0095-1137/04/$08.00⫹0 DOI: 10.1128/JCM.42.11.5327–5331.2004
Copyright © 2004, American Society for Microbiology. All Rights Reserved.
Molecular Detection of Linezolid Resistance in
Enterococcus faecium
and
Enterococcus faecalis
by Use of 5
⬘
Nuclease Real-Time PCR
Compared to a Modified Classical Approach
Guido Werner,* Birgit Strommenger, Ingo Klare, and Wolfgang Witte
Robert Koch Institute, Wernigerode Branch, Wernigerode, Germany
Received 2 March 2004/Returned for modification 27 May 2004/Accepted 5 July 2004
A nucleotide transversion from guanine to uracil in the 23S rRNA confers linezolid resistance. We describe a real-time PCR using two Taqman probes that detects a single mutated allele among the genomes of
Enterococcus faecium and Enterococcus faecalis. Results were confirmed by a classical approach involving LabChip technology assayed with an Agilent Bioanalyzer 2100.
Linezolid is an oxazolidinone that has excellent activity against many gram-positive bacteria (17, 18, 20, 22). Preven-tion of initiaPreven-tion complex formaPreven-tion in protein biosynthesis is assumed to be the mechanism of action (27). In vitro resistance to linezolid is mediated via mutations in the central region of domain V of 23S rRNA (12, 21, 33) and/or by as-yet-unknown mechanisms (24, 31). However, resistance in wild-type isolates ofStaphylococcus,Enterococcus, andStreptococcusis conferred by a single nucleotide transversion from guanine to uracil at
position 2576 in 23S rRNA (Escherichia colinumbering) (9, 10,
21, 29, 33). Isolates for which the MICs areⱖ8 mg/liter are
defined as resistant (6). Identification of the resistance geno-type is complicated by the various numbers of 23S rRNA alleles among the genomes of these bacteria, for example, five
copies inStaphylococcus aureus, four inEnterococcus faecalis,
and six inEnterococcus faecium(2, 15, 19, 23). In vitro studies
showed that one out of six mutated alleles inE. faeciumand
two out of four mutated alleles inE. faecaliswere sufficient to
confer linezolid resistance (14, 15). A correlation between the
number of mutated alleles and the MIC was described forE.
faecalisand E. faecium(14, 15). Homozygous susceptible or homozygous resistant isolates (all alleles mutated) can be de-tected by molecular tests, such as DNA sequencing of PCR-amplified fragments (14, 15, 25) or a restriction digestion fol-lowing PCR amplification (15, 32). However, heterozygous linezolid-resistant isolates could be confirmed only by time-consuming and laborious methods (15). But these are exactly the isolates encountered in clinical practice: during linezolid therapy, primarily susceptible strains acquire resistance by stepwise mutation (1, 7, 8, 9) and probably by subsequent recombination (14). For rapid molecular detection and
molec-ular confirmation of such isolates, we chose a 5⬘ nuclease
real-time PCR assay with Taqman probes, a method which allows rapid, sensitive, and quantitative detection of single-nucleotide polymorphisms.
Seventy-two strains were included; 10 of them were linezolid
resistant (MICⱖ8g/ml). They all emerged during linezolid
therapy (one strain from the United States,E. faecium 3819
[8]; five from Austria,E. faecalis3932 and E. faecium3935,
3936, 3938, and 3939 [9]; and four from Germany isolated from
a single patient,E. faecalis3696 and 3697 andE. faecium3695
and 3698 [E. Halle, J. Padberg, S. Rousseau, I. Klare, G.
Werner, and W. Witte, Correspondence, Infection32:182–183,
2004]). The 10 enterococcal strains were partly clonally related or identical (e.g., strains 3935, 3936, 3938, and 3939; strains 3695 and 3698; and strains 3696 and 3698 [data not shown]), but the linezolid MICs for them were different, suggesting a different number of mutated 23S alleles. The 62
linezolid-susceptible isolates (MICs of 0.5 to 2 g/ml) were clonally
diverse (data not shown). DNA extraction and purification were done using standard procedures and commercial kits (QIAGEN, Hilden, Germany), and DNA was quantified by fluorescence labeling (Pico Green kit; Molecular Probes, Lei-den, The Netherlands). Classical PCR was performed with DNA beads (Amersham Pharmacia, Freiburg, Germany). Real-time PCR was done with an ABI 7000 using a SYBR Green kit and a Taqman kit (Applied Biosystems, Darmstadt, Germany). The assay was first evaluated with primers 23S_TQF and 23S_TQR and the SYBR Green kit, and a 100 pM concentration of each primer and 1 ng of purified PCR product (later with 2 ng of genomic DNA) amplified in a classical approach with primers 23S_F and 23S_R were then added (Table 1). The specificity of products was confirmed by melting-curve analysis. The assay design was then applied to the Taqman kit system, including two labeled Taqman probes
possessing 3⬘MGB (minor groove binder) VIC-LIZ-TQ-S
(de-tecting a wild-type or susceptible allele) and FAM-LIZ-TQ-R (detecting a mutated or resistant allele) probes (Table 1).
