CIRCULATING CATECHOLAMINES AND SWIMMING
PERFORMANCE IN THE ATLANTIC COD, GADUS MORHUA
BY P. J. BUTLER*, M. AXELSSON, F. EHRENSTROM, J. D. METCALFEt AND S. NILSSON
Department of Zoophysiology, University of Goteborg, PO Box 250 59, S-400 31 Goteborg, Sweden
Accepted 4 July 1988
Summary
Sectioning the first four pairs of spinal nerves prevents the large increase in circulating catecholamine concentrations seen in Atlantic cod swimming at their critical velocity (Ucnt)- There is also a significant reduction in the swimming performance of the fish. To test whether this reduced performance results from the lack of increase in plasma catecholamine levels or from the fact that other organs are also denervated by the operative procedure, a mixture of adrenaline and noradrenaline was infused into swimming, denervated fish. This caused a significant increase in their Ucrjt- It is concluded, therefore, that the rise in plasma
catecholamine levels seen in Atlantic cod swimming at their maximum sustainable velocity enhances the swimming performance of these fish.
Introduction
There are a number of ways in which circulating catecholamines could enhance the supply of oxygen to the active muscles in exercising fish and thus improve the animals' swimming performance. In resting rainbow trout in well-aerated water, approximately 40% of secondary lamellae are not perfused with blood (Booth, 1978) and, even in those lamellae that are perfused, up to 20% of the blood may be in basal channels which are probably nonrespiratory (Tuurala et al. 1984). Adrenaline increases interlamellar recruitment in vivo and in vitro (Booth, 1979; Holbert et al. 1979) and an increase in perfusion pressure (which could be mediated by catecholamines) causes inter- and intralamellar recruitment in vitro (Fairell et al. 1979,1980). During intralamellar recruitment a greater proportion of blood perfuses the central lamellar spaces, i.e. there is a move away from the basal channels. There may, therefore, be a thinning of the epithelium of the lamellae, thus reducing the diffusion distance between blood and water. It has also
* Permanent address: Department of Zoology and Comparative Physiology, University of Birmingham, Birmingham B15 2TT, UK.
t Permanent address: Directorate of Fisheries Research, Fisheries Laboratory, Ministry of Agriculture, Fisheries and Food, Lowestoft, Suffolk NR33 OHT, UK.
378 P. J. BUTLER AND OTHERS
been suggested that adrenaline increases the permeability of the gills to oxygen (Isaia, 1984), although direct experimental evidence is lacking.
There are, therefore, a number of mechanisms by which plasma catecholamines could enhance oxygen transfer across the gills. In fact, addition of adrenaline to the perfusate does increase the difference in the partial pressures of oxygen between arterial and venous blood (Pa^—PvoJ (Wood et al. 1978) and oxygen uptake (Perry et al. 1985) in isolated, head preparations of rainbow trout perfused with saline. It has recently been demonstrated that there are modest increases in the levels of plasma catecholamines in rainbow trout swimming to apparent exhaustion (Butler et al. 1986). However, as these authors point out, the concentration of adrenaline is at least an order of magnitude lower than that used in the in vitro studies mentioned above. It may not, of course, be justifiable to extrapolate from in vitro observations to the whole animal, as any number of other mechanisms in the whole animal could influence the effect of catecholamines (Butler etal. 1986).
It is known that environmental hypoxia and damage to the gills can reduce swimming performance in fish (Jones, 1971; Bushnell et al. 1984; Duthie & Hughes, 1987). So, if plasma catecholamines do play an important role in providing an increased oxygen supply to the active muscles during exercise, particularly by enhancing gas exchange, then preventing their release should impair aerobic swimming performance.
The object of the present study was to test this possibility in the Atlantic cod,
Gadus morhua, by determining the effect of surgically preventing the release of
catecholamines on the swimming performance of the fish. In case the operation had other effects, in addition to preventing the release of catecholamines, swimming performance was also determined in these fish while infusing a mixture of adrenaline and noradrenaline.
