ABSTRACT
Proteorhodopsins are retinal-binding membrane proteins that may act as light-driven
proton pumps to generate energy that can be used for metabolism and growth. These proteins
were originally characterized in prokaryotes, but have now been found in many eukaryotic
marine species including one of the ocean’s most important primary producers, the pennate
diatom Pseudo-nitzschia granii. The precise function of proteorhodopsins in P. granii is
unknown. Evidence suggests that these proteins are expressed in response to iron limitation to
function in the formation of ATP when the photosynthetic machinery, which is highly dependent
upon iron availability, is not providing enough energy for the cell. The goal of my research is to
elucidate the function of these proteins in diatoms by examining the relative expression levels of
the rhodopsin and carotenoid oxygenase genes in response to iron and light limitation. My results
suggest that proteorhodopsins do serve as an alternative to photosynthesis for the generation of
ATP under iron limitation and that they may also serve to supplement energy production by
photosynthesis under light limitation. This research will provide valuable insight into these
diatoms’ mechanisms of coping with a common source of environmental stress.
INTRODUCTION
A class of proteins called bacteriorhodopsins were discovered in 1971 in the membrane
of the archaeon Halobacterium salinarum. These proteins are retinal-containing membrane
proteins which are able to transport protons across a membrane by undergoing a series of
conformational changes fueled by absorbing light (Beja et al. 2001). These proteins were
believed to only exist in halophilic archaea for some time. However, it was soon discovered that
exist in native marine bacteria taken from the surface waters of Monterey Bay (Beja et al 2001).
This means that heterotrophic bacteria in the ocean are not only capable of producing energy by
their normal means of ingesting the necessary nutrients, but could also be able to harness the
energy of light by using rhodopsins. These findings have led to more extensive research of these
proteins and their possible functions in both prokaryotic and, more recently, eukaryotic
photosynthetic organisms.
Another recent study showed that xanthorhodopsin, a member of the rhodopsin family of
proteins, was present in the eubacterium Salinibacter ruber. This study also demonstrated that
this xanthorhodopsin complex contained both retinal proteins and antenna carotenoid pigment.
This carotenoid pigment could be important to the rhodopsin complex for several reasons, most
notably that it would increase the absorbance of light at different spectra from the retinal
proteins. By transferring the energy gained from light absorption to the retinal proteins, these
pigments could further facilitate the pumping of protons in an unfavorable direction across a
membrane to store energy (Baloshov et al. 2005).
Most recently, a study from 2011 found that some marine eukaryotic organisms,
including dinoflagellates and diatoms, have proteorhodopsins present within their membranes
which were acquired through lateral gene transfer from prokaryotic bacteria (Slamovitz et al.
2011). The proteorhodopsin protein complex in eukaryotic organisms consists of a
transmembrane opsin subunit which passes through the membrane seven times in an α-helical
configuration. This opsin subunit is bound to a complex of retinal and other proteins which are
used to absorb light (Giuliano et al. 2003). In diatoms, proteorhodopsins are most likely located
in the plastid membranes since the proteorhodopsin gene is similar to other pigments and
These findings raise the question as to what advantages eukaryotic photosynthetic organisms
would gain by expressing proteorhodopsins. They already contain the photosynthetic apparatus
which uses light energy to generate a proton gradient, which is then used to store energy in the
form of ATP. Why would they need another protein complex that performs essentially the same
task? Evidence suggests that expressing proteorhodopsins could pose an advantage for diatoms
in iron-limited environments since proteorhodopsin expression was elevated within iron-limited
natural phytoplankton communities (Marchetti et al. 2012).
This hypothesis is also backed by the fact that many of the proteins involved in carrying
out photosynthesis require iron to function efficiently. Conversely, proteorhodopsins require
small amounts of iron to function properly. Retinal proteins, which are formed from the cleavage
of β-carotene, are one of the major components of the light absorbing complexes found in
proteorhodopsins. The iron requirement of the rhodopsin protein complexes comes from
carotenoid oxygenases, the enzymes that cleave β-carotene (Giuliano et al. 2003). This
requirement is still much lower than the iron requirements of the photosynthetic machinery,
which lends further evidence to the hypothesis that proteorhodopsins could function as
light-driven proton pumps in iron-limited environments. Furthermore, phytoplankton have been shown
to adjust their methods of converting light to usable energy based on the availability of nutrients
and metals such as iron. All of this provides support to this hypothesis that marine photosynthetic
organisms, such as diatoms, would benefit from expressing proteorhodopsins since they could
provide an alternative mechanism to photosynthesis for energy generation in extremely
iron-limited environments (Marchetti et al. 2012).
