205 b i o l l imm434 cIIam41 recombinants were selected as Spi (ZISSLER, SIGNER and SCHAEFER 1971) gal- plaques on strain GG191 lac gal (P2) and the %a18 red114 imm434 reciprocal recombinants as gal plaques on strain GG198 lac gal recA. These phage carry the left or the right arms (heavy lines c and b) respectively, from the bar parental genomes and were used, in turn, as parentals for a second set of crosses as indicated in b and c. The Aredl14 imm434 and XlacW205 imm434 recombinants were selected as gal- plaques o n strain GG198. These recombinants were tested for Bar phenotype by plating on the rap strain DH173. Only one of the two possible recombinants out of 20 plaques tested, for each of the original mutants, was & .
-
bar. These crosses positioned b a r l 0 2 , 102 and 203 to the left, and bar205 and bar304 to the right of attP.tained by recombination between X phage and each of the respective plasmids. C600 cells harboring the different plas- mids were infected with XlacW205 red114 imm434 and the lysates spotted on lawns of C600 on T B plates. Lysogens from cells infected with recombinant phasmids were selected by streaking out on LB-ampicillin medium. The resulting colonies were grown in liquid ampicillin medium and in- duced with mitomycin C at 2 pg/ml. Infection of bacteria harboring pBR322 lacking the cloned attP DNA segment did not yield Ap' colonies. Lysates of about 10' to pfu per mi were obtained and assayed for their Bar phenotype, as described above.
RESULTS
Genetic location of bar loci: Deletions or deletion- substitutions that remove XDNA on either side of the phage attachment site, attP, are bur (HENDERSON a n d WEIL 1976). T h u s , we initially mapped the putative bar point mutations relative to attP. T h e mutagenized phage carried lacW205 (OPPENHEIM, GOTTESMAN a n d GOTTESMAN (1 982), a deletion-substitution that elim- inates most XDNA sequences between theJ gene and t h e Hind111 restriction site a t position 27479, left of
attP; this phage is phenotypically Bar+. Therefore it
is likely that a bar locus lies between the lacW205 substitution and the core in attP. Two sets of Int mediated genetic crosses (Figure l ) , positioned three of t h e bar mutations to the left and two to the right of t h e attP site. Since Int-mediated recombination with the five bar mutants occurred at the same fre-
+
/ucW2tWH
bio 11 ] ihm434cIm41" 7
I
I
I
I
Jam6
L """"""""""- "- """"""" iit2 red114 ihm 434
FIGURE 2.-Mapping of bar1 mutants by homologous recombi-
nation. Phage XlacW205 bioll imm434 cIIam41 carried the barl01, 102 or 203 mutations and were obtained from cross a in Figure 1.
The heavy line indicates the location of the bar mutations as inferred from the results in Figure 1. The lac transducing (solid line) and nontransducing (dashed line) recombinant phage were screened by plaque phenotype among X J' gL - + recombinants selected on strain
GG288 lac sup' recA grown on indicator plates. Strain GG288 prevents the growth of both parental phage. The Bar phenotype of 100 plaques of each recombinant of the indicated types was tested by plating on strain GG289 rap recA. Out of 100 plaques in each case 11, 7, and 4 lac bar recombinants were found for bar101, 102 and 1 0 3 respectively (for further details see MATERIALS AND METH-
om).
quency as for a wild type phage (Figure 1, data not shown), the mutants are unlikely to affect attP or t h e int gene.
Fine mapping of X bar mutations: T h e five bar mutations were mapped more precisely by homolo- gous DNA recombination. The three mutations pre- viously located to the left of attP were mapped relative to the lacW205 deletion-substitution. From recombi- nants selected in the cross represented in Figure 2,
phage were found among 100 recombinants tested, whereas about 7% of the lac+ recombinants tested were bar-. These results imply that bar is located between the lacW205 substitution and the attP cross- over site. T h e low frequency of bar rescue among lac+ recombinants suggests that the position of the bar mutations is much closer to the core of attP than to the lac DNA substitution.