Materials and methods
Data were obtained from 25 Atlantic cod, Gadus morhua, of either sex and with a body mass of 380-920g and length of 35-46cm. They were captured off the Swedish west coast, kept in recirculated, aerated sea water at 10-12°C in the aquarium of the Zoological Institute in Goteborg and were used within 1 week of capture.
The fish were anaesthetized initially in tricaine methane sulphonate (Sigma) (100mgI"1) in sea water until ventilatory movements ceased. They were then transferred to an operating table and sea water containing the anaesthetic (approximately SOmgl"1) was continuously pumped over the gills during the operation.
response to 'stress'. In the remaining fish (sham-operated fish) incisions were made in the skin and muscles, and spinal nerves 1-4 were identified but not sectioned. In all fish a catheter (PE50) was implanted into the afferent artery of the third gill arch for the measurement of ventral aortic blood pressure and heart rate and for the sampling of blood. The catheter was passed through the upper part of the operculum and sutured to the skin. It was filled with heparinized (approximately 50i.u. ml"1) 0-9% NaCl and attached during the experiment to a Statham P23 pressure transducer connected via a three-way tap to a Grass polygraph recorder. In some fish that had undergone denervation of the head kidney, a polyethylene catheter (i.d. 0-4 mm, o.d. 0-8 mm) was introduced into the gonadial vein for the infusion of catecholamines.
Fish were allowed to recover from the operation for at least 24 h before any experiment, during which time the effects of surgery and anaesthesia wore off and the cardiovascular variables stabilized (see Smith etal. 1985). Some fish recovered in holding tanks whereas others recovered in a Blazka-type water channel (see Axelsson & Nilsson, 1986, for details). All fish were transferred to the water channel at least 3 h before the commencement of an experiment to give them time to recover from the stress of handling. The water in the experimental tube was fully aerated and continuously replaced at a rate of 21min~1 from the main seawater system. The temperature was 10-12°C during all experiments.
In an initial series of experiments, critical swimming velocity, U ^ , (Brett, 1964) was determined for 17 intact fish (i.e. fish that had undergone no operative procedure) as described by Butler et al. (1986). Ucrit was uncorrected for the
presence of the fish in the channel (see Jones et al. 1974, for a discussion). In nine of these fish, the head kidneys were subsequently denervated and the remaining eight underwent sham operations. Their Ucrit was determined again, but this time
heart rate and ventral aortic blood pressure were monitored continuously and 0-6 ml of blood was removed from the fish when at rest, before swimming and at Ucnt (see Butler et al. 1986, for details). This mixed venous blood was analysed for partial pressure and content of oxygen (PvO2 and CvO2, respectively), pH (pHv),
haematocrit, and levels of lactate, plasma adrenaline and noradrenaline.
In a subsequent series of experiments, the head kidney was denervated in eight fish. Heart rate and ventral aortic blood pressure were monitored and blood samples (0-45 ml) were taken from the fish at rest and while swimming at Ucrit. In
four fish a mixture of adrenaline and noradrenaline (15-20 jxmoll"1 adrenaline, 10/imoir1 noradrenaline in saline) was continually infused, by a precision pump (Braun Limited), into the gonadial vein after lOmin of swimming in an attempt to simulate the levels reached in sham-operated fish swimming at U ^ . The fish were allowed 3 h to recover before being made to swim again while saline was infused into the gonadial vein. In the other four fish, saline was infused during the first swimming bout and the catecholamines were infused during the second bout.
380 P. J. BUTLER AND OTHERS
during the early stages of the swimming bout, swimming performance was reduced. Thus, infusion was at a low rate (4Julmin~1) initially and then increased
progressively (to 20/zlmin"1 and then to 40/xlmin"1) so that the desired level of cat6cholamines was reached towards the end of the swimming bout. The timing of the change from one infusion rate to another was dependent on the assumed swimming ability of the fish (which was easier to estimate when saline was infused first).