Optimization included 5⬘nuclease assays with various
concen-trations of primers (1, 10, and 100 pM), genomic DNA (0.066, 0.125, 0.25, 0.5, 1, and 2 ng), and probes (25, 50, 100, and 200 nM). An alternative FAM-LIZ-TQ-R probe was also tested (Table 1). All samples were assayed at least in triplicate.
The nucleotide transversion G2576T in linezolid-resistant enterococci generates a new restriction endonuclease site
rec-ognized by enzymes like MaeI (C2TAG) or NheI (G2CTAGC)
(mutated nucleotides are underlined) (15, 32). After endonu-clease treatment, linezolid-susceptible enterococci still showed a nondigested fragment, and homozygous linezolid-resistant
* Corresponding author. Mailing address: Robert Koch Institute, Wernigerode Branch, Burgstr. 37, D-38855 Wernigerode, Germany. Phone: 49 3943 679 210. Fax: 49 3943 679 207. E-mail: wernerg @rki.de.
5327
on May 15, 2020 by guest
http://jcm.asm.org/
enterococci showed two fragments (however, a 43-bp fragment was not detectable) (Fig. 1). The band pattern in linezolid-resistant enterococci with a heterozygous genotype revealed three fragments (43 bp not detectable). Restriction was done
with 5l of purified 23S_R-23S_F PCR product, 1⫻buffer,
and 20 U of NheI at 37°C for 0.5 to 2 h. Separation of frag-ments was done with a 2% agarose gel and with a LabChip 1000 kit. LabChip technology requires a Bioanalyzer 2100 (Agilent Technologies, Waldbronn, Germany), allowing quick (3-min/sample) and easy-to-perform separation of DNA, RNA, or proteins in specialized microglass capillary chips. Each lane possesses two internal standards which are scaled to the external standard running on each chip. Inherent BioSizing
software automatically calculates the size and quantity of each fragment in relation to the internal and external standards (Fig. 1 and Table 2).
Selection of strains.Data for the 10 linezolid-resistant
iso-lates (sevenE. faeciumand threeE. faecalisisolates) are given
in Table 2. The species was confirmed by using ddI-specific
primer pairs (4) (Table 1). The 23S rRNA alleles were ampli-fied classically by using genomic DNA and primers 23S_F and
23S_R. Digestion with NheI identified twoE. faeciumisolates
(3935 and 3939) and a singleE. faecalisisolate (3696) with all
23S alleles mutated (Fig. 1; see text below for details).
Taqman PCR.Homozygous linezolid-susceptible and
-resis-tant isolates, includingE. faeciumATCC 19434 (susceptible),
E. faecium3939 (resistant),E. faecalisV583 (susceptible), and
E. faecalis 3696 (resistant; see also above), were chosen to
establish a 5⬘nuclease assay. Primer concentrations of 100 pM
each and concentrations of 25 nM for the probe FAM-LIZ-TQ-R, detecting the resistant allele, and 100 nM for the probe VIC-LIZ-TQ-S, detecting the susceptible allele, led to the best results (data not shown in detail). An alternative probe detect-ing the resistant allele did not perform better (data not shown). As a threshold for all experiments, a value of 0.2 could be assigned: all nonspecific signals were beneath this value, and all specific signals were above it. Based on these results, gene dosage experiments were performed as follows. Genomic DNA from homozygous linezolid-susceptible and -resistant
test isolates (E. faecium and E. faecalis) was quantified and
mixed in a manner simulating DNA from wild-type isolates. All possible ratios of wild-type to mutated alleles were covered (forE. faecalis, 0:4, 1:3, 2:2, 3:1, and 4:0; forE. faecium, 0:6, 1:5, 2:4, 3:3, 4:2, 5:1, and 6:0). Existence of the appropriate alleles was precisely detected by the corresponding probes, which means that a single mutated allele was detected among
four inE. faecalisand among six inE. faecium (Fig. 2).