Two mutations, bar205 and bar304 previously po- sitioned to the right of attP were mapped relative to
the bio256 and imm434 regions (Figure 3). T h e bio256
mutation substitutes X DNA from attP to the 3' ter- minal N gene sequences leaving about 480 bp of X DNA between bio and imm434 (FRANKLIN and BEN- NETT 1979, DANIELS et al. 1983). We crossed bar
imm434 and bio256 cI857 phage as indicated in Figure
3. T h e mutation, bar205, was found in 40 out of 40 phage which had recombined between bio256 and
imm434 and which carried the imm X region, whereas
the other mutation, bar304, was not found (0 out of 40) in such recombinants (Figure 3). These results indicate that bar205 maps to sequences deleted by the
bio256 substitution, i.e., the attP-N region, and that
bar304 maps to the right of the N gene region.
The bar205 mutation: T o locate the bar205 muta- tion more precisely, its map position was determined relative to two other bar+ deletion mutations in the
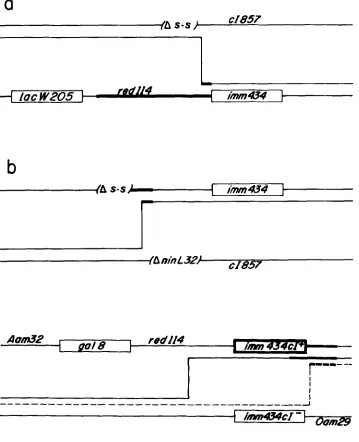
attP-N region (Figure 4). T h e As-s deletion removes X
DNA between the two Sal1 sites at positions 32745 and 33244, thereby eliminating part of the bet and gam genes (COURT and OPPENHEIM 1983). T h e
ninL32 deletion, though not precisely defined, elimi-
nates DNA in the ealO-N region (INOKUCHI and DOVE 1978; FRANKLIN and BENNETT 1979). Recombinants of the type shown in Figure 4a carried the bar+ or
bar- alleles with similar frequencies in about 100
plaques tested, suggesting that bar205 is located be- tween As-s and imm434. A XAs-s imm434 bar- recom- binant from this cross was used as one of the parental phage in the cross depicted in Figure 4b. Recombi- nants of the type shown, generated by crossovers between the As-s and AninL32 deletions, yielded 38%
of bar- phage in about 100 recombinants tested. All
these results located bar205 between the two deletions in a region including the genes kil, C H I , ea10 and ral.
The bar304 mutation: T h e bar304 mutation, pre- viously located to the right of the bio256 deletion- substitution, was mapped relative to a cI mutation of
imm434 and an amber mutation in the 0 gene. Among
the am+ recombinants in the cross shown in Figure 5 all the cI+ and 32% of the cI-, of 100 plaques tested in each case, were bar. This result indicates that
bar304 is located between the cI and Oam29 muta-
tions.
Functional mapping of bar sites using deletions:
bio 256
I
cf 857FIGURE 3,"Genetic location of bar205 and bar304. T h e XlacW205 red114 imm434 phage were either bar205 or bar304 mutants. T h e heavy line indicates the segment of the X genome where bar mutations were located, from results in Figure 1 . T h e indicated recombinants were selected by plating on the strain GG609 lac recA (himm434) which prevents the growth of both parental phage. Forty recombinant plaques were further purified on strain GG198 supF lac recA, and screened for Bar phenotype on
DH 173 rap a t 40".
T h e mutations described above defined several bar sites on the X genome. T o examine the extent of these sites, deletion and deletion-substitution mutants were tested for their Bar phenotype.
