Original Article
Lower miR-145-5p expression and its potential
pathways in hepatocellular carcinoma: a
bioinformatics analysis with RNA-seq
and microarray data
Dongning Huang, Li Qin, Jian Huang, Haixin Huang
Department of Medical Oncology, Fourth Affiliated Hospital of Guangxi Medical University/Liuzhou Worker’s Hos-pital, Liuzhou, Guangxi Zhuang Autonomous Region, China
Received June 23, 2017; Accepted February 5, 2018; Epub April 15, 2018; Published April 30, 2018
Abstract:Background: The aim of our study was to investigate the clinical value of expression of miR-145-5p, and clarify its potential target genes and molecular function in hepatocellular carcinoma (HCC). Methods: Eleven micro-array datasets containing 325 HCC and 319 non-cancerous liver tissues were extracted from the Gene Expression Omnibus (GEO) database to evaluate the expression level of miR-145-5p in HCC. Additionally, we also collected
354 HCC and 50 normal liver tissues from The Cancer Genome Atlas (TCGA) database to confirm the differentially
expressed level of precursor miR-145 between HCC and normal liver tissues. Furthermore, the prospective target
genes of miR-145-5p in HCC were predicted and the possible correlative signaling pathways were clarified. Results: MiR-145-5p was evidently decreased in HCC tissues according to the GEO datasets, which was also proved by the data from TCGA. Altogether, 144 genes were considered as the most likely target genes of miR-145-5p for HCC.
Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis clarified that the most enriched pathways
included pathways in cancer, chronic myeloid leukemia and adherens junction. Nine hub genes (SMAD2, SMAD3, FOXO1, SMAD4, NRAS, MAPK1, SP1, MAPK9, CBL) with were screened out from the 144 genes, as evaluated by PPI network. Conclusions: The absence or down-regulation of miR-145-5p may play a pivotal role in the tumorigenesis
of HCC. MiR-145-5p may influence the development of HCC via targeting multiple genes and pathways, which need
to be further investigated.
Keywords: miR-145-5p, hepatocellular carcinoma, Gene Expression Omnibus (GEO), The Cancer Genome Atlas (TCGA), targets
Introduction
Hepatocellular carcinoma (HCC) is a
malignan-cy, which ranks the third most universal cause
for cancer-associated death in the whole world
[1-3]. Several risk factors including hepatitis B
or C virus infection, cirrhosis, aflatoxin and alco
-hol have been confirmed to be associated with
the occurrence of HCC [4-9]. Although
increas-ing improvements have been gained on the
diagnosis and treatment of HCC in the past
sev-eral years, the ovsev-erall survival of HCC remains
still an alarming situation [10-14]. The
five-year-survival rate was only 12% in advanced HCC
patients, and half of them had a median
sur-vival time of less than one year [15, 16]. Thus,
a precise diagnosis and timely treatment at
early phase is urgent for HCC patients.
available several studies were all based on
small size of patients [24-28]. Furthermore, the
expression data of miR-145-5p in public high
throughput datasets has never been utilized to
reveal its potential clinical significance in HCC.
When regarding the target genes of
miR-145-5p in HCC, only single target was confirmed by
different reports [26, 28-35], for instance, YTH
domain family 2 (YTHDF2), Rho-associated
coiled-coil kinase 1 (ROCK1), Fascin homolo-
gue 1 (FSCN1), Cullin 5 (CUL5),
metalloprotein-ase-17 (ADAM17), OCT4-pseudogene 4
[image:2.612.93.521.71.575.2]pg4), insulin receptor substrate 1 (IRS1) and
IRS2. No study has focused on a gene network
of the potential targets of miR-145-5p. Th-
erefore, a new study is needed to assess the
clinical influence and the potential signaling
pathways of miR-145-5p in HCC.
Considering the limitation of those previous
publications on miR-145-5p in HCC and the
new demand in HCC research, our current study
gathered the expression levels of miR-145-5p
in HCC and non-cancerous liver tissue from
Gene Expression Omnibus (GEO) microarray
datasets and The Cancer Genome Atlas (TCGA)
dataset, both of which contain high throughput
data of microarray or miRNA-sequencing of a
massive various types of cancer. We also
pre-dicted the potential target genes via online
software, including miRWalk, Microt4,
miRan-da, mirbridge, miRDB, miRMap, miRNAMap,
Pictar2, PITA, RNA22, RNAhybrid and Targe-
tscan. After that, bioinformatics analyses su-
ch as Gene ontology (GO) analysis, Kyoto
Encyclopedia of Genes and Genomes (KEGG)
pathway analysis and protein-protein
interac-tion (PPI) network were conducted to reveal the
prospective molecular mechanism of
miR-145-5p in HCC.
Materials and methods
Data extraction
We gathered the HCC-related miRNA
microar-ray datasets from GEO (https://www. ncbi.nlm.
nih.gov/geo/) and ArrayExpress (http://www.
ebi.ac.uk/arrayexpress/). The keywords used
in the search strategy were: “miRNA and
‘hepa-tocellular carcinoma’”. The inclusion criteria of
suitable microarray datasets were as following:
(i) the samples needed to be from human; (ii)
the microarray datasets consisted of HCC and
non-cancerous liver tissues; (iii) the sample
size should be more than 3 per study; (iv) the
expression level of the target miRNA,
miR-145-5p was securable. Additionally, the level 3
miRNA expression profile was downloaded from
TCGA (https://cancergenome.nih.gov/), which
included 50 non-cancerous normal liver and
354 HCC tissues, and their relevant
clinico-pathological parameters were also downloaded
from TCGA database for a further analysis.
Statistical analysis
The analysis of the collected data from GEO
and TCGA database was processed by using
SPSS 23.0. Quantitative variable was
comput-ed and presentcomput-ed as means ± standard
devia-tion (SD). The student’s t test was applied to
assess the alteration between two
indepen-dent quantitative variables. As for
meta-analy-sis, Stata 12.0 (College Station, Texas, USA)
was applied. Standard mean difference (SMD)
method was performed to assess the
differ-ence of miR-145-5p with continuous variables.
Q test and I
2statistic were performed to
evalu-ate the heterogeneity. If large heterogeneity
was existent (P<0.05, I
2>50%), the
random-effects model was selected to pool the SMD;
conversely, the fixed-effects model was utilized.
Receiver operator characteristic curve (ROC)
was performed to assess the diagnostic
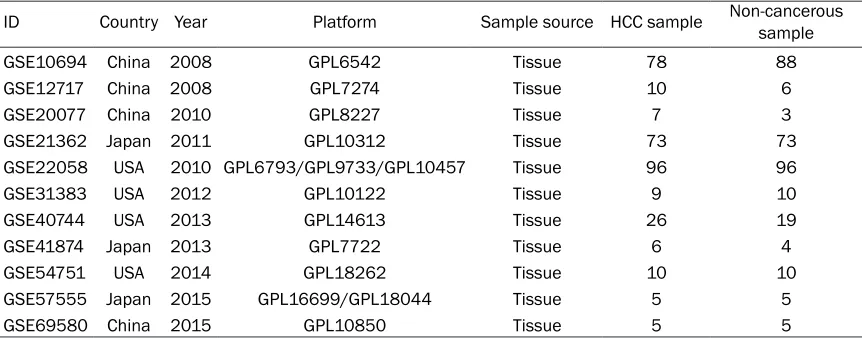
poten-Table 1.
Basic information of the 11 collected microarray datasets
ID Country Year Platform Sample source HCC sample Non-cancerous sample
GSE10694 China 2008 GPL6542 Tissue 78 88
GSE12717 China 2008 GPL7274 Tissue 10 6
GSE20077 China 2010 GPL8227 Tissue 7 3
GSE21362 Japan 2011 GPL10312 Tissue 73 73
GSE22058 USA 2010 GPL6793/GPL9733/GPL10457 Tissue 96 96
GSE31383 USA 2012 GPL10122 Tissue 9 10
GSE40744 USA 2013 GPL14613 Tissue 26 19
GSE41874 Japan 2013 GPL7722 Tissue 6 4
GSE54751 USA 2014 GPL18262 Tissue 10 10
GSE57555 Japan 2015 GPL16699/GPL18044 Tissue 5 5
[image:3.612.91.522.84.253.2]tial of miR-145-5p in HCC.
P
-value <0.05 was
considered to gain statistical significance.
genes were explored via the database of KEGG
pathways.