De-tection with the FAM-labeled probe revealed that the change
in the␦CTvalue was smaller the more alleles had mutated
(Fig. 2). Similarly, when using the VIC-labeled probe, the
change in the␦CTvalue was smaller the less alleles had
[image:2.585.49.542.81.237.2]mu-tated (data not shown). However, even under optimized test
FIG. 1. Detection of 23S alleles of linezolid-resistantE. faecium
and E. faecalis isolates by NheI digestion of purified pooled PCR fragments and subsequent separation in a LabChip 1000 measured in an Agilent Bioanalyzer 2100. The lowest and highest bands correspond to internal size markers. Lanes: M, external size markers; 1,E. faecium
ATCC 19434; 2,E. faecium3698; 3, E. faecium3695; 4,E. faecium
3819; 5,E. faecium3936; 6,E. faecium3938; 7,E. faecium3935; 8,E. faecium3939; 9,E. faecalisV583; 10,E. faecalis3932; 11,E. faecalis
[image:2.585.57.270.459.638.2]3697; 12,E. faecalis3696.
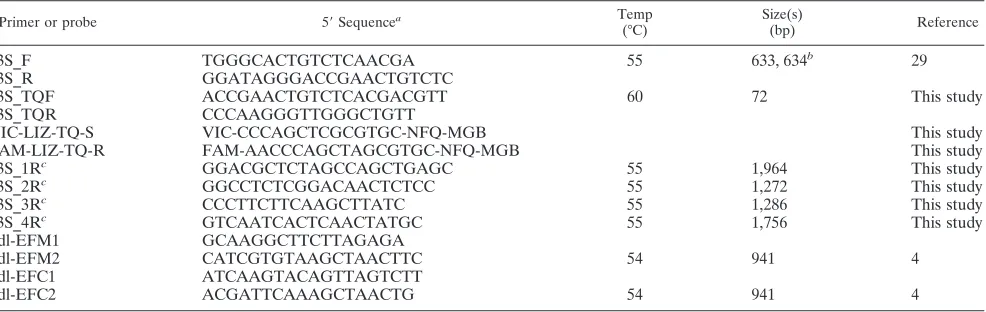
TABLE 1. Primers and probes used in this study
Primer or probe 5⬘Sequencea Temp (°C)
Size(s)
(bp) Reference
23S_F TGGGCACTGTCTCAACGA 55 633, 634b 29
23S_R GGATAGGGACCGAACTGTCTC
23S_TQF ACCGAACTGTCTCACGACGTT 60 72 This study
23S_TQR CCCAAGGGTTGGGCTGTT
VIC-LIZ-TQ-S VIC-CCCAGCTCGCGTGC-NFQ-MGB This study
FAM-LIZ-TQ-R FAM-AACCCAGCTAGCGTGC-NFQ-MGB This study
23S_1Rc GGACGCTCTAGCCAGCTGAGC 55 1,964 This study
23S_2Rc GGCCTCTCGGACAACTCTCC 55 1,272 This study
23S_3Rc CCCTTCTTCAAGCTTATC 55 1,286 This study
23S_4Rc GTCAATCACTCAACTATGC 55 1,756 This study
ddl-EFM1 GCAAGGCTTCTTAGAGA
ddl-EFM2 CATCGTGTAAGCTAACTTC 54 941 4
ddl-EFC1 ATCAAGTACAGTTAGTCTT
ddl-EFC2 ACGATTCAAAGCTAACTG 54 941 4
aNFQ, nonfluorescent quencher; VIC and FAM, 5⬘-linked fluorescence labels (Applied Biosystems); MGB, minor groove binder. bThe first size applies toE. faecalis; the second applies toE. faecium.
cThe forward primer is 23S_F.
on May 15, 2020 by guest
http://jcm.asm.org/
conditions, the exact number of mutated versus wild-type al-leles could not be quantitated. Nevertheless, the definite de-tection of a single mutated allele convinced us to prove the test scheme with wild-type isolates. All 72 isolates were investi-gated by using the approach described above (data not shown). All 10 linezolid-resistant isolates were precisely identified, al-lowing differentiation between isolates of homozygous and het-erozygous linezolid resistance genotypes based on a signal from one or both of the Taqman probes used (data not shown).