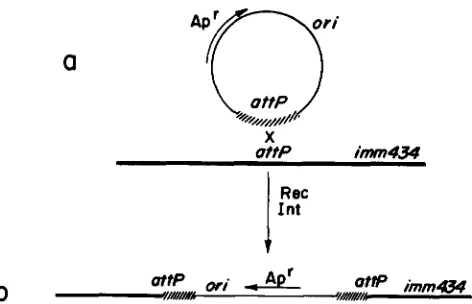
Plasmids harboring different deletions of the attach- ment site, attP, were used to delimit the bar1 region. These plasmids are substrates for integrative recom- bination (MIZUUCHI and MIZUUCHI 1980). We carried out functional mapping of bar1 with recombinant phasmids between X phage and each of the attP dele- tion plasmids (see Figure 7). The results in Figure 6 (lines 1 to 3) show that the phasmids that carry at least a 127-bp DNA segment of attP are Bar+, whereas phasmids that carry only a 26-bp fragment are Bar-. These segments define the size limits for an active
bar1 region and suggest that the recombinant phas-
mids are a useful tool for Bar functional analysis. Deletions, or gal or bio deletion-substitutions which affect the attachment site are Bar- (HENDERSON and WEIL 1976). T o determine the left boundary of the bar region near the attachment site, we tested the Bar phenotype of phage carrying deletions within the 248- bp DNA segment flanked by the Hind111 site at posi- tion 27479 and the core in attP (Figure 6, lines 4-6). This analysis showed that phages carrying segments
of attP DNA between positions -1 60 and -90 are
Bar'. Thus, this result together with the previous one obtained with phasmids located a bar1 boundary be- tween 90 and 26 bp to the left of the core in attP.
T h e bar1 boundary to the right of the attP region has not been as precisely defined. Two phage deletion mutants have been tested: one, XAredIO (STERNBERG et al. 1979), which leaves intact about 870 bp to the right of the attP core, is bar+; the other, XAint-xis3080 (R. WEISBERG, unpublished data), which deletes a segment of DNA from attP through int and xis, is bar. In addition a deletion-substitution mutant, Xbi0936, which removes only 4 1 bp to the right of the attP core (DANIELS et al. 1983) is also bar. Thus the DNA region adjacent to the attP core on the right side contains
bar, but its extent is thus far poorly defined.
The bar11 region, defined by the bar205 mutation,
rap Inhibition of X Phage Growth
a
(A
s-s)
d B 5 7b
AuM2
red114
1
i m 4 3 4 c g
r---
"I
I I
I
"-"""""
""""""""""""-1(see above). A third deletion, As-x, which eliminates the same DNA segment as As-s plus a contiguous 254-
bp sequence up to the XhoI site between the cIII and ea10 genes at position 33498 of X DNA, is Bar-. Thus at least part of the bar11 region lies between the Sal1 (33244) and XhoI (33498) restriction sites.
Transcriptional effects involved in Rap exclu- sion: Infection of rap bacteria with known X mutants has shown the involvement of phage transcription in Rap inhibition. T h e sex1 mutant of X , defective in the p L promoter (ROBERTS 1969) and the nutL44 muta- tion, which prevents pL transcript elongation beyond the transcription terminator tL1 (SALSTROM and SZY- BALSKI 1978) are phenotypically Bar- (Table 3, lines
3 and 4). In contrast, the intC226 mutation, which
FIGURE 4.-Fine genetic mapping of bar205. (a) The XlacW205 red114 imm434 phage carried the bar205 mutation. The heavy line indicates the location of the bar205 mutation as inferred from the results in Figure
3. The Spi- imm434 recombinants were selected on the strain GG240 supF lac (X, P2) and they were further purified on strain GG191 suPF (P2). All recombinants tested on DH173
rap were bar. One of these recombi- nants was used as a parental phage in cross b. The indicated recombinants (b) were selected by plating on the strain GG95 recA (X) and further pu- rified on strain S601 recA. The Bar phenotype of the recombinant plaques was tested on DH173 rap
38% of about 100 recombinants tested were bar. The heavy line in- dicates the position of bar205 as in- ferred from the results of the cross in a (see text).
FIGURE 5,"Further genetic loca- tion of mutation bar304. The paren- tal phage XAam32ga18 red1 14 bar304 imm434 was obtained by crossing the XlacW205 red1 14 imm434 bar304 mutant with XAam32 gal8 biolO cIam34 and selecting for gal+ plaques on strain GG198 gal lac recA. The selected xA32 imm434 candidates car- ried bar304. The red+ O+ recombi- nants indicated in the cross were se- lected by plating on strain N2668
l i d t s ) sup" which prevents the growth of both parental phage. Phenotypi- cally the recombinants were cI+ (solid line) or cI- (dashed line). One hundred plaques of each type were tested for bar on strain DH173. The heavy lines indicate the position of bar304 as inferred from the results of these crosses.