P
-value <0.05 was considered of
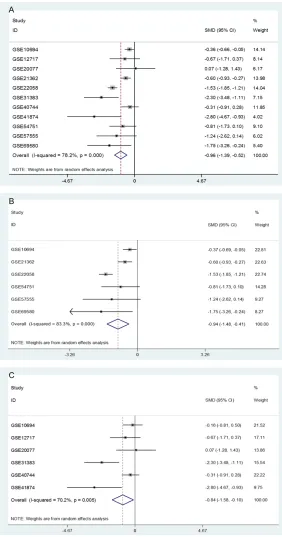
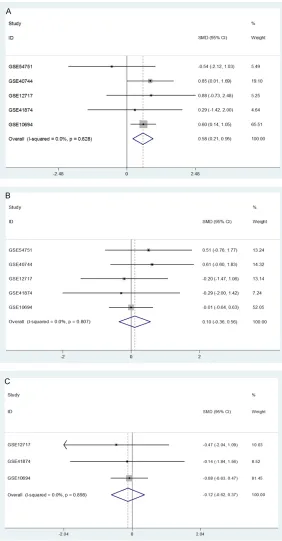
sig-Figure 2. Forest plots of the down-regulation of miR-145-5p in HCC tissues based on microarray data. A. Forest plot for miR-145-5p expression in HCC tissues versus non-cancerous liver tissues. B. Forest plot for miR-145-5p expression in the subgroup of HCC tissues and adjacent non-cancerous sues. C. Forest plot for miR-145-5p expression in the subgroup of HCC tis-sues and healthy liver tistis-sues.
Prediction of target genes
Twelve online target prediction
programs including miRWalk,
Microt4, miRanda, mirbridge,
miRDB, miRMap, miRNAMap,
Pictar2, PITA, RNA22, RNA-
hybrid, Targetscan were used
to predict the target genes
[36-40]. The potential target
genes of miR-145-5p were
required to be predicted by at
least six prediction platforms.
In order to achieve a more
comprehensive interactional
mechanism of targets of
miR-145-5p in HCC, we also
col-lected those validated targets
from literatures. The
search-ing key words were as
follow-ing: “(miR-145 OR miRNA-145
OR microRNA-145 OR miR145
OR miRNA145 OR
microR-NA145 OR “miR 145” OR
“miRNA 145” OR “microRNA
145”OR 145-5p OR
miR-NA-145-5p OR
microRNA-145-5p) AND target”. Only the
targets verified by dual
lucif-erase assay were gathered.
Furthermore, to restrict the
potential target genes in the
tumorigenesis and
develop-ment of HCC, the genes ac-
hieved above were interacted
with another group of genes
assessed by Chen’s group
with natural language
pro-cessing (NLP) analysis, which
includes 1,800
well-estab-lished genes of HCC [8, 41,
42].
GO and KEGG pathway analy
-ses of target genes
[image:4.612.90.372.70.607.2]nificance in GO and KEGG pathway analyses
[43-49].
The hub genes were uploaded to Search Tool
for the Retrieval of Interacting Genes/Proteins
(STRING) version 9.1 online tool
(http://string-db.org/) to establish PPI network. The STRING
software, collaboratively launched by European
Molecular Biology Laboratory (EMBL), Swiss
Institute of Bioinformatics (SIB) and University
of Zurich (UZH), is a database containing all
well-known and predicted protein interactions.
The interactions include direct (physical) and
indirect (functional) associations as derived
from four sources, comprised of literature
cura-tion, genome analysis and prediccura-tion,
high-throughput experimentations as well as
co-expression researches. The PPI network was
visualized by Cytoscape software (version
3.3.3) [47, 50-52].
Results
Down-regulation of miR-145-5p expression in
HCC based on GEO and TCGA databases
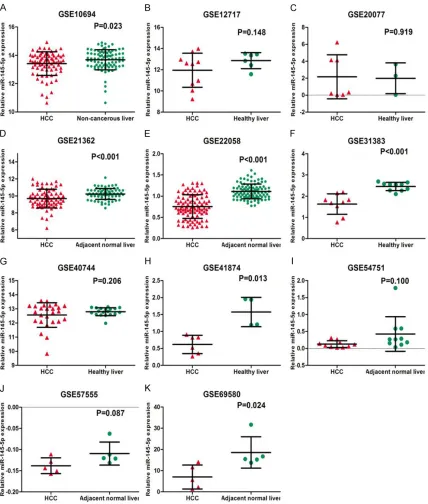
A total of 11 microarray datasets (GSE10694,
GSE12717, GSE20077, GSE21362, GSE22058,
GSE31383, GSE40744, GSE41874, GSE54751,
GSE57555, and GSE69580) were finally
incl-uded from GEO and ArrayExpress databas-
es according to the standard of inclusion.
Among the 11 microarray datasets, six
microar-ray datasets showed an apparently lower
expression level of miR-145-5p in HCC tissues
than that in adjacent normal tissues, including
GSE10694 (P=0.023), GSE21362 (P<0.001),
GSE22058 (P<0.001), GSE31383 (P<0.001),
GSE41874 (P=0.013), and GSE69580 (P=
0.024,
Figure 1
), while no obvious distinction
of miR-145-5p expression was noted between
HCC and adjacent liver tissues in the rest of
5 microarrays (GSE12717, GSE20077, GSE-
“40744, GSE54751 and GSE57555). In order to
get a better view of the clinical value of
miR-145-5p in HCC from different studies, a
com-prehensive meta-analysis was performed with
these 11 included microarrays (
Table 1
). Since
a major heterogeneity existed among these 11
microarray datasets (I
2=78.2%, P<0.001), a
random-effects model was selected. The
meta-analysis displayed that miR-145-5p expressed
markedly lower in HCC tissue than that in
non-cancerous liver tissue (SMD=-0.96, 95% CI:
-1.39 to -0.52, P<0.001,
Figure 2A
). Subse-
quently, due to different control groups
(adja-cent liver of HCC and healthy liver), subgroup
analysis was conducted. Results showed that
the expression of miR-145-5p was consistently
lower in HCC tissues than that in both of
adja-cent liver tissues (SMD=-0.94, 95% CI: -1.48 to
-0.41, P<0.001,
Figure 2B
), and healthy liver
tissues (SMD=-0.84, 95% CI: -1.58 to -0.10,
P=0.026,
Figure 2C
). We also divided the
patients into Asian group and Caucasian group.
In both groups, miR-145-5p was still markedly
down-regulated in HCC tissues than that in
non-cancerous tissues (data not shown).
The result of sensitivity analysis for GEO data
showed that the significant down-regulation of
miR-145-5p always existed no matter any of
the microarray datasets was removed, which
indicated the result of the current
meta-analy-sis based on included microarray datasets was
reliable (
Table 2
,
Figure 3
). The result of Begg’s
test (z=1.56, Pr>|z|=0.119,
Figure 4A
) and
Egger’s test (t=-0.74, Pr>|z|=0.478,
Figure 4B
)
indicated that there were no significant publica
-tion bias in the included microarray datasets.
Additionally, a total of 364 liver cancer and 50
normal liver tissues from TCGA dataset were
analyzed. The 364 liver cancer tissues
consist-ed of 354 HCC and 10 non-HCC samples. The
precursor miR-145 level was significantly
down-regulated in liver cancer tissues including HCC
Table 2.
The result of the meta-analysis
based on the rest of 10 microarray datasets
ID I2 SMD P
GSE10694 73.0% -1.056 <0.001 GSE12717 80.3% -0.987 <0.001 GSE20077 79.6% -1.024 <0.001 GSE21362 79.3% -1.036 <0.001 GSE22058 56.4% -0.738 <0.001 GSE31383 77.3% -0.847 <0.001 GSE40744 78.8% -1.047 <0.001 GSE41874 78.3% -0.876 <0.001 GSE54751 80.3% -0.977 <0.001 GSE57555 80.2% -0.941 <0.001 GSE69580 79.7% -0.911 <0.001
and non-HCC (P<0.001,
Table
3
) and in 354 cases of HCC
(P<0.001,
Table 3
;
Figure 5A
)
when compared to
non-can-cerous liver tissues. Besides,
the ROC curve demonstrated
a significant diagnostic value
of miR-145 for HCC according
to the TCGA data (AUC=0.859,
P<0.001,
Table 3
;
Figure 5B
).
Based on the TCGA data, the
cut-off value was 12.76. The
sensitivity was 0.82 and the
specificity 0.76.