Ventral aortic blood pressure was measured with a Statham P23 pressure transducer connected to a Grass polygraph recorder system model 7D. Mean pressure was determined as diastolic+ one-third pulse pressure. Heart rate was obtained from the pulsatile pressure signal via a Grass 7P44 tachograph unit. Partial pressures of oxygen in water and prebranchial (venous) blood and pH (pH v) in the blood were determined by an appropriate Radiometer electrode and a PHM73 blood gas analyser. Oxygen content in the blood was measured by the method described by Tucker (1967), using distilled water at 0°C equilibrated with O2 for calibration, and haematocrit was determined with a Wifug
microhaematoc-rit centrifuge. Plasma lactate was assayed enzymatically using a Sigma kit (no. 826-UV) and a Perkin-Elmer Lambda 3 spectrophotometer.
The levels of adrenaline and noradrenaline in plasma were measured using ion-pair, reversed-phase high performance liquid chromatography (HPLC) with electrochemical detection. The sample preparation and analysis were performed basically as described by Ehrenstrom & Johansson (1985, 1987). Blood samples (200/xl) were immediately added to 20/zl of a mixture of 0-2moll"1 EGTA and 0-2moll"1 glutathione and centrifuged. The plasma was removed and stored at —20°C for no longer than 1 week. Such storage has no effect on the levels of catecholamines (Butler et al. 1978). The plasma samples (40-150jd) were deproteinized by addition of 0-2moll"1 perchloric acid (PCA) to give a total volume of 1-15 ml. After addition of an internal standard (3,4-dihydroxybenzyl-amine hydrobromide, DHBA; 600pg in 50[A of 0-1 moll"1 PCA), the samples were vortex-mixed for a few seconds and centrifuged in a Sigma model 3K-2 centrifuge at 4280g for lOmin at 4°C. The supernatant (1-0 ml) was transferred to tubes containing 10 mg of acid-washed alumina (chromatographic alumina, Sigma no. A-1772), 25 //I of 10 % EDTA and 25 /il of 1-5 % glutathione. After addition of 600fA of Tris buffer (2-0moll"1, pH8-6), the samples were vortex-mixed and the catecholamines allowed to adsorb onto the alumina for 30 min in a rotary mixer. The supernatant was aspirated off, the alumina was washed five times with 1 ml of chilled, deionized and filtered (0-45 pan membrane filters, Millipore) water and the catecholamines were eluted with 100/zl of 0 1 moll"1 PCA. The eluates were transferred to 0-25 ml polyethylene tubes (Eppendorf), capped and stored at —20°C until analysis by HPLC. Recoveries of standard catecholamine samples treated in the same fashion were estimated (70-85 %) and subsequent calculations were corrected for losses in the purification procedure.
detector (model M460). The electrochemical cell was a Bioanalytical systems dual glassy carbon cell with a Ag/AgCl reference electrode (model RE-1). A Rheodyne injection valve (model 7125) equipped with a 20/il loop was used for injection of samples onto the HPLC column. This consisted of a Supelco 7-5 cm x 4-6 mm analytical column (Supelcosil, 3/xm RP C18 silica particles). The
mobile phase consisted of 93-3 mmol I"1 sodium acetate, ll-34mmol P1 citric acid, 70mgP11-heptanesulphonic acid and lOmgl"1 EDTA(pH5-25). The separation of adrenaline and noradrenaline was completed in less than 4min at a flow rate of l-Smlmin"1. The output from the detector was displayed on a Shimadzu data processor (Chromatopac model C-R3A). This system was sufficiently sensitive to detect 0-1-0-5 x l O ~9m o l P1 catecholamines (1-5 pg per injection) using 40-150/d of plasma.
Mean values are expressed ±S.E. of the mean. The Wilcoxon's sign rank test for paired samples or the Mann-Whitney U-test were used (both as two-tailed tests, except where indicated) and the level of significance was set to P< 0-05.
Results
The effects of denervating the head kidney on the measured variables at rest, while swimming at U ^ i and upon Ucrit itself are given in Table 1. With the
exception of plasma lactate concentration, with was significantly lower (61 %) in the denervated group, none of the measured variables in the resting fish was affected by the denervation. When swimming at Ucrit, there were in both groups
significant decreases in mixed venous pH, PO2 and CO2 and significant increases in
heart rate, mean ventral aortic blood pressure, haematocrit, and concentrations of plasma lactate and plasma catecholamines. The values of most of these variables at Ucnt were similar for both the groups. However, plasma lactate concentration at Ucrit was significantly lower (50 %) in the denervated group, although it increased to three times the resting value in this group of fish compared with an increase to 2-3 times the resting value in the sham-operated group.
