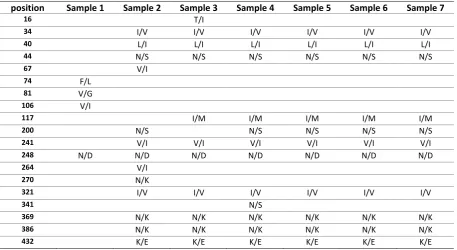
Molecular Characterization and Phylogenetic Analysis of Neuraminidase Gene in A/H1N1 Influenza Virus Isolates Circulating in Iran, 2014-2015.
Full text
Figure
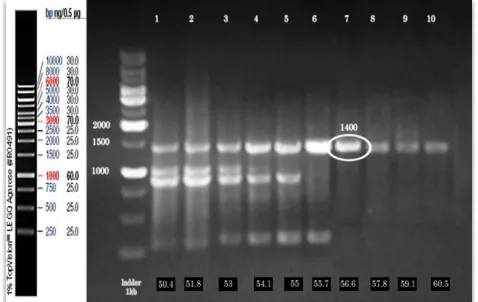

Related documents
Chen, “Some completely monotonic functions involving the gamma and polygamma functions,” Journal of the Korean Mathematical Society, vol. Chen, “Four logarithmically
A correlation seems to exist between chronic recurrent cer- vical nerve root neurapraxia and cervical canal stenosis in tackled football players [1,5-7] and risk of more
The present study undertakes a systematic approach to identify functionally important nsSNPs in human CALR gene in an attempt to understand how do these
The legal basis for the issuance of the Perpu, namely Article 12 of the 1945 Constitution, is the legitimacy of the Government to issue a legal norm or regulation
Particularly, such various aspects related to education are analyzed as: unequal development of the level of education in different states, which represents one of the key
The present work address itself to the investigations about the influence of barriers on the breakdown voltages in transformer oil for moderately homogeneous
Main purpose of this study is to examine to what extent Private Commercial Banks in Batticaloa District to make customers satisfaction and loyalty a special focus of
In addition, Higher- Order Path Terminating Equipment (HO PTE) and Lower Order Path Terminating Equipment (LO PTE) produce Remote Error Indications (REI) based on