Suppressive role of
miR-145-5p expression in the progress
of HCC
To comprehend the clinical
significance of miR-145-5 in
the progress of HCC, the
rela-tionship between miR-145-5p
and available
clinicopathologi-cal parameters of HCC in GEO
datasets was explored. The
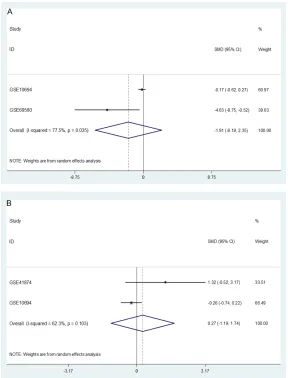
result exhibited that
miR-145-5p expressed significantly
hi-gher in younger group (<50
years old) than in older group
(≥50 years old) (SMD=-0.58,
95% CI: 0.21 to 0.95, P<0.002,
Figure 6A
). However, no
signif-icant difference was found
when gender, HBV infection,
liver cirrhosis and condition of
metastasis were considered
(all P>0.05,
Figures 6B
,
6C
,
7
).
Regarding to TCGA data, the
level of miR-145 was signifi
-cantly down-regulated in HBV
positive group when compar-
ed to HBV negative group
(P=0.047,
Table 3
). And the
patients with alcohol fatty li-
ver disease presented signi-
ficantly higher expression of
miR-145 than that without
(P=0.031,
Table 3
). However,
no apparent relationships we-
re noted between miR-145
expression and other
avail-Figure 3. Sensitivity analysis of the 11 microarray datasets.
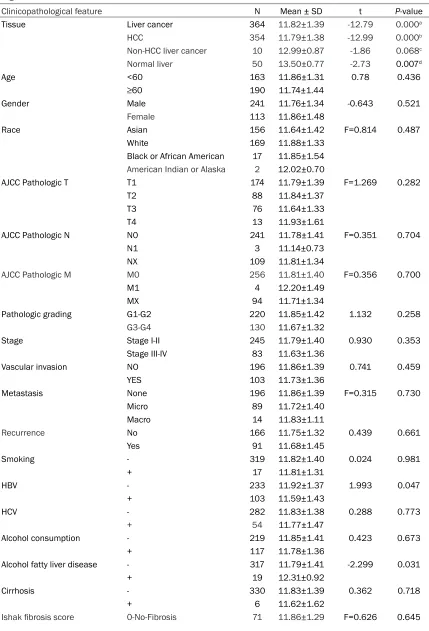
Table 3.
The correlation between miR-145-5p and the clinicopathological parameters of HCC
accord-ing to TCGA data
Clinicopathological feature N Mean ± SD t P-value
Tissue Liver cancer 364 11.82±1.39 -12.79 0.000a
HCC 354 11.79±1.38 -12.99 0.000b
Non-HCC liver cancer 10 12.99±0.87 -1.86 0.068c
Normal liver 50 13.50±0.77 -2.73 0.007d
Age <60 163 11.86±1.31 0.78 0.436
≥60 190 11.74±1.44
Gender Male 241 11.76±1.34 -0.643 0.521
Female 113 11.86±1.48
Race Asian 156 11.64±1.42 F=0.814 0.487
White 169 11.88±1.33
Black or African American 17 11.85±1.54
American Indian or Alaska 2 12.02±0.70
AJCC Pathologic T T1 174 11.79±1.39 F=1.269 0.282
T2 88 11.84±1.37
T3 76 11.64±1.33
T4 13 11.93±1.61
AJCC Pathologic N N0 241 11.78±1.41 F=0.351 0.704
N1 3 11.14±0.73
NX 109 11.81±1.34
AJCC Pathologic M M0 256 11.81±1.40 F=0.356 0.700
M1 4 12.20±1.49
MX 94 11.71±1.34
Pathologic grading G1-G2 220 11.85±1.42 1.132 0.258
G3-G4 130 11.67±1.32
Stage Stage I-II 245 11.79±1.40 0.930 0.353
Stage III-IV 83 11.63±1.36
Vascular invasion NO 196 11.86±1.39 0.741 0.459
YES 103 11.73±1.36
Metastasis None 196 11.86±1.39 F=0.315 0.730
Micro 89 11.72±1.40
Macro 14 11.83±1.11
Recurrence No 166 11.75±1.32 0.439 0.661
Yes 91 11.68±1.45
Smoking - 319 11.82±1.40 0.024 0.981
+ 17 11.81±1.31
HBV - 233 11.92±1.37 1.993 0.047
+ 103 11.59±1.43
HCV - 282 11.83±1.38 0.288 0.773
+ 54 11.77±1.47
Alcohol consumption - 219 11.85±1.41 0.423 0.673
+ 117 11.78±1.36
Alcohol fatty liver disease - 317 11.79±1.41 -2.299 0.031
+ 19 12.31±0.92
Cirrhosis - 330 11.83±1.39 0.362 0.718
+ 6 11.62±1.62
able clinicopathological features, including age,
gender, race, AJCC Pathologic TNM, Pathologic
grading, condition of vascular invasion,
condi-tion of metastasis, condicondi-tion of cirrhosis or HCV
infection (all P>0.05,
Table 3
).
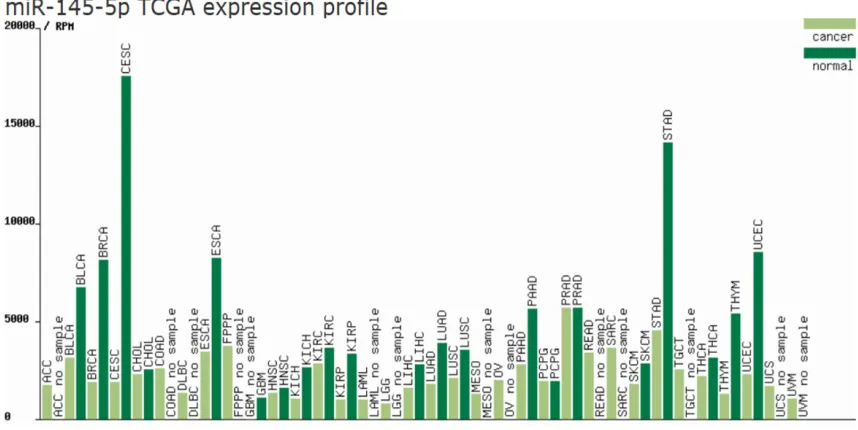
To gain the acquaintance of the expression
pat-tern of miR-145-5p in multiple diseases.
Human MicroRNA Expression Database (HMED)
was used to display the miRNA expression in
more than 400 data sets from smRNA data in
NCBI GEO and SRA (
Figure 8
, http://bioinfo.life.
hust.edu.cn/smallRNA/index.php), as well as
from TCGA (
Figure 9
, http://bioinfo.life.hust.
edu.cn/miR_path), containing samples from
various diseases and tissues.
Prospective signaling pathways of miR-145-5p
in HCC
Prediction of the target genes of miR-145-5p in
HCC: A total of 12 online prediction software
were used in current study to predict
prospec-tive target genes of miR-145-5p in HCC. Genes
which appeared at least 6 times as mentioned
above were selected for further study. Finally,
1434 predicted target genes of miR-145-5p
were obtained. Subsequently, we obtained the
final potential target genes by overlapping
1434 predicted target genes and 1800
HCC-related genes from NLP, which ended up with
144 genes.
GO analysis and KEGG pathway analysis
GO analysis and KEGG pathway analyses were
performed to explore the biological function of
144 target genes, and we found that most of
the genes were enriched in the pathways of
regulation of cell proliferation, enzyme linked
receptor protein signaling pathway and positiv
e
regulation of cell proliferation
according to
bio-logical processes (BP) (
Table 4
), and most of
1,2-Portal Fibrosis 30 12.16±1.393,4-Fibrous Speta 28 11.89±1.46
5-Nodular Formation and
incomplete Cirrhosis 8 12.16±1.44 6-Established Cirrhosis 70 11.74±1.27
Child pugh classification grade A 202 11.76±1.44 F=0.062 0.940
B 18 11.84±1.07
C 1 11.40
[image:8.612.91.522.222.423.2]a. Liver cancer VS Normal liver. b. HCC VS Normal liver. c. Non-HCC liver cancer VS Normal liver. d. HCC VS Non-HCC liver cancer.
the genes were related with enzyme binding,
[image:9.612.90.372.70.611.2]growth factor binding, protein kinase activity
receptor protein signaling pathway. As for KEGG
pathway analysis, pathways in cancer, chronic
Figure 6. Forest plots for evaluation of the correlation between miR-145-5p expression and different clinical pathological parameters in HCC performed
by fixed-effects models. A: Forest plots with fixed-effects model for the miR-145-5p level in the younger group (<50 years old) versus older group (≥50 years old). B: Forest plots with fixed-effects model for the miR-145-5p level in the male group versus female group. C: Forest plots with fixed-effects model
for the miR-145-5p level in the HBV(+) group versus HBV(-) group.
according to molecular
func-tion (MF) (
Table 4
), and as for
cell component (CC), the
tar-get genes were mainly enri-
ched in the terms of cell sur
-face, plasma membrane part,
plasma membrane (
Table 4
).