causes constitutive transcription from the
PI
promoter across the attP site (SHIMADA and CAMPBELL 1974),and the cro27 mutation defective in the repression of pL (EISEN et al. 1975), cause more severe inhibition of the respective phage in a rap host (Table 3, lines 5
and 7 ) . This particular phenotype was designated Bar' (for super bar), and was first observed by S. GOTTES- MAN (personal communication) with XintC226. The
intC and cro27 mutations in combination with the Ab2 deletion conferred different phenotypes. T h e Ab2 intC226 phage was Bar+, an intermediate phenotype between Bar' of AintC226 and Bar- of XAb2. In con- trast, the double mutant Ab2 cro27 retained a Bar' phenotype of the cro27 mutant (Table 3, lines 6 and
TABLE 3
Bar phenotype of X mutants affected in transcription
X Phage" Phenotype*
1 + Bar'
2 Ab2 Bar-
3 sex1 Bar-
4 nutL44 Bar-
5 cro27' Bars
6 Ab2 cro27' Bars
7 intC226 Bar"
8 Ab2 intC226 Bar'
a All the phage were cI857 except XnutL44 imm434.
* The Bar phenotype was determined by evaluating the respec- tive eops at 40" (No. plaques on rup-/No. plaques on rap') as described in MATERIALS AND METHODS. Bar', eop 0.5 (small and irregular plaques); Bar-, eop about 1.5; Bar', eop < (cro27) or about (intC226).
' cro27 phage were assayed at 39"; these phage do not grow at temperatures below 37' or above 40' (EISEN et al. 1970).
direct correlation between the level of transcription through the bar regions and the stringency of Rap inhibition.
DISCUSSION
We have isolated bacteriophage X mutants which overcome the need for E . coli rap+ gene for growth. Their respective mutations, named bar, were geneti- cally located to three regions of the phage genome: burl overlaps the phage attachment site attP; bar11
maps in the cIII-ea10 region, and bar111 lies within, or near the imm434 region (Figure 8). We have also shown that phage growth inhibition in rap bacteria
region by functional analysis of dele- tions. The phage assayed were XlacW205 A53 imm434, XlucW205
A56 imm434 and XAb2 c1857 (see MA-
TERIALS AND METHODS). The plas- mids pPG100, pPA150, or pPA177 were first crossed with the bur' phage XlacW205 red114 imm434, and the recombinant phasmids were then as- sayed for Bar phenotype on strain DH173. The line segments show the XDNA present in the phage or in the plasmids assayed; pPA150 carries, in addition, a transposed DNA segment from +144 to +174 (MIZUUCHI and
MIZUUCHI 1980). Segments are
aligned at the zero position (the cen- tral base pair in the Xatt core). The numbers at the ends of the segments indicate the base pair positions at the X DNA limits. The Bar' phenotype is defined as 10.000-fold less eop than bar- phage. The Bar- pheno- type for recombinant phasmids showed an eop about 20-fold higher that of the bur+ recombinants.
correlates with transcription from the X pL or PI
promoters.
T h e data presented here define the left boundary of the bar1 region to within a 90 bp DNA segment in the left arm of the attP site present in the XA56
deletion (Figure 6, line 5). T h e barI region extends into the right arm of attP for at least 40 bp, as defined by the Bar- phenotype of the Abio936 deletion-substi- tution phage (DANIELS et al. 1983). Thus the barI region straddles the core and spans as much as 130 bp of the attP site. Our results with the recombinant phasmids between X bar+ phage and each of the two different plasmids which harbor shortened attP sites, are only partially conclusive. T h e plasmid pPAl50, which harbors a 26-bp segment of attP DNA, gener- ated a Bar- phasmid (Figure 6, line 3). This observa- tion indicates that the 26 bp segment carries an incom- plete barI region. We cannot derive, however, a firm conclusion from the Bar+ result obtained with the plasmid pPA 177, which carries a 1 14-bp segment of XattP DNA (Figure 6, line 2). Since these phasmids carry two shortened attP sites and probably two short- ened burl regions (Figure 7), it is not clear whether one or both barI regions are functional for rap exclu- sion.