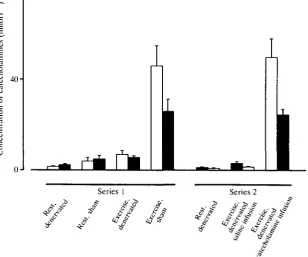
There were substantially larger differences between the two groups in the levels of plasma catecholamines at Ucrit (Fig- !)• Plasma adrenaline concentration in the sham-operated fish at Ucrit was 6-7 times greater than that in the denervated fish. It rose to 11-5 times the resting value in sham-operated fish and to 4-3 times the resting value in the denervated group. Plasma noradrenaline concentration at U ^ , was 4-6 times higher in the sham-operated fish than in the denervated group. It increased to 5-2 times the resting value in the former and to two times the resting value in the latter.
382 P . J. BUTLER A N D OTHERS
Table 1. Physiological variables in Atlantic cod, Gadus morhua, at rest and while
swimming at their critical swimming velocity (Ucril)
Sham-operated fish (N = 8) Denervated fish (N = 9) Rest Uni, Rest Ucn,
Heart rate (beats min"1) Mean ventral aortic blood
pressure (kPa) Mixed venous Po2 (kPa)
Mixed venous oxygen content (mmoir1) Mixed venous pH Haematocrit (%) Plasma [lactate]
43 ± 2-4 3-9 ±0-18
5-4 ±0-24 0-9 ±0-13
7-78 ± 0-032 14-3 ±0-65 2-6 ±0-56 4-0 ±1-55 5-0 ±1-48 21-6 ±0-26 54 ±1-0* 5-1 ±0-26* 4-0 ±0-38* 0-5 ±0-11* 7-50 ±0-038* 18-5 ±1-51*
60 ± 0-67*
460 ±8-85* 25-8 ±5-46* 21-7 ±0-42 43-6 ±2-39 37-7 ±1-81 87 ±4-4 605 ±62
39-5 ± 1-20
41 ±2-3 3-5 ±0-13 5-4 ±0-35 0-8 ±0-12 7-75 ±0-044 13-3 ±0-67 l-0±0-21t 1-6 ±0-30 2-3 ±0-53 21-7 ±0-24 51 ±1-7* 4-6 ±0-20* 3-7 ±0-32* 0-4 ±0-08* 7-63 ±0-055* 14-9 ±0-89* 3-0±0-92*t 6-9±l-77*t 5-6±0-75*t 21-5 ±0-36 41-9 ±1-44 31-5 ±2-03 75±4-7f 558 ±38
38-7 ± 0-82 Plasma [adrenaline]
( x l O -9m o i rl) Plasma [noradrenaline]
( x l O - ' m o i r1)
Po, of inspired water (kPa^ UOT, before operation (B)
(cms"1)
UCTJ, after operation (A)
(cms"1)
A/BxlO0(%)
Mass (g) Length (cm)
In nine fish the first four pairs of spinal nerves were sectioned to denervate the head kidney (denervated fish) and in eight fish the nerves were identified but not sectioned (sham-operated fish).
* Indicates a mean value significantly different from that for the same variable at rest within the same group.
t Indicates a mean value significantly different from that for the same variable in the sham-operated group.
between the two groups when using a two-tailed test, it was with a one-tailed test, and the ratio between U ^ after and Ucrit before the operation was significantly
lower in the denervated group of fish (when using a two-tailed test).
catechol-8 0 i
40-o U
0J
[image:7.451.68.377.93.350.2]Series 1 Series 2
Fig. 1. Levels of plasma adrenaline (open columns) and noradrenaline (filled columns) in Atlantic cod, Gadus morhua. In series 1 the head kidney of the fish was denervated in nine fish (denervated) whereas in eight animals the nerves were identified but not sectioned (sham). Data were obtained in fish at rest or when swimming at critical velocity, U ^ , , (exercise). In series 2 the head kidney was denervated in all (eight) fish which were infused with saline or catecholamines while swimming. When infusing with catecholamines the objective was to obtain plasma concentrations similar to those recorded in sham-operated fish swimming at U^i,. Values are means + S.E.