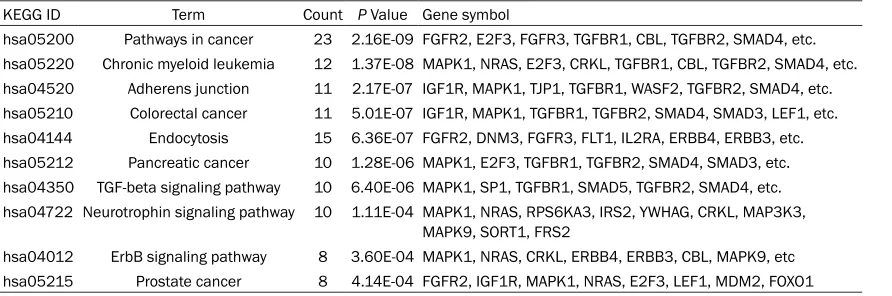
In addition, the KEGG pathway
analysis indicated the most
possible pathways of the 144
target genes including
path-ways in cancer, chronic
my-eloid leukemia and adherens
junction (
Table 5
).
The PPI network of the target
genes of miR-145-5p: The PPI
network of the 144 determ-
ined target genes of
miR-145-5p was performed by STRING
(
Figure 10
). And nine target
genes with the highest layout
degree were picked up from
the 144 genes, which
includ-ed SMAD2, SMAD3, FOXO1,
SMAD4, NRAS, MAPK1, SP1,
MAPK9 and CBL (
Figure 11
).
Discussion
In this study, we confirm that
obviously lower expression of
miR-145-5p can be achieved
in HCC tissues than that in
non-cancerous hepatic
tis-sues according to microarr-ay
and miRNA-seq data from
GEO and TCGA public
databas-es, which proves the potential
suppressive role of
miR-145-5p in H
CC as only identified by
real time RT-qPCR previously
[24-28, 30, 33, 53-56]. Fur-
thermore, 144 genes are
over-lapped from prediction and
NLP [8, 41, 42], which can be
regarded as prospective
tar-gets of miR-145-5p
specifi-cally in HCC. Via GO analysis,
the predicted 144 prospective
target genes are signi
ficantly
enriched in the pathways of
myeloid leukemia and adherens junction are
highlighted. Finally, nine hub genes (SMAD2,
SMAD3, FOXO1, SMAD4, NRAS, MAPK1, SP1,
MAPK9 and CBL) were identified to be most
essential potential target genes of miR-145-5p
in HCC.
The abnormal expression of miR-145-5p has
been reported in various malignant tumors. It
has been identified to execute the function as a
tumor suppressor in prostate cancer, gastric
cancer, lung cancer, Hodgkin lymphoma, colon
cancer and skin cancer [57-62]. However, Baici
et al [63] pointed out an opposite expression
pattern of miR-145-5p level being significantly
higher in non-melanoma skin cancer, which
[image:10.612.89.377.70.448.2]Due to the stability of miRNAs in
serum/plas-ma, also the convenience to be detected as a
non-invasive method in clinic, miRNAs have the
potentials to be novel circulating biomarkers for
the early screening of HCC [64-66]. Never-
theless, the diagnostic value of circulating
miR-145-5p was contradictory based on three
avail-able studies. The miScript miRNA PCR Array
was performed to examine the level of miR-145
in the plasma or serum with 23 HCC with HBV,
20 samples of liver cirrhosis, 20 samples of
chronic hepatitis B and 16 cases of healthy
controls. MiR-145-5p was elevated in the the
plasma or serum HCC as compared to other
groups [67]. But evident down-regulation of
miR-145-5p expression was observed in HCC
Figure 7. Forest plots for evaluation of the correlation between miR-145-5p expression and different clinical pathological parameters in HCC performed by random-effects models. A. Forest plot with random-effects model for the miR-145-5p level in the liver cirrhosis group versus non-cirrhosis group. B. Forest plot with random-effects model for the miR-145-5p level in the metas-tasis group versus non-metasmetas-tasis group.
suggests that distinct
expres-sion forms can be seen in
dif-ferent cancers. This
heteroge-neous expression is also
shown from HMED (
Figures 8
and
9
), which provides miRNA
expression in more than 400
datasets from smRNA data in
NCBI GEO, SRA and TCGA.
blood samples [53]. Similarly, expression of
miR-145-5p was related to a decreased risk of
HCC by other group [56]. The possibility for
cir-culating miR-145-5p as a diagnostic biomarker
in HCC still needs to be validated with larger
sample size.
The biological function of miR-145-5p has also
been noticed in HCC cells in vitro. Overexpr-
[image:11.612.93.521.78.303.2]ession of miR-145-5p by being transfected with
miRNA mimic in HCC cell lines clearly
sup-pressed cell proliferation, cell migration, as well
as the ability of invasion. Meanwhile,
miR-145-5p could also induce cell apoptosis [28, 30].
Furthermore, both of chemoresistance and the
process of EMT could be influenced by
miR-145-5p in HCC cells [31]. These evidences
assist to explain the clinicopathological value
Figure 8. The miR-145-5p expression in more than 400 data sets from smRNA data in NCBI GEO and SRA. The figure
was downloaded from http://bioinfo.life.hust.edu.cn/smallRNA/index.php.
[image:11.612.91.520.361.576.2]of miR-145-5p as observed from clinical
samples.
When concerning the molecular mechanism
and target genes of miR-145-5p in HCC, only
single target has been verified by different
groups with luciferase reporter assay, including
YTHDF2 [29], ROCK1 [30], FSCN1 [28], CUL5
[26], ADAM17 [33], OCT4-pg4 [34], IRS1 and
IRS2 [35]. Since some other target genes could
exist and have not yet been clarified, we per
[image:12.612.86.541.81.569.2]-formed the in silico investigation based on
pre-diction and NLP analysis. By overlapping the
predicted target genes from online platforms
and the abnormal expressed genes from NLP
analysis, we finally achieved a group of 144
genes, which could have high credibility to be
the prospective targets of miR-145-5p in HCC.
Further, bioinformatics analyses were
conduct-ed to dissect the relevant signal pathways and
molecular function of miR-145-5p in HCC.
According to the GO analysis, the target genes
Table 4.
Go functional annotation of most significant targets of miR-145-5p
GO ID Term Count P Value Gene symbol Biological process
GO:0042127 Regulation of cell proliferation 40 1.96E-17 FGFR2, E2F3, FGFR3, NRP1, ERBB4, ERBB3, SOX4, FOXO1, etc.
GO:0007167 Enzyme linked receptor protein signaling
pathway 25 4.76E-14 FGFR2, IRS2, NRP1, FGFR3, FLT1, ERBB4, ERBB3, TGFBR1, CBL, SMAD5, TGFBR2, SMAD4, SMAD3, FOXO1, SMAD2, PXN, etc.
GO:0008284 Positive regulation of cell proliferation 27 5.08E-14 FGFR2, E2F3, NRP1, FGFR3, ERBB4, SOX4, RPS6KB1, TGFB2, etc.
GO:0010604 Positive regulation of macromolecule
meta-bolic process 34 1.25E-11 E2F3, SOX4, FOXO1, SOX9, CITED2, TGFB2, HOXA1, IGF1R, H2AFX, TCF4, IRS2, IKZF1, TGFBR1, SMAD5, SMAD4, SMAD3, LEF1, etc. GO:0045935 Positive regulation of nucleobase, nucleoside,
nucleotide and nucleic acid metabolic process 28 9.85E-11 E2F3, SOX4, FOXO1, ABCA1, SOX9, CITED2, HOXA1, IGF1R, H2AFX, TCF4, IKZF1, TGFBR1, SMAD5, SMAD4, SMAD3, LEF1, etc. GO:0051173 Positive regulation of nitrogen compound
metabolic process
28 2.00E-10 E2F3, SOX4, FOXO1, ABCA1, SOX9, CITED2, HOXA1, IGF1R, H2AFX,
TCF4, IKZF1, TGFBR1, SMAD5, SMAD4, SMAD3, LEF1, etc.