Photosynthetic organisms could benefit from expressing proteorhodopsins for other
been shown that the maximum absorbance of proteorhodopsins can be shifted from green to blue
light with the substitution of just one amino acid. Furthermore, proteorhodopsins are capable of
absorbing light at different wavelengths from other protein complexes involved in
photosynthesis (Fuhrman et al. 2008). Since the availability of certain wavelengths of light can
vary with geographic location, oceanic depth, and other factors, being able to harness many
different forms of light into usable energy would provide a huge advantage for these organisms.
Proteorhodopsins could also benefit photosynthetic organisms through their ability to act as
sensory proteins. By providing extra sensitivity to light levels, these proteins could provide a
“clock” by which the photosynthetic organism could carry out different biological processes
which were specific to the time of day at which they are necessary (Fuhrman et al. 2008). When
the potential advantages of these other functions of proteorhodopsins are combined with the
advantage these proteins could provide by providing an alternative mechanism for the synthesis
of ATP in severely iron-limited environments, these proteins could provide a huge evolutionary
advantage to photosynthetic organisms.
METHODS
Cultures of the pennate diatom Pseudo-nitzschia granii were grown under four different
treatments: iron-replete with normal light exposure (abbreviated Fe+, HL), iron-replete with low
light exposure (Fe+, LL), iron-deficient with normal light exposure (Fe-, HL), and iron-deficient
with low light exposure (Fe-, LL). Normal light treatments were exposed to light levels of 150
µmol photons m-2 s-1, while low light treatments were exposed to light levels of 60 µmol photons
Medium for these cultures was made using AQUIL, an artificial sea water medium. This
medium was double-chelexed to remove all trace metals in a trace metal clean (TMC) facility.
This double-chelexed medium was then microwaved and placed in 2-liter acid-washed, Q-H2
O-rinsed, autoclaved polycarbonate bottles, where it was left to cool overnight. A mixture of 50 µL
of DFB and 11.07 µL of 1.4 ∗ 10−4 M Fe per liter was also prepared and left overnight to ensure
adequate mixing. After cooling, ethylenediaminetetraacetic acid (EDTA)-trace metals (minus
iron) and vitamins (B12, thiamine, and biotin) were added according to AQUIL medium
concentrations (Price et al. 1989). Premixed Fe-EDTA (1:1) was added to the iron-replete
medium to achieve iron concentrations of 10-19 mol L-1 (used for “Fe+, HL” and “Fe+, LL”
cultures). The previously prepared Fe-DFB mixture was added to the iron-limited medium (used
for “Fe-, HL” and “Fe-, LL” cultures).
In vivo fluorescence of each culture was measured regularly using a 10-AU Fluorometer
(Turner Designs) in order to monitor the growth of the diatoms in each culture. Fluorescence
increases with increasing amounts of chlorophyll a, which corresponds to an increase in cell
growth and division in P. granii, making this an accurate method to measure growth rates of
these diatoms. Upon reaching fluorescence values greater than 50.0, cultures were transferred to
keep the diatoms within their exponential phase of growth. Transfers consisted of placing 100.0
µL of the old culture into a new, clean tube containing fresh medium to prevent the diatoms from
exhausting the nutrient supplies.
Using the measured fluorescence values, growth rates were calculated by plotting the
natural logarithm of the fluorescence values against time for each culture. Assuming a strong
linear correlation (r2 ≥ 0.98), growth rates were equated to the slope of the line of best fit of these
of growth rates. Growth rates of the different treatments were compared to verify that P. granii
growth is in fact limited by both iron and light availability. Fv/Fm measurements were made
using a FIRe fluorometer to measure the photosynthetic efficiency of each culture. This data was
used in conjunction with the calculated growth rates to further show the physiological effects of
iron and light limitation on P. granii.
Once the small cultures were acclimated to their treatment conditions, large two liter
cultures were grown in duplicate for each treatment. These cultures were filtered at the late
exponential phase of their growth. Multiple filters were used for each culture. Some filters from
each culture were shipped to Dr. Brian Hopkinson at the University of Georgia for protein
expression analysis using a proteorhodopsin-specific antibody. The RNA was extracted from the
remaining filters of each culture using an RNAqueous-4PCR RNA isolation kit (Ambion). After
determining the RNA concentration of each sample, the RNA samples were DNased and then
reverse transcribed using a SuperScript III First Strand Synthesis kit (Invitrogen). The cDNA
generated from reverse transcription was then used in quantitative-PCR (q-PCR) using primers
specific to the proteorhodopsin, carotenoid oxygenase, and actin genes in P. granii to determine
the levels of expression of the proteorhodopsin and carotenoid oxygenase genes relative to actin
in the transcriptomes of each sample. The comparative CT method as described by Schmittgen
and Livak (2008) was used to measure the expression of proteorhodopsin and carotenoid
oxygenase relative to actin in each sample, and these ratios were compared to determine the
relative expression levels of the rhodopsin gene within each treatment.