rap Inhibition of Phage Growth
FIGURE 7.-Construction of phage-plasmid recombinants for
Bar phenotype analysis. Plasmid derivatives of pBR322, which
harbor different segments of XattP DNA were recombined with a
X bar+ phage carrying a subnormal supply of DNA (a). The insertion
recombinants (b), mediated either by the phage Int or the bacterial
Rec system, were assayed for Bar phenotype on rap bacteria (see
MATERIALS AND METHODS). A correlation was found between the
size of the cloned attP segment and the Bar+ phenotype of the
recombinant phasmid (see Figure 6). Notice that the insertion recombinants carry two hybrid attP sites which may harbor short-
ened attP right and left arms according to the size of the DNA
segment cloned in the plasmid. Slashes indicate X homologies at
attP.
alleviates E . coli rap exclusion, was genetically mapped between the J gene and attP of the X chromosome (HENDERSON and WEIL 1976). It is likely that mutl is a bar1 type mutation because there are no known bar sites in the Ab region other than barI. In fact, the lacW205 deletion-substitution eliminates several kb of DNA of the b region without affecting the Bar phe- notype. T h e location of the barI region reported here is consistent with earlier observations showing that phage deletions or deletion-substitutions affecting the attP site are able to grow on rap bacteria (HENDERSON and WEIL 1976).
Two other sites, bar11 and barIII, are reported here. T h e bar11 region, defined by the bar205 and As-x mutations, was located to the ea10 cIII region (see Figures 4 and 8, and the section on Functional map- ping of bar sites using deletions). Neither cIII nor the neighboring ea10 active gene products appear to par- ticipate directly in rap exclusion because the mutant XcIIJ67 (data not shown) and the deletion mutant XninL32 (Figure 4), which removes part of the coding sequence for ealO, are Bar+. T h e bar111 region, de- fined by genetic mapping of the bar304 mutation is within, or near, the right end of the imm434 region (Figures 5 and 8). T h e bar111 mutation could affect a transcription regulatory element in imm434 or it could alter a bar region analogous to barI and barII. It has been shown that phage immunity type is important for the Bar phenotype: Ximm434 is more severely excluded than immX in rap bacteria (HENDERSON and WEIL 1976). This observation may be related to the high level of leftward transcription from the pL pro-
moter in 434 hybrids (PTASHNE 1971) (see below). We observed a direct correlation between the mag- nitude of transcription through the barI and bar11 regions and X phage exclusion in E. coli rap. Both the barI and bar11 regions are part of the X left operon and therefore are transcribed by the N-antiterminated RNA polymerases which initiate at pL (Figure 8)
(COURT and OPPENHEIM 1983; SCHMEISSNER et al. 1984). T h e data presented here show that mutations which reduce transcription through bar1 and bar11 by impairing either initiation at the pL promoter (sexl), or the N-antitermination of the p L initiated transcript (nutL44) confer a Bar- phenotype (Table 3, lines 3 and 4). Conversely, constitutive pL transcription in the cro27 mutant resulted in more severe rap exclu- sion of phage growth (Bars phenotype, Table 3, line
5). The X pI promoter located about 8 kb downstream of p L , allows transcription into the int gene and through barI region (Figure 8) (SCHMEISSNER et al.
1984). The XintC226 mutant which causes constitutive transcription from PI (SHIMADA and CAMPBELL 1974), also shows a BarS phenotype (Table 3, line 7). The epistasis of cro27 to Ab2 in the phage carrying both mutations (Table 3, line 6) probably reflects the con- trol of cro gene product on the transcription of both the bar11 region and the right segment of barI present in the Ab2 deletion; intC, which transcribes constitu- tively only the segment of bar1 region present in Ab2, shows an intermediate Bar+ phenotype (Table 3, line 8). Further experiments are necessary to understand these epistatic relationships.
Taken together we interpreted the results in Table 3 to mean that transcription through bar regions is a primary effect in the rap exclusion of X. Experimental results with plasmids lend support to this proposal: an active promoter cloned upstream of a X bar region prevents maintenance of the plasmid in rap bacteria
(P. GUZMAN, unpublished data).