GO:0009891 Positive regulation of biosynthetic process 29 2.18E-10 E2F3, SOX4, FOXO1, ABCA1, SOX9, CITED2, TGFB2, HOXA1, IGF1R,
TCF4, IRS2, IKZF1, TGFBR1, SMAD5, SMAD4, SMAD3, LEF1, etc. GO:0010557 Positive regulation of macromolecule
biosyn-thetic process 28 2.82E-10 E2F3, SOX4, FOXO1, SOX9, CITED2, TGFB2, HOXA1, IGF1R, TCF4, IRS2, IKZF1, TGFBR1, SMAD5, SMAD4, SMAD3, LEF1, SMAD2, etc. GO:0031328 Positive regulation of cellular biosynthetic
process 28 7.84E-10 E2F3, SOX4, FOXO1, ABCA1, SOX9, CITED2, HOXA1, IGF1R, TCF4, IRS2, IKZF1, TGFBR1, SMAD5, SMAD4, SMAD3, LEF1, SMAD2, etc.
GO:0051270 Regulation of cell motion 16 9.14E-10 IRS2, ADAM10, NRP1, FLT1, ERBB4, TGFBR1, SMAD3, RPS6KB1,
TGFB2, CITED2, IGF1R, MAPK1, ETS1, ADAM17, UNC5C, etc. Molecular function
GO:0019899 Enzyme binding 21 1.83E-07 FMNL2, IRS2, ADAM10, TGFBR1, TGFBR2, SMAD3, SMAD2, etc.
GO:0019838 Growth factor binding 10 8.98E-07 IGF1R, FLT1, IL2RA, ERBB3, CTGF, TGFBR1, TGFBR2, SORT1, etc.
GO:0004672 Protein kinase activity 21 1.86E-06 FGFR2, NRP1, FGFR3, FLT1, ERBB4, ERBB3, NUAK1, TGFBR1, etc.
GO:0003700 Transcription factor activity 27 2.50E-06 E2F3, SOX4, FOXO1, SOX9, CITED2, HOXA1, HEY1, TCF4, etc.
GO:0005160 Transforming growth factor beta receptor binding
5 1.94E-05 TGFBR1, TGFBR2, SMAD3, SMAD2, TGFB2
GO:0046983 Protein dimerization activity 18 2.30E-05 CLCN3, ACHE, ADAM10, IKZF1, ERBB3, TGFBR1, TGFBR2, etc.
GO:0004714 Transmembrane receptor protein tyrosine
kinase activity
7 4.98E-05 FGFR2, IGF1R, FLT1, NRP1, FGFR3, ERBB4, ERBB3
GO:0016563 Transcription activator activity 15 5.16E-05 E2F3, SMAD5, SMAD4, SMAD3, FOXO1, LEF1, SMAD2, HLTF, etc.
GO:0046332 SMAD binding 6 8.54E-05 TGFBR1, HIPK2, TGFBR2, SMAD4, SMAD3, SMAD2
GO:0030528 Transcription regulator activity 31 1.24E-04 E2F3, SOX4, FOXO1, SOX9, CITED2, HOXA1, HEY1, TCF4, etc.
Cell component
GO:0009986 Cell surface 16 7.05E-07 PVR, FGFR2, CLCN3, ACHE, ADAM10, IL2RA, TGFBR2, etc.
GO:0044459 Plasma membrane part 43 1.18E-06 CLCN3, ACHE, FGFR3, ERBB4, TLN2, ERBB3, SLC7A8, NEDD9, etc.
GO:0005886 Plasma membrane 59 5.43E-06 PVR, NRP1, TLN2, SLC7A8, RPS6KB1, SPRY4, CD47, CTGF, etc.
GO:0016323 Basolateral plasma membrane 11 1.80E-05 TJP1, ERBB4, TLN2, ERBB3, LASP1, P2RY2, TGFBR1, ADAM17, etc.
GO:0016324 Apical plasma membrane 9 3.12E-05 CLCN3, ERBB3, P2RY2, TGFBR1, ADAM17, CFTR, DPP4, etc.
GO:0005654 Nucleoplasm 21 1.54E-04 RAD23B, E2F3, SMAD5, SMAD4, SMAD3, BNIP3, FOXO1, etc.
GO:0045177 Apical part of cell 9 2.50E-04 CLCN3, ERBB3, P2RY2, TGFBR1, ADAM17, CFTR, DPP4, etc.
GO:0005667 Transcription factor complex 9 7.24E-04 E2F3, SMAD5, SMAD4, SMAD3, LEF1, SMAD2, DACH1, TCF4, etc.
GO:0031252 Cell leading edge 7 0.001740 TLN2, WASF2, ADAM17, NEDD9, CDK6, CDH2, PXN
were mainly contributed to HCC development
by regulating cell proliferation and affecting the
enzyme linked receptor protein signaling
path-way from BP terms and these key pathpath-ways
have already been reported in HCC [68, 69]. MF
analysis suggested that enzyme binding,
growth factor binding and protein kinase
activ-ity were the most important terms, consistent
with literatures [70]. CC analysis pointed out
that the target genes were mainly enriched in
cell surface, plasma membrane part and
plas-ma membrane, which demonstrated that a
membrane-related function could play a signifi
-cant role in HCC [71]. For KEGG pathways, the
target genes were closely related to many
clas-sical pathways of malignant tumors including
leukemia, colorectal cancer, pancreatic cancer
and prostate cancer, which suggested the
com-mon carcinogenic function of miR-145-5p in
multiple cancers and these pathways have also
been proved to be able to cause the incidence
and and accelerate the development of HCC
[72, 73].
Among the 144 obtained target genes of
miR-145
-5p for HCC, nine specific genes were
screened out by PPI network as hub genes for
miR-145-5p in HCC. Among these nine hub
genes, several have been documented to play
substantial roles in HCC and can act as target
genes of other miRNAs. Yang et al reported that
miR-101 weakened TGF-β signaling transduc
-tion via targeting SMAD2 in HCC [74]. Fu et al
proved that the up-regulation of SMAD3 led to
the down-regulation of Bcl-2, which thus
pro-moted the apoptosis of HCC cells [75]. Yu et al
found that miR-144 effected the invasion and
metastasis of HCC via regulating SMAD4 [76].
The study of Zeng et al revealed that miR-130a
directly target at FOXO1 to decelerate the
inva-sion and migration capacity of HCC cells [77];
and Xu et al indicted the regulation of FOXO1
induced abnormal proliferation of HCC cells
[78]. Keng et al found that the activation of
NRAS caused the hyperplasia of liver cells,
which might induce the occurrence of HCC [79].
Li et al concluded that MPAK1 enriched in
opi-oid signaling way to influence on
HCV-associated HCCs [80]. Zhao et al determined
that the lower expression of SP1 resulted in
poor prognosis of HCC [81]. Zhang et al also
confirmed that CBL had the potential to be a
biomarker to estimate the prognosis of HCC
[82]. To sum up, the nine hub genes are tightly
associated with HCC and may play important
roles in the development of HCC. And
miR-145-5p may target these nine genes and modulate
HCC cells via similar signal pathways which
have been clarified by the above studies. To
definite the direct targets of miR-145-5p and
understand its specific mechanism and related
pathways in HCC, further investigation is
necessary.
Conclusion
This study verifies that miR-145-5p is markedly
down-regulated in HCC tissues, suggesting that
the loss or down-regulation of miR-145-5p
expression could be a prospective molecular
mechanism triggering abnormal oncogenic
sig-naling in HCC, via targeting multiple pathways
and genes. The hub genes of SMAD2, SMAD3,
FOXO1, SMAD4, NRAS, MAPK1, SP1, MAPK9
and CBL may be the pivotal target genes of
miR-145-5p in HCC, which provides new space
for the HCC research.
Table 5.