Iron limitation and, to a lesser extent, light limitation both impacted the growth rates and
photosynthetic efficiencies (measured as Fv/Fm) of P. granii, which was expected based on
previous studies. Using two sample t-tests, the growth rates of all treatments were significantly
different from one another at a 95% confidence interval. Therefore, both iron deficient and light
deficient environments caused the growth of P. granii to be significantly limited. The
photosynthetic efficiencies of the diatoms under each treatment were slightly different.
Iron-replete treatments had significantly higher photosynthetic efficiencies than iron-limited
treatments at 95% confidence. However, high light treatments were not significantly different
from low light treatments with regard to photosynthetic efficiency, signifying that the availability
of light has very little impact on photosynthetic efficiency. This suggests that iron plays a much
larger role in altering the physiological responses of these diatoms to their environment than
light, although light still has a noticeable effect on the growth rates of the diatoms within iron
treatments. The growth rates and photosynthetic efficiencies of the larger cultures appear to
follow the same trend.
Transcription of the rhodopsin gene in P. granii was substantially up-regulated in
iron-limited treatments. Under high light, rhodopsin expression increased approximately ten-fold in
iron-limited treatments relative to iron-replete treatments, and the expression levels relative to
iron status were about 60 times higher in low light treatments. The rhodopsin gene was also
up-regulated slightly in low light environments compared to high light environments. An
approximate five-fold increase in rhodopsin expression was observed between low light and high
light treatments in iron-limited treatments, but there was no noticeable difference in expression
have a much more significant effect on the expression of the rhodopsin gene in P. granii
compared to light limitation, although both factors have a noticeable effect.
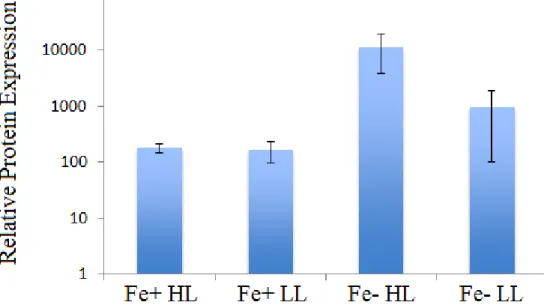
The results of the protein expression analysis also show that proteorhodopsin expression
is up-regulated significantly under iron limitation. However, the up-regulation under low light
observed in the relative transcript expression of the rhodopsin gene is not consistent with the
protein expression data. Under iron-replete conditions, light limitation showed no significant
effect on the expression of the proteorhodopsin protein, and under iron limitation, the protein
was expressed more highly under high light than low light.
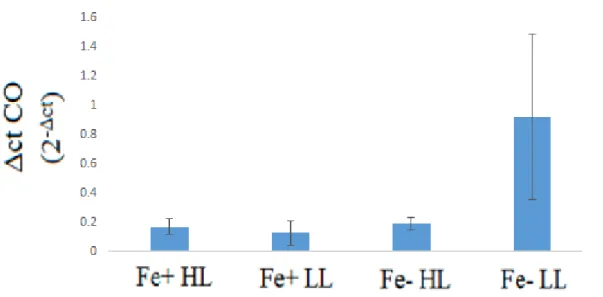
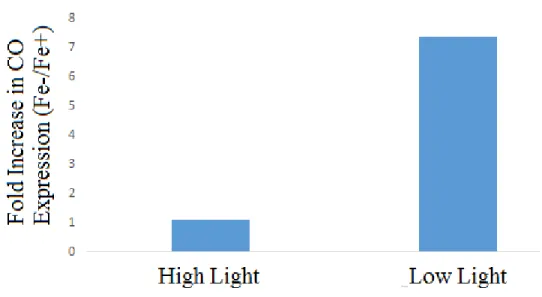
Expression of the carotenoid oxygenase gene was significantly increased only under both
iron and light limitation. There was no significant difference in expression of this gene between
the other three treatments. As a result, there was no difference in expression between light
treatments under iron replete treatments, but an approximate five-fold increase in expression was
observed in the low light treatments relative to the high light treatments when iron was limiting.