amines during both bouts of swimming. (Note that haematocrit did not increase at Ucrit during either of the swimming bouts.) The concentrations of plasma lactate at
Ucrjt were similar to that seen in denervated fish during the first series of
experiments. During both bouts of swimming, plasma lactate level was approxi-mately 1-5 times the resting value at UCnt- During saline infusion, both adrenaline
and noradrenaline concentrations at U ^ i were 2-1 times the resting values, whereas during catecholamine infusion they were, at U ^ , 38 times and 22 times the resting values, respectively (Fig. 1). The actual concentrations reached at U ^ , during catecholamine infusions were similar to those recorded in sham-operated fish swimming at U ^ (see Table 1).
384
P. J. BUTLER AND OTHERSTable 2. The effects of infusing either saline or a mixture of the catecholamines
adrenaline and noradrenaline (for details see text) on a number of physiological variables and critical swimming velocity (Ucrit) in eight Atlantic cod, Gadus
morhua, in which the first four pairs of spinal nerves were sectioned to denervate the
head kidney
Heart rate (beats min ') Mean ventral aortic blood
pressure (kPa) Mixed venous pH Haematocrit (%) Plasma [lactate]
Plasma [adrenaline] ( x l O - ' m o i r1) Plasma [noradrenaline]
(xHT'moir
1)
Ucrit Mass (g) Length (cm) Rest Before saline infusion 38 ±1-5 3-9 ±0-34 7-70 ±0-022 16-3 ±1-06 1-9 ±0-19 1-4 ±0-32 0-7 ±0-12 Before catecholamine infusion 38 ±1-8 3-8 ±0-16
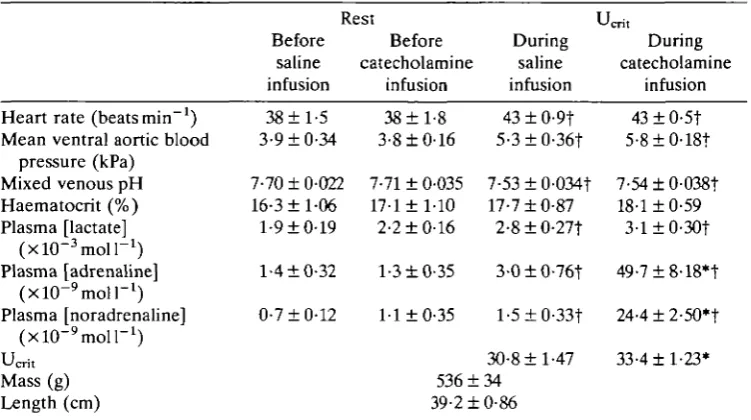
7-71 ± 0-035 17-1 ±1-10 2-2 ±0-16 1-3 ±0-35 1-1 ±0-35 536 ± 39-2 ± I During saline infusion
43 ± 0-9t 5-3±0-36t 7-53±0-034t 17-7 ±0-87 2-8±0-27f 3-0±0-76t l-5±0-33t 30-8 ±1-47 34 0-86 Jcrit During catecholamine infusion
43 ± 0-5t 5-8±0-18t 7-54±0-038t 18-1 ±0-59 3-l±0-30t 49-7±8-18*t 24-4±2-50*t 33-4 ±1-23*
•Indicates a mean value significantly different from that for the same variable during (or before) saline infusion for fish at rest or when swimming at U^f
t Indicates a mean value significantly different from that for the same variable in resting fish before the infusion of saline or catecholamines.
infused with saline (Table 2) was similar to that in the denervated fish without infusion (Table 1).
The swimming performance (i.e. U^O of the former group of fish was slightly (8%) but significantly greater when they were infused with catecholamines than when they were infused with saline.
Discussion
catecholamines in denervated fish significantly increased their U ^ , indicates that these hormones enhance the swimming performance of these animals. The data only indicate a marginal improvement, but in a number of the fish there was a noticeable improvement in the 'style' of swimming. Instead of the frequency of falling back to the grid increasing with time, it actually decreased for a short while after commencement of the highest rate of infusion of the catecholamines.