KEGG pathway analysis of miR-145-5p targets genes
KEGG ID Term Count P Value Gene symbolhsa05200 Pathways in cancer 23 2.16E-09 FGFR2, E2F3, FGFR3, TGFBR1, CBL, TGFBR2, SMAD4, etc. hsa05220 Chronic myeloid leukemia 12 1.37E-08 MAPK1, NRAS, E2F3, CRKL, TGFBR1, CBL, TGFBR2, SMAD4, etc. hsa04520 Adherens junction 11 2.17E-07 IGF1R, MAPK1, TJP1, TGFBR1, WASF2, TGFBR2, SMAD4, etc. hsa05210 Colorectal cancer 11 5.01E-07 IGF1R, MAPK1, TGFBR1, TGFBR2, SMAD4, SMAD3, LEF1, etc. hsa04144 Endocytosis 15 6.36E-07 FGFR2, DNM3, FGFR3, FLT1, IL2RA, ERBB4, ERBB3, etc. hsa05212 Pancreatic cancer 10 1.28E-06 MAPK1, E2F3, TGFBR1, TGFBR2, SMAD4, SMAD3, etc. hsa04350 TGF-beta signaling pathway 10 6.40E-06 MAPK1, SP1, TGFBR1, SMAD5, TGFBR2, SMAD4, etc. hsa04722 Neurotrophin signaling pathway 10 1.11E-04 MAPK1, NRAS, RPS6KA3, IRS2, YWHAG, CRKL, MAP3K3,
MAPK9, SORT1, FRS2
Acknowledgements
The authors thank Prof. Gang Chen from the
First Affiliated Hospital of Guangxi Medical
University for providing genes generated by
natural language processing (NLP) analysis.
The authors also thank GEO and TCGA for the
high throughput data.
Disclosure of conflict of interest
None.
Address correspondence to: Haixin Huang, De-
partment of Medical Oncology, Fourth
[image:14.612.91.523.74.581.2]Affiliat-ed Hospital of Guangxi MAffiliat-edical University/Liu- zhou Worker’s Hospital, 1 Liushi Road, Liuzhou
545005, Guangxi Zhuang Autonomous Region, China. Tel: 3815405; Fax: 0086-772-3815405; E-mail: [email protected]
References
[1] Ye XD, Yuan Z, Zhang J and Yuan Z. Radiologi-cal biomarkers for assessing response to lo-coregional therapies in hepatocellular carci-noma: from morphological to functional imaging (Review). Oncol Rep 2017; 37: 1337-1346.
[2] Li K, Lan Y, Wang J and Liu L. Chimeric antigen receptor-engineered T cells for liver cancers, progress and obstacles. Tumour Biol 2017; 39: 1010428317692229.
[3] Wu W, Liu S, Liang Y, Zhou Z and Liu X. MiR-7 inhibits progression of hepatocarcinoma by targeting KLF-4 and promises a novel diagnos-tic biomarker. Cancer Cell Int 2017; 17: 31. [4] Fu S, Zhou RR, Li N, Huang Y and Fan XG.
Hep-atitis B virus X protein in liver tumor microenvi-ronment. Tumour Biol 2016; [Epub ahead of print].
[5] Ali MA, Lacin S, Abdel-Wahab R, Uemura M, Hassan M, Rashid A, Duda DG and Kaseb AO. Nonalcoholic steatohepatitis-related hepato-cellular carcinoma: is there a role for the an-drogen receptor pathway? Onco Targets Ther 2017; 10: 1403-1412.
[6] Zhang B, Han S, Feng B, Chu X, Chen L and Wang R. Hepatitis B virus X protein-mediated non-coding RNA aberrations in the develop-ment of human hepatocellular carcinoma. Exp Mol Med 2017; 49: e293.
[7] Gan TQ, Tang RX, He RQ, Dang YW, Xie Y and Chen G. Upregulated MiR-1269 in
hepatocel-lular carcinoma and its clinical significance. Int
J Clin Exp Med 2015; 8: 714-721.
[8] Zhang X, Tang W, Chen G, Ren F, Liang H, Dang Y and Rong M. An encapsulation of gene signa-tures for hepatocellular carcinoma, MicroR-NA-132 predicted target genes and the corre-sponding overlaps. PLoS One 2016; 11: e0159498.
[9] Berretta M, Cavaliere C, Alessandrini L, Stanzi-one B, Facchini G, Balestreri L, Perin T and Canzonieri V. Serum and tissue markers in he-patocellular carcinoma and cholangiocarcino-ma: clinical and prognostic implications. Onco-target 2017; 8: 14192-14220.
[10] Li H and Zhang L. Liver regeneration microenvi-ronment of hepatocellular carcinoma for pre-vention and therapy. Oncotarget 2017; 8: 1805-1813.
[11] Liu Z, Wang C, Jiao X, Zhao S, Liu X, Wang Y and Zhang J. miR-221 promotes growth and invasion of hepatocellular carcinoma cells by constitutive activation of NFkappaB. Am J Transl Res 2016; 8: 4764-4777.
[12] Xing H, Yan C, Cheng L, Wang N, Dai S, Yuan J, Lu W, Wang Z, Han J, Zheng Y and Yang T. Clin-ical application of protein induced by vitamin K antagonist-II as a biomarker in hepatocellular carcinoma. Tumour Biol 2016; [Epub ahead of print].
[13] Huang WT, Chen ZX, He RQ, Wu YZ, Yin SY, Li-ang XN, Chen G, YLi-ang H, Peng ZG and YLi-ang LH. Clinicopathological role of miR-30a-5p in hepa-tocellular carcinoma tissues and prediction of its function with bioinformatics analysis. Onco Targets Ther 2016; 9: 5061-5071.
[14] Yang F, Li QJ, Gong ZB, Zhou L, You N, Wang S, Li XL, Li JJ, An JZ, Wang DS, He Y and Dou KF. MicroRNA-34a targets Bcl-2 and sensitizes hu-man hepatocellular carcinoma cells to sorafenib treatment. Technol Cancer Res Treat 2014; 13: 77-86.
[15] Cao DD, Xu HL, Liu L, Zheng YF, Gao SF, Xu XM and Ge W. Thalidomide combined with thans-catheter artierial chemoembolzation for pri-mary hepatocellular carcinoma: a systematic review and meta-analysis. Oncotarget 2017; 8: 44976-44993.
[16] Lin J, Wu L, Bai X, Xie Y, Wang A, Zhang H, Yang X, Wan X, Lu X, Sang X and Zhao H. Combina-tion treatment including targeted therapy for advanced hepatocellular carcinoma. Oncotar-get 2016; 7: 71036-71051.
[image:15.612.96.283.76.265.2][17] He R, Yang L, Lin X, Chen X, Lin X, Wei F, Liang X, Luo Y, Wu Y, Gan T, Dang Y and Chen G. MiR-30a-5p suppresses cell growth and enhances apoptosis of hepatocellular carcinoma cells via
targeting AEG-1. Int J Clin Exp Pathol 2015; 8: 15632-15641.
[18] Zhang M, Li M, Li N, Zhang Z, Liu N, Han X, Liu Q and Liao C. miR-217 suppresses prolifera-tion, migraprolifera-tion, and invasion promoting apop-tosis via targeting MTDH in hepatocellular car-cinoma. Oncol Rep 2017; 37: 1772-1778. [19] Zhou X, Huang Z, Xu L, Zhu M, Zhang L, Zhang
H, Wang X, Li H, Zhu W, Shu Y and Liu P. A pan-el of 13-miRNA signature as a potential bio-marker for predicting survival in pancreatic cancer. Oncotarget 2016; 7: 69616-69624. [20] Wang W, Zhang L, Zheng K and Zhang X.
miR-17-5p promotes the growth of osteosarcoma in a BRCC2-dependent mechanism. Oncol Rep 2016; 35: 1473-1482.
[21] Tang R, Zhong T, Dang Y, Zhang X, Li P and Chen G. Association between downexpression of MiR-203 and poor prognosis in non-small cell lung cancer patients. Clin Transl Oncol 2016; 18: 360-368.
[22] Niyazi M, Pitea A, Mittelbronn M, Steinbach J, Sticht C, Zehentmayr F, Piehlmaier D, Zitzels-berger H, Ganswindt U, Rodel C, Lauber K, Belka C and Unger K. A 4-miRNA signature pre-dicts the therapeutic outcome of glioblastoma. Oncotarget 2016; 7: 45764-45775.
[23] Tang R, Liang L, Luo D, Feng Z, Huang Q, He R, Gan T, Yang L and Chen G. Downregulation of MiR-30a is associated with poor prognosis in lung cancer. Med Sci Monit 2015; 21: 2514-2520.