Furthermore, under high light, there was no noticeable difference in expression between iron
treatments. However, there was an approximately seven-fold increase in expression in the
Figure 2. Photosynthetic efficiencies of small maintenance cultures and large cultures.
Figure 3. Relative levels of gene expression of rhodopsins in P. granii. Note that Δct represents the difference in the cycle threshold values of rhodopsin and actin for each treatment.
Figure 5. Rhodopsin transcript expression relative to light status.
Figure 7. Relative levels of gene expression of carotenoid oxygenase. Note the Δct CO represents the expression of carotenoid oxygenase relative to actin.
Figure 9. Carotenoid oxygenase transcript expression relative to iron status.
DISCUSSION
P. granii grown under iron-limiting conditions substantially up-regulated the expression
of both the rhodopsin gene and protein relative to P. granii grown under iron-replete conditions.
Additionally, higher levels of transcript expression were observed under light limitation,
although this trend was not supported by the protein expression analysis. However, the
up-regulation of both the gene and the protein suggests that proteorhodopsins in diatoms could serve
as an alternative mechanism of ATP synthesis when photosynthesis is inhibited by limited iron
availability.
The relative expression levels of the carotenoid oxygenase gene provide further support
for this hypothesis. A significant up-regulation in the expression of this gene was only observed
in the iron and light limited treatment. This suggests that the diatoms are generating a larger
amount of proteorhodopsin-associated pigments in these conditions. If proteorhodopsins are
selective advantage to these diatoms under iron and light limitation since these pigments would
increase the proportion of available light energy that could be converted to cellular energy by
absorbing more light in the green and blue wavelengths. In other words, since these pigments
absorb light energy at different wavelengths from those involved in photosynthesis (primarily
chlorophyll a and b), expressing more of these pigments would allow the diatoms to utilize more
of the available light energy for the production of ATP. It would also be advantageous as it
would allow for greater energy production through the proteorhodopsins when energy production
from photosynthesis is decreased due to limited iron availability.
Because proteorhodopsins lack a source of reductant that can be used in carbon fixation
(Fuhrman et al. 2008), it is unlikely that proteorhodopsins could replace photosynthesis as the
primary means of generating enough energy for growth and division in the diatom species that
express them. However, given the above evidence, it is likely that these proteins can act as
light-driven proton pumps and can provide an additional source of energy under iron limitation, which
could be critical for the survival of these diatoms during periods of iron limitation. Ultimately,
this means that these diatoms are capable of surviving in a broader range of conditions than
previously believed, which could have large implications on our perception of global primary
productivity and nutrient cycling, particularly in chronically iron-limited regions of the world’s
REFERENCES
1. Beja, O., Spudich, E. N., Spudich, J. L., Leclerc, M., DeLong, E. F. (2001). Proteorhodopsin
phototrophy in the ocean. Nature411, 786-789.
2. Balashov, S. P., Imasheva, E. S., Boichenko, V. A., Anton, J., Wang, J. M. (2005).
Xanthorhodopsin: A proton pump with a light harvesting carotenoid antenna. Science
309, 2061-2064.
3. Slamovits, C. H., Okamoto, N., Burri, L., James, E. R., Keeling, P. J. (2011). A bacterial
proteorhodopsin proton pump in marine eukaryotes. Nature Communications 2(183), 1-6.
4. Giuliano, G., Al-Babili, S., Lintig, J.V. (2003). Carotenoid oxygenases: cleave it or leave it.
Trends in Plant Science8, 145-149.
5. Scala, S., and Bowler, C. (2001). Molecular insights into the novel aspects of diatom biology.
Cellular and Molecular Life Sciences58, 1666-1673.
6. Marchetti, A., Schruth, D. M., Durkin, C. A., Parker, M. S., Kodner, R. B., Berthiaume, C. T.,
Morales, R., Allen, A. E., Armbrust, E. V. (2012). Comparative metatranscriptomics
identifies molecular bases for the physiological responses of phytoplankton to varying
iron availability. Proceedings of the National Academy of Sciences of the United States of
America109(6), E317-E325.
7. Fuhrman, J.A., Schwalbach, M.S., Stingl, U. (2008). Proteorhodopsins: an array of
8. Price N. M., Harrison, G. I., Hering, J. G., Hudson, R. J., Nirel, P. M., Palenik, B., Morel, F.,
M. (1989). Preparation and chemistry of the artificial algal culture medium Aquil.
Biological Oceanography6(5), 443–461.
9. Schmittgen, T.D., and Livak, K.J. (2008) Analyzing real-time PCR data by the comparative