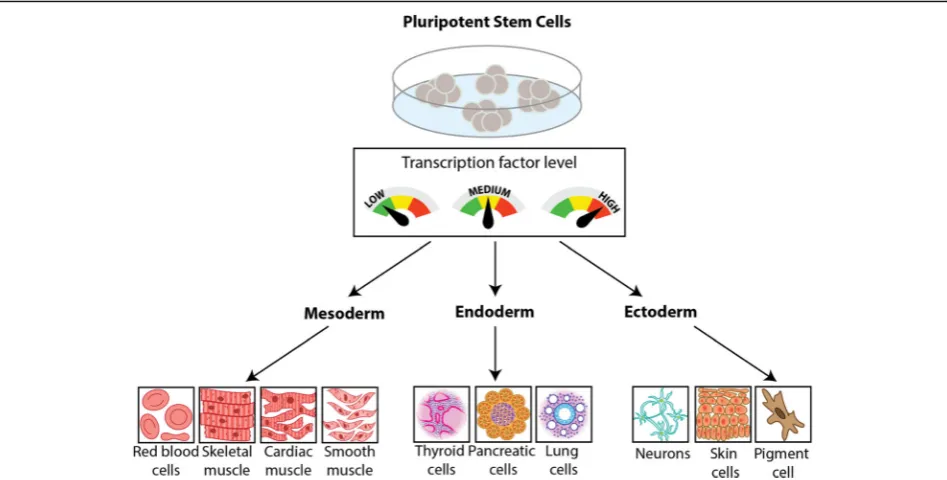
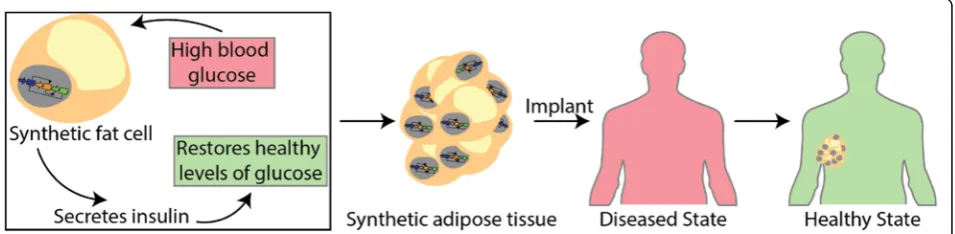
Genetic circuits to engineer tissues with alternative functions
Full text
Figure
Related documents
3(c) shows the performance on the task of Butterfly identification for our algorithm compared to random teacher on MTurk workers as we vary the length of teaching phase... 5.3
The purpose of our study was to assess the long- term efficacy and safety of implanting of two iStent inject Model GTS-400 stents combined with phacoemulsification in patients
Also, the deformation retract of the isometric folding of Eguchi-Hanson space and any folding homeomorphic to this type of folding is found to be different from the deformation
● BACULA, BAREOS (“open source” backup software) ● DP, NBU, TSM, NSR, GALAXY (commercial stuff) ● RBME (“open source” rsync backup made easy).. BACKUP type NETFS
However, this solution can be effective only during idle mode and most of the fuel consumption comes during the active mode (hoisting the containers). Figure 1-4 presents a
Eisenstat and Angluin (2007) proved, by giving a probabilistic construction of an abstract set system, that the upper bound of Theorem A is asymptotically tight if VC-dim ( R ) ≥ 5
DICTIONARIES WITH POLISH ANATOMICAL TERMINOLOGY Józef Mayer (anthropologist) and Fryderyk Sko- bel (physiologist), both professors at the Jagiello- nian University in 1838 published
But none of these blood vessels have been observed to enter the femoral head via this ligament (Figure 2). Figure 2: Ligamentum teres and synovial villi at 77 mm CRL fetus in