[24] Lupini L, Pepe F, Ferracin M, Braconi C, Calle-gari E, Pagotto S, Spizzo R, Zagatti B, Lanuti P, Fornari F, Ghasemi R, Mariani-Costantini R, Bolondi L, Gramantieri L, Calin GA, Sabbioni S, Visone R, Veronese A, Negrini M. Over-expres-sion of the 483-3p overcomes the miR-145/TP53 pro-apoptotic loop in hepatocellular carcinoma. Oncotarget 2016; 7: 31361-31371. [25] Zhang L, Xiang ZL, Zeng ZC, Fan J, Tang ZY and
Zhao XM. A microRNA-based prediction model for lymph node metastasis in hepatocellular carcinoma. Oncotarget 2016; 7: 3587-3598. [26] Gao F, Sun X, Wang L, Tang S and Yan C.
Down-regulation of MicroRNA-145 Caused by hepati-tis B virus X protein promotes expression of CUL5 and contributes to pathogenesis of hep-atitis B virus-associated hepatocellular carci-noma. Cell Physiol Biochem 2015; 37: 1547-1559.
[27] Shi KQ, Lin Z, Chen XJ, Song M, Wang YQ, Cai YJ, Yang NB, Zheng MH, Dong JZ, Zhang L and Chen YP. Hepatocellular carcinoma associat- ed microRNA expression signature: integrat- ed bioinformatics analysis, experimental
vali-dation and clinical significance. Oncotarget
2015; 6: 25093-25108.
[28] Wang G, Zhu S, Gu Y, Chen Q, Liu X and Fu H. MicroRNA-145 and MicroRNA-133a inhibited proliferation, migration, and invasion, while promoted apoptosis in hepatocellular carcino-ma cells via targeting FSCN1. Dig Dis Sci 2015; 60: 3044-3052.
[29] Yang Z, Li J, Feng G, Gao S, Wang Y, Zhang S, Liu Y, Ye L, Li Y and Zhang X. MicroRNA-145 modulates N6-methyladenosine levels by tar-geting the 3’-untranslated mRNA region of the N6-methyladenosine binding YTH domain fam-ily 2 protein. J Biol Chem 2017; 292: 3614-3623.
[30] Ding W, Tan H, Zhao C, Li X, Li Z, Jiang C, Zhang Y and Wang L. MiR-145 suppresses cell prolif-eration and motility by inhibiting ROCK1 in he-patocellular carcinoma. Tumour Biol 2016; 37: 6255-6260.
[31] Ju BL, Chen YB, Zhang WY, Yu CH, Zhu DQ and Jin J. miR-145 regulates chemoresistance in hepatocellular carcinoma via epithelial mesen-chymal transition. Cell Mol Biol 2015; 61: 12-16.
[32] Wang Y, Hu C, Cheng J, Chen B, Ke Q, Lv Z, Wu J and Zhou Y. MicroRNA-145 suppresses hepa-tocellular carcinoma by targeting IRS1 and its downstream Akt signaling. Biochem Biophys Res Commun 2014; 446: 1255-1260.
[33] Yang XW, Zhang LJ, Huang XH, Chen LZ, Su Q, Zeng WT, Li W and Wang Q. miR-145 suppress-es cell invasion in hepatocellular carcinoma cells: miR-145 targets ADAM17. Hepatol Res 2014; 44: 551-559.
[34] Wang L, Guo ZY, Zhang R, Xin B, Chen R, Zhao J, Wang T, Wen WH, Jia LT, Yao LB and Yang AG. Pseudogene OCT4-pg4 functions as a natural micro RNA sponge to regulate OCT4 expres-sion by competing for miR-145 in hepatocellu-lar carcinoma. Carcinogenesis 2013; 34: 1773-1781.
[35] Law PT, Ching AK, Chan AW, Wong QW, Wong CK, To KF and Wong N. MiR-145 modulates multiple components of the insulin-like growth factor pathway in hepatocellular carcinoma. Carcinogenesis 2012; 33: 1134-1141.
[36] Dweep H, Gretz N and Sticht C. miRWalk data-base for miRNA-target interactions. Methods Mol Biol 2014; 1182: 289-305.
[37] Xiong DD, Lv J, Wei KL, Feng ZB, Chen JT, Liu KC, Chen G and Luo DZ. A nine-miRNA signa-ture as a potential diagnostic marker for breast carcinoma: An integrated study of 1,110 cas-es. Oncol Rep 2017; 37: 3297-3304.
[38] Zeng JH, Xiong DD, Pang YY, Zhang Y, Tang RX,
Luo DZ and Chen G. Identification of molecular
[39] Gan TQ, Xie ZC, Tang RX, Zhang TT, Li DY, Li ZY and Chen G. Clinical value of miR-145-5p in NSCLC and potential molecular mechanism ex-ploration: a retrospective study based on GEO, qRT-PCR, and TCGA data. Tumour Biol 2017; 39: 1010428317691683.
[40] Wang D, Zhu ZM, Tu YL, Dou CQ, Xu Y, Tan XL, Han MM, Yang ZJ, Jin X, Zhang B, Cai S and Liu
ZW. Identfication of key miRNAs in pancreatitis
using bioinformatics analysis of microarray data. Mol Med Rep 2016; 14: 5451-5460. [41] Zhang X, Ye ZH, Liang HW, Ren FH, Li P, Dang
YW and Chen G. Down-regulation of miR-146a-5p and its potential targets in hepatocellular carcinoma validated by a TCGA- and GEO-based study. FEBS Open Bio 2017; 7: 504-521.
[42] Huang WT, Wang HL, Yang H, Ren FH, Luo YH, Huang CQ, Liang YY, Liang HW, Chen G and Dang YW. Lower expressed miR-198 and its potential targets in hepatocellular carcinoma: a clinicopathological and in silico study. Onco-Targets Ther 2016; 9: 5163-5180.
[43] Mou T, Zhu D, Wei X, Li T, Zheng D, Pu J, Guo Z
and Wu Z. Identification and interaction analy -sis of key genes and microRNAs in hepatocel-lular carcinoma by bioinformatics analysis. World J Surg Oncol 2017; 15: 63.
[44] Xu X and Li H. Integrated microRNAgene analy-sis of coronary artery disease based on miRNA
and gene expression profiles. Mol Med Rep
2016; 13: 3063-3073.
[45] Wang DW, Yu SY, Cao Y, Yang L, Liu W, Er XQ,
Yao GJ and Bi ZG. Identification of CD20, ECM,
and ITGA as biomarkers for osteosarcoma by integrating transcriptome analysis. Med Sci Monit 2016; 22: 2075-2085.
[46] Sun J, Zhang L, He Y, Zhang K, Wu L, Fan Y and Xie Z. To unveil the molecular mechanisms of Qi and blood through systems biology-based investigation into Si-Jun-Zi-Tang and Si-Wu-Tang formulae. Sci Rep 2016; 6: 34328. [47] Yan H, Li Z, Shen Q, Wang Q, Tian J, Jiang Q and
Gao L. Aberrant expression of cell cycle and material metabolism related genes contrib-utes to hepatocellular carcinoma occurrence. Pathol Res Pract 2017; 213: 316-321.
[48] Liu P, Jiang W, Zhou S, Gao J and Zhang H. Combined analysis of ChIP sequencing and gene expression dataset in breast cancer. Pathol Oncol Res 2017; 23: 361-368.
[49] Cao L, Zhang Q, Cheng S, Chen Z, Hua Z, Yang J, Liu D and Cui N. Long non-coding RNAs and genes contributing to the generation of cancer stem cells in hepatocellular carcinoma
identi-fied by RNA sequencing analysis. Oncol Rep
2016; 36: 2619-2624.
[50] Gan S, Qiu S, Feng Y, Zhang Y, Qian Q, Wan Z
and Tang J. Identification of genes associated
with the effect of inflammation on the neuro -transmission of vascular smooth muscle cell. Exp Ther Med 2017; 13: 1303-1312.
[51] Wang Y. Identifying key stage-specific genes
and transcription factors for gastric cancer based on RNA-sequencing data. Medicine (Baltimore) 2017; 96: e5691.
[52] Wu YS, Chen YT, Bao YT, Li ZM, Zhou XJ, He JN,
Dai SJ and Li CY. Identification and verification
of potential therapeutic target genes in berber-ine-treated Zucker diabetic fatty rats through bioinformatics analysis. PLoS One 2016; 11: e0166378.
[53] Bandopadhyay M, Banerjee A, Sarkar N, Pani-grahi R, Datta S, Pal A, Singh SP, Biswas A, Chakrabarti S and Chakravarty R. Tumor sup-pressor micro RNA miR-145 and onco micro RNAs miR-21 and miR-222 expressions are differentially modulated by hepatitis B virus X protein in malignant hepatocytes. BMC Cancer 2014; 14: 721.