Any explanation for rap exclusion has to take into account the results of burst size from the mixed infec- tion experiments discussed previously (see Introduc- tion, HENDERSON and WEIL 1976); it also has to be consistent with the correlation between the magnitude of transcription through bar regions and the extent of phage exclusion. Accordingly we propose that tran- scription through bar regions somehow inactivates an essential function limiting in the rap mutant. T h e more transcription the more severe the exclusion. In this model the burst sizes of X+ and X bar in mixed infection of rap bacteria are dependent on the avail- ability of the essential function. The affected function, in rap bacteria could be the peptidyl-tRNA-hydrolase.
This product is essential for the bacteria as indicated by the fact that a thermosensitive pth mutant has a thermosensitive hydrolase (ATHERLY and MENNINGER
I
imm434t I O t t P P I
cRteol0
tu N nutL P L CIpR cfo t R 01 I I I
intC226 nutL44 sex1
bar
I
barI
cro27 borII
"
--
FIGURE 8,"Transcription through Xbar sites. Transcripts (wavy lines) from PI, pL, and pR and the location of bar regions are indicated. The arrowheads on the wavy lines represent transcription termination sites. Continuation of the lines beyond the arrowheads indicate antitermination mediated by AN protein. Also shown are the locations of sites, genes, mutations, the Ab2 deletion, and the imm434 substitution. The map is not drawn to scale. , .
putative bar point mutations, and functional analysis of the E . coli rap strain may provide new clues for the understanding of this novel host-phage interaction.
We are grateful to REFUGIO GARC~A for measuring the eops in Table 3, to DON COURT, KIYOSHI MIZUUCHI, JOHN SALSTROM and BOB WEISBERG for phage strains and plasmids, to SUE GOTTESMAN
for sharing unpublished observations, to D. COURT, ALLISON RAT- TRAY and CECI MONTAAEZ for critical review of the manuscript, and to ROSY CRUCES, NACHO CRUZ and GUSTAVO JUAREZ for the skillful preparation of the manuscript. This work was supported by research grants from the Consejo Nacional de Ciencia y Tecnologia and the Fundacibn Zevada, in Mexico. During the development of these experiments G . G. was supported by a fellowship from the John Simon Guggenheim Foundation.
LITERATURE CITED
APPLEYARD, R. K., 1954 Segregation of new lysogenic types dur- ing growth of a double lysogenic strain derived from Escherichia coli K12. Genetics 39: 440-452.
ATHERLY, A. G . , and J. R. MENNINGER, 1972 Mutant E. coli strain with temperature sensitive peptidyl-transfer RNA hydrolase. Nature New Biol. 240: 245-246.
BOLIVAR, F., R. L. RODRIGUEZ, P. J. GREENE, M. C. BETLACH, J. L. HEYNEKER, H. W. BOYER, J. H. CROSA and S. FALKOW,
1977 Construction and characterization of new cloning ve-
hicles. 11. A multipurpose cloning system. Gene 2 95-1 13.
COURT, D., and A. B. OPPENHEIM, 1983 Phage lambda's accesory genes, pp. 251-278 in Lambda 11, edited by R. W. HENDRIX, J. W. ROBERTS, F. W. STAHL and R. A. WEISBERG. Cold Spring
Harbor Laboratory, Cold Spring Harbor, NY.
DANIELS, D. L., J. L. SCHROEDER, W. SZYBALSKI, F. SANGER and F. R. BLATTNER, 1983 Appendix I , A molecular map of coli- phage lambda, pp. 469-517 in Lambda 11, edited by R. W. HENDRIX, J. W. ROBERTS, F. W. STAHL and R. A. WEISBERG. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY. EISEN, H., P. BRACHET, L. PEREIRA DA SILVA and F. JACOB,
1970 Regulation of repressor expression in X . Proc. Natl.
Acad. Sci. USA 66: 855-862.
EISEN, H., M. GEORGIOU, C. P. GEORGOPOULOS, G . SELZER, G . GUSSIN and I. HERSKOWITZ, 1975 The role of gene cro in phage development. Virology 68: 266-269.