Of course, we have no idea which of the many possible mechanisms are the most important. Adrenergic vasomotor fibres, and not circulating catecholamines, are thought to be important in elevating blood pressure during exercise in cod (Axelsson & Nilsson, 1986). This proposal is supported by the present observation that the rise in ventral aortic pressure during swimming persists after denervation of the head kidney. Release from nerve terminals could also explain the significant rise in plasma catecholamine levels in swimming fish after denervation of the head kidney. Also, the rise in cardiac output during exercise in the cod is not impaired by total removal of autonomic influences (Axelsson, 1988). Unfortunately, it was not possible to obtain arterial blood samples, so we have no data on oxygenation of the arterial blood in denervated fish, and we did not measure intracellular pH (pHi) of the red blood corpuscles (RBCs). An interesting observation, which warrants further study, was that if the levels of plasma catecholamines were abnormally high at moderate swimming velocities, swimming was inhibited. In view of the findings of Smith et al. (1985) and Axelsson & Nilsson (1986), this inhibition is not likely to result from vasoconstriction in the locomotory muscles. It is known that both adrenaline and noradrenaline cause vasodilatation in the branchial vasculature of cod, with adrenaline being the more potent (Wahlqvist, 1980). This /3-adrenoceptor dilatation of afferent lamellar arterioles is probably mediated entirely by circulating catecholamines (Nilsson & Pettersson, 1981) which, together with <*-adrenoceptor constriction of efferent lamellar arterioles, may enhance oxygen transfer across the gills (Pettersson, 1983). Pettersson (1983) concluded that a'-adrenoceptor stimulation is the more potent in this respect, and it can be selectively mediated by the nervous system (Nilsson & Pettersson, 1981). The denervation procedure used in the present study would, therefore, have eliminated this effect.
There is evidence to suggest that the ability of rainbow trout and salmon to perform aerobic exercise after burst (anaerobic) swimming is not compromised because the elevated plasma catecholamine levels produced during the burst swimming serve to maintain pHi of the RBCs in the face of plasma acidosis (Primmett et al. 1986; Randall et al. 1987). As there are increases in both plasma catecholamine levels and plasma acidosis in cod swimming at Ucrit, this
phenom-enon could be an important component of the overall effect of circulating catecholamines on swimming performance in this fish.
386 P. J. BUTLER AND OTHERS
References
AXELSSON, M. (1988). The importance of nervous and humoral mechanisms in the control of cardiac performance in the Atlantic cod Gadus morhua at rest and during non-exhaustive exercise. J. exp. Biol. 137, 287-301.
AXELSSON, M. & NILSSON, S. (1986). Blood pressure control during exercise in the Atlantic cod, Gadus morhua. J. exp. Biol. 126, 225-236.
BOOTH, J. H. (1978). The distribution of blood flow in the gills of fish: application of a new technique to rainbow trout (Salmo gairdncri). J. exp. Biol. 73, 119-129.
BOOTH, J. H. (1979). The effects of oxygen supply, epinephrine, and acetylcholine on the distribution of blood flow in trout gills. J. exp. Biol. 83, 31-39.
BRETT, J. R. (1964). The respiratory metabolism and swimming performance of young sockeye salmon. J. Fish. Res. Bd Can. 21, 1183-1226.
BUSHNELL, P. G., STEFFENSEN, J. F. & JOHANSEN, K. (1984). Oxygen consumption and swimming performance in hypoxia-acclimated rainbow trout, Salmo gairdneri. J. exp. Biol.
113, 225-235.
BUTLER, P. J., METCALFE, J. D. & GINLEY, S. A. (1986). Plasma catecholamines in the lesser spotted dogfish and rainbow trout at rest and during different levels of exercise. J. exp. Biol.
123, 409-421.
BUTLER, P. J., TAYLOR, E. W., CAPRA, M. F. & DAVISON, W. (1978). The effect of hypoxia on the levels of circulating catecholamines in the dogfish Scyliorhinus canicula. J. comp. Physiol.
127, 325-330.