[54] Karakatsanis A, Papaconstantinou I, Gazouli M, Lyberopoulou A, Polymeneas G and Voros D. Expression of microRNAs, miR-21, miR-31, 122, 145, 146a, 200c, miR-221, miR-222, and miR-223 in patients with hepatocellular carcinoma or intrahepatic
chol-angiocarcinoma and its prognostic signifi -cance. Mol Carcinog 2013; 52: 297-303. [55] Gao P, Wong CC, Tung EK, Lee JM, Wong CM
and Ng IO. Deregulation of microRNA expres-sion occurs early and accumulates in early stages of HBV-associated multistep hepatocar-cinogenesis. J Hepatol 2011; 54: 1177-1184. [56] Wang C, Hann HW, Ye Z, Hann RS, Wan S, Ye X,
Block PD, Li B, Myers RE, Wang X, Juon HS, Ci-van J, Chang M, Bae HS, Xing J and Yang H. Prospective evidence of a circulating microR-NA signature as a non-invasive marker of he-patocellular carcinoma in HBV patients. Onco-target 2016; [Epub ahead of print]
[57] Wang C, Tao W, Ni S, Chen Q, Zhao Z, Ma L, Fu Y and Jiao Z. Tumor-suppressive microRNA-145 induces growth arrest by targeting SENP1 in human prostate cancer cells. Cancer Sci 2015; 106: 375-382.
[58] Wang W, Ji G, Xiao X, Chen X, Qin WW, Yang F, Li YF, Fan LN, Xi WJ, Huo Y, Wen WH, Yang AG and Wang T. Epigenetically regulated miR-145 suppresses colon cancer invasion and metas-tasis by targeting LASP1. Oncotarget 2016; 7: 68674-68687.
[59] Ren K, Li Z, Li Y, Zhang W and Han X. Long non-coding RNA taurine-upregulated gene 1 pro-motes cell proliferation and invasion in gastric cancer via negatively modulating miRNA-145-5p. Oncol Res 2017; 25: 789-798. [60] Mataki H, Seki N, Mizuno K, Nohata N,
and Inoue H. Dual-strand tumor-suppressor microRNA-145 (miR-145-5p and miR-145-3p) coordinately targeted MTDH in lung squamous cell carcinoma. Oncotarget 2016; 7: 72084-72098.
[61] Zhang Y, Wen X, Hu XL, Cheng LZ, Yu JY and Wei ZB. Downregulation of miR-145-5p corre-lates with poor prognosis in gastric cancer. Eur Rev Med Pharmacol Sci 2016; 20: 3026-3030.
[62] Paydas S, Acikalin A, Ergin M, Celik H, Yavuz B
and Tanriverdi K. Micro-RNA (miRNA) profile in
Hodgkin lymphoma: association between clini-cal and pathologiclini-cal variables. Med Oncol 2016; 33: 34.
[63] Balci S, Ayaz L, Gorur A, Yildirim Yaroglu H, Ak-bayir S, Dogruer Unal N, Bulut B, Tursen U and
Tamer L. microRNA profiling for early detection
of nonmelanoma skin cancer. Clin Exp Derma-tol 2016; 41: 346-351.
[64] Huang YH, Liang KH, Chien RN, Hu TH, Lin KH, Hsu CW, Lin CL, Pan TL, Ke PY and Yeh CT. A circulating MicroRNA signature capable of as-sessing the risk of hepatocellular carcinoma in cirrhotic patients. Sci Rep 2017; 7: 523. [65] Reichl P and Mikulits W. Accuracy of novel
di-agnostic biomarkers for hepatocellular carci-noma: an update for clinicians (Review). Oncol Rep 2016; 36: 613-625.
[66] Mao B and Wang G. MicroRNAs involved with hepatocellular carcinoma (Review). Oncol Rep 2015; 34: 2811-2820.
[67] Wang Y, Gao Y, Shi W, Zhai D, Rao Q, Jia X, Liu
J, Jiao X and Du Z. Profiles of differential ex -pression of circulating microRNAs in hepatitis B virus-positive small hepatocellular carcino-ma. Cancer Biomark 2015; 15: 171-180. [68] Xu WH, Zhang JB, Dang Z, Li X, Zhou T, Liu J,
Wang DS, Song WJ and Dou KF. Long non-cod-ing RNA URHC regulates cell proliferation and apoptosis via ZAK through the ERK/MAPK sig-naling pathway in hepatocellular carcinoma. Int J Biol Sci 2014; 10: 664-676.
[69] Wang J, Su H, Han X and Xu K. Inhibition of fi -broblast growth factor receptor signaling im-pairs metastasis of hepatocellular carcinoma. Tumour Biol 2014; 35: 11005-11011.
[70] Zhao S, Li H, Wang Q, Su C, Wang G, Song H, Zhao L, Luan Z and Su R. The role of c-Src in the invasion and metastasis of hepatocellular carcinoma cells induced by association of cell surface GRP78 with activated alpha2M. BMC Cancer 2015; 15: 389.
[71] Kondoh N, Wakatsuki T, Ryo A, Hada A, Aihara T, Horiuchi S, Goseki N, Matsubara O, Takena-ka K, Shichita M, TanaTakena-ka K, Shuda M and
Ya-mamoto M. Identification and characterization
of genes associated with human hepatocellu-lar carcinogenesis. Cancer Res 1999; 59: 4990-4996.
[72] Sun CG, Cao XJ, Zhou C, Liu LJ, Feng FB, Liu RJ, Zhuang J and Li YJ. Construction of
gene/pro-tein interaction networks for primary myelofi -brosis and KEGG pathway-enrichment analysis of molecular compounds. Genet Mol Res 2015; 14: 16126-16132.
[73] Yang J, Chen L, Kong X, Huang T and Cai YD. Analysis of tumor suppressor genes based on gene ontology and the KEGG pathway. PLoS One 2014; 9: e107202.
[74] Yang J, Lu Y, Lin YY, Zheng ZY, Fang JH, He S and Zhuang SM. Vascular mimicry formation is promoted by paracrine TGF-beta and SDF1 of
cancer-associated fibroblasts and inhibited by
miR-101 in hepatocellular carcinoma. Cancer Lett 2016; 383: 18-27.
[75] Fu QH, Zhang Q, Zhang JY, Sun X, Lou Y, Li GG, Chen ZL, Bai XL and Liang TB. LB-100 sensi-tizes hepatocellular carcinoma cells to the ef-fects of sorafenib during hypoxia by activation of Smad3 phosphorylation. Tumour Biol 2016; 37: 7277-7286.
[76] Yu M, Lin Y, Zhou Y, Jin H, Hou B, Wu Z, Li Z, Jian Z and Sun J. MiR-144 suppresses cell pro-liferation, migration, and invasion in hepato-cellular carcinoma by targeting SMAD4. Onco-Targets Ther 2016; 9: 4705-4714.
[77] Zeng YB, Liang XH, Zhang GX, Jiang N, Zhang T, Huang JY, Zhang L and Zeng XC. miRNA-135a promotes hepatocellular carcinoma cell migra-tion and invasion by targeting forkhead box O1. Cancer Cell Int 2016; 16: 63.
[78] Lou K, Chen N, Li Z, Zhang B, Wang X, Chen Y, Xu H, Wang D and Wang H. MicroRNA-142-5p overexpression inhibits cell growth and induc-es apoptosis by regulating FOXO in hepatocel-lular carcinoma cells. Oncol Res 2017; 25: 65-73.
[79] Keng VW, Tschida BR, Bell JB and Largaespada DA. Modeling hepatitis B virus X-induced hepa-tocellular carcinoma in mice with the Sleeping Beauty transposon system. Hepatology 2011; 53: 781-790.
[80] Li YL, Zheng MX and Wang G. A personalized
approach identifies disturbed pathways and
key genes in hepatitis C virus-cirrhosis with he-patocellular carcinoma. Eur Rev Med Pharma-col Sci 2016; 20: 4266-4273.
[81] Zhao N, Li S, Wang R, Xiao M, Meng Y, Zeng C, Fang JH, Yang J and Zhuang SM. Expression of microRNA-195 is transactivated by Sp1 but in-hibited by histone deacetylase 3 in hepatocel-lular carcinoma cells. Biochim Biophys Acta 2016; 1859: 933-942.