FRANKLIN, N. C., and G . N. BENNETT, 1979 The N protein of bacteriophage lambda, defined by its DNA sequence, is highly basic. Gene 8: 107-1 19.
GOTTESMAN, M. M., H. L. H I C K S ~ ~ ~ M. GELLERT, 1973 Genetics and function of DNA ligase in Escherichia coli. J. Mol. Biol. 77:
531-547.
GUARNEROS, G . , and H. ECHOLS, 1973 Thermal asymmetry of
site-specific recombination by bacteriophage X. Virology 52:
GUARNEROS, G . , G. MACHADO, P. GUZMAN and E. GARAY,
1987 Genetic and physical location of the Escherichia coli rap
locus, which is essential for growth of bacteriophage lambda. J. Bacteriol. 1 6 9 5188-5192.
HENDERSON, D., and J. WEIL, 1976 A mutant of Escherichia coli that prevents growth of phage lambda and is bypassed by lambda mutants in a nonessential region of the genome. Virol-
HOVE-JENSEN, B. 1983 Chromosomal location of the gene encod- ing phosphoribosylpyrophosphate synthetase in Escherichia coli. J. Bacteriol. 1 5 4 177-184.
HSU, P. L., W. Ross and A. LANDY, 1980 The X phage att site: functional limits and interaction with Int protein. Nature 285:
INOKUCHI, H., and W. F. DOVE, 1978 Deletion of control sites in the pL operon of phage lambda. l'irology 88: 261-262.
MILLER, H. I., and D. I. FRIEDMAN, 1980 An Escherichia coli gene product required for lambda site specific recombination. Cell
20: 711-719.
MIZUUCHI, M., and K. MIZUUCHI, 1980 Integrative recombination of bacteriophage X : extent of the DNA sequence involved in attachment site function. Proc. Natl. Acad. Sci. USA 77: 3220- 3224.
MONTA~EZ, C., J. BUENO, U. SCHMEISSNER, D. L. COURT and G . GUARNEROS, 1986 Mutations of bacteriophage lambda that define independent but overlapping RNA processing and tran- scription termination sites. J. Mol. Biol. 191: 29-37.
OHLSSON, B. M., P. F. STRIGINI and J. R. BECKWITH, 1968 Allelic amber and ochre suppressors. J. Mol. Biol. 3 6 209-218.
OPPENHEIM, A,, S. GOTTESMAN and M. GOTTESMAN, 1982
Regulation of int gene expression. J. Mol. Biol. 1 5 8 327-346.
PARKINSON, J. S., and R. J. HUSKEY, 1971 Deletion mutants of bacteriophage lambda. I. Isolation and characterization. J. Mol. Biol. 5 6 369-384.
PTASHNE, H., 1971 Repressor and its action, pp. 221-237 in The Bacteriophage Lambda, edited by A. D. HERSHEY. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
ROBERTS, J. W., 1969 Termination factor for RNA synthesis. Nature 244: 1168-1 174.
SALSTROM, J. S., and W. SZYBALSKI, 1978 Coliphage X nutL-: a unique class of mutants defective in the site of N product utilization for antitermination of leftward transcription. J. Mol. Biol. 1 2 4 195-221.
SCHMEISSNER, U., K. MCKENNEY, M. ROSENBERG and D. COURT,
1984 Removal of a terminator structure by RNA processing
regulates int gene expression. J. Mol. Biol. 1 7 6 39-53.
SHIMADA, K., and A. CAMPBELL, 1974 Int-constitutive mutants of bacteriophage lambda. Proc. Natl. Acad. Sci. USA 71: 237- 241.
30-38.
ogy 71: 546-559.
rap h
SMITH, G . R., 1975 Deletion mutations of the immunity region bination in growth of bacteriophage lambda 11. Inhibition of
of coliphge X. Virology 64: 544-552. growth by prophage P2, pp. 469-475 in The Bacteriophage
STERNBERG, N., D. HAMILTON, L. ENQUIST and R. A. WEISBERG, Lambda, edited by A. D. HERSHEY. Cold Spring Harbor Lab-
1979 A simple technique for the isolation of deletion mutants
of phage lambda. Gene 8: 35-51. oratory, Cold Spring Harbor, N.Y.