DUTHIE, G. G. & HUGHES, G. M. (1987). The effects of reduced gill area and hyperoxia on the oxygen consumption and swimming speed of rainbow trout. J. exp. Biol. 127, 349-354.
EHRENSTROM, F. & JOHANSSON, P. (1985). A method for very rapid determinations of catechols using ion-pairing reverse phase HPLC with electrochemical detection: effects of L-dopa treatment on the catechol content in various rat brain structures. Life Sci. 36, 867-879. EHRENSTROM, F. & JOHANSSON, P. (1987). Orcadian rhythms and contents of catechols in
different brain structures, peripheral organs and plasma of the Atlantic cod, Gadus morhua. Comp. Biochem. Physiol. 87C, 193-202.
FARRELL, A. P., DAXBOECK, C. & RANDALL, D. J. (1979). The effect of input pressure and flow on the pattern and resistance to flow in the isolated perfused gill of a teleost fish. /. comp. Physiol. 133, 233-240.
FARRELL, A. P., SOBIN, S. S., RANDALL, D. J. & CROSBY, S. (1980). Intralamellar blood flow patterns in fish gills. Am. J. Physiol. 239, R428-R436.
HOLBERT, P. W., BOLAND, E. J. & OLSON, K. R. (1979). The effect of epinephrine and acetylcholine on the distribution of red cells within the gills of the Channel catfish (Ictalurus punctatus). J. exp. Biol. 79, 135-146.
ISAIA, J. (1984). Water and nonelectrolyte permeation. In Fish Physiology, vol. XB (ed. W. S. Hoar & D. J. Randall), pp. 1-38. New York: Academic Press.
JONES, D. R. (1971). The effect of hypoxia and anaemia on the swimming performance of rainbow trout (Salmo gairdneri). J. exp. Biol. 55, 541-551.
JONES, D. R., KICENIUK, J. W. & BAMFORD, O. S. (1974). Evaluation of the swimming performance of several fish species from the Mackenzie river. /. Fish Res. Bd Can. 31, 1641-1647.
NILSSON, S., ABRAHAMSSON, T. & GROVE, D. J. (1976). Sympathetic nervous control of adrenaline release from the head kidney of the cod, Gadus morhua. Comp. Biochem. Physiol. 55C, 123-127.
NILSSON, S. & PETTERSSON, K. (1981). Sympathetic nervous control of blood flow in the gills of the Atlantic cod, Gadus morhua. J. comp. Physiol. 144, 157-163.
PERRY, S. F., DAXBOECK, C. & DOBSON, G. P. (1985). The effect of perfusion flow rate and adrenergic stimulation on oxygen transfer in the isolated, saline-perfused head of rainbow trout (Salmo gairdneri). J. exp. Biol. 116,251-269.
PETTERSSON, K. (1983). Adrenergic control of oxygen transfer in perfused gills of the cod, Gadus morhua. J. exp. Biol. 102, 327-335.
catecholamines in erythrocyte pH regulation and oxygen transport in rainbow trout (Salmo
gairdneri) during exercise. /. exp. Biol. 122, 139-148.
RANDALL, D. J., MENSE, D. & BOUTIUER, R. G. (1987). The effects of burst swimming on aerobic swimming in Chinook salmon (Oncorhynchus tshawytscha). Mar. Behav. Physiol. 13, 77-88.
SMITH, D. G., NILSSON, S., WAHLQVIST, I. & ERIKSSON, B.-M. (1985). Nervous control of blood pressure in the Atlantic cod, Gadus morhua. J. exp. Biol. 117, 335-347.
TUCKER, V. A. (1967). Method for oxygen content and dissociation curves on microliter blood
samples. J. appl. Physiol. 23, 410-414.
TUURALA, H., PART, P., NIKTNMAA, M. & SOIVIO, A. (1984). The basal channels of secondary lamellae in Salmo gairdneri gills - a non-respiratory shunt. Comp. Biochem. Physiol. 79, 35-39.
WAHLQVIST, I. (1980). Effects of catecholamines on isolated systemic and branchial vascular beds of the cod, Gadus morhua. J. comp. Physiol. 137, 139-143.