Classical approach.We analyzed PCR products amplifying
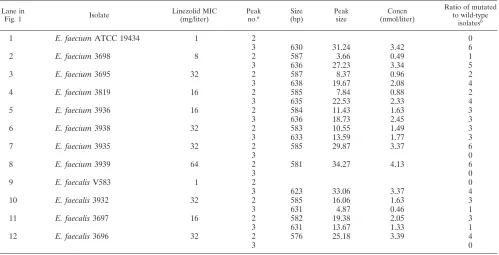
23S rRNA alleles (primers 23S_F and 23S_R) from our 10 linezolid-resistant enterococci digested with NheI (Fig. 1) (for details, see above). Quantitative evaluation based on LabChip technology and BioSizing software revealed a distribution of allele types in heterozygous enterococci based on calculated molarities of the corresponding bands (Fig. 1 and Table 2). The results have been confirmed by three independent
exper-iments. The genome of E. faecalis V583 has been released,
which allowed us to establish a verification test based on a separate amplification of all four 23S alleles followed by a subsequent confirmation of the corresponding allele type (G or T) by Taqman PCR (Table 1). The four PCRs were success-fully applied to our test isolates (3932 and 3697), a
homozy-gous susceptibleE. faecalisisolate (V583), and a homozygous
resistantE. faecalisisolate (3696). PCR products were purified,
quantified, and subjected to Taqman PCR. Each Taqman PCR showed a signal only with a single probe, suggesting a homozy-gous type for the four PCRs per isolate (data not shown). Calculations made with BioSizing software were confirmed as
follows: inE. faecalis3932, alleles 1, 3, and 4 showed a G-to-T
mutation, whereas inE. faecalis3697, alleles 1, 2, and 4 were
mutated (3696 possessed only T-type alleles; V583 possessed only G-type alleles).
A 5⬘ nuclease assay using Taqman probes is a modern,
time-saving PCR technique allowing online detection and quantification of amplified DNA (5, 11). Usage of distinctly designed probes, including an MGB motif, enables detection of single DNA nucleotide polymorphisms with great specificity and sensitivity. This is useful, for example, for detecting resis-tance characters based on single nucleotide polymorphisms such as fluoroquinolone or rifampin resistance (13, 28, 30). Even more sophisticated is the molecular detection of linezolid
resistance, in which inStaphylococcusandEnterococcus, four
to six gene copies code for 23S rRNA targeted by this antibi-otic. Woodford and coworkers (32) described a real-time PCR using LightCycler technology, which is different from Taqman technology (two probes per allele versus one probe per allele; sustaining of probes in LightCycler technology versus degra-dation of probes during Taqman PCR). Mutated and wild-type alleles were detected by a single probe and distinguished by different melting curves (32). This assay design cannot be ap-plied to other real-time PCR cyclers. We established a Taqman assay using two probes differing by a single nucleotide. Both probes are independently and in combination capable of de-tecting the susceptible and resistant allele types. The probes did not show any cross-hybridization: there were no nonspe-cific signals for the opposite allele types (Fig. 2). Our assay detected a single mutated 23S allele among four to six copies
in the genomes ofE. faecalisandE. faeciumin vitro and in the
[image:3.585.44.543.90.345.2]in vivo-generated resistant isolates. This detection allows pre-diction of future linezolid resistance during therapy even be-fore it is detected phenotypically, since a mutation in a single
TABLE 2. Results of NheI-digested 23S ribosomal DNA-pooled PCR products resolved with LabChip 1000 technology in an Agilent Bioanalyzer 2100
Lane in
Fig. 1 Isolate
Linezolid MIC (mg/liter)
Peak no.a
Size (bp)
Peak size
Concn (nmol/liter)
Ratio of mutated to wild-type
isolatesb
1 E. faeciumATCC 19434 1 2 0
3 630 31.24 3.42 6
2 E. faecium3698 8 2 587 3.66 0.49 1
3 636 27.23 3.34 5
3 E. faecium3695 32 2 587 8.37 0.96 2
3 638 19.67 2.08 4
4 E. faecium3819 16 2 585 7.84 0.88 2
3 635 22.53 2.33 4
5 E. faecium3936 16 2 584 11.43 1.63 3
3 636 18.73 2.45 3
6 E. faecium3938 32 2 583 10.55 1.49 3
3 633 13.59 1.77 3
7 E. faecium3935 32 2 585 29.87 3.37 6
3 0
8 E. faecium3939 64 2 581 34.27 4.13 6
3 0
9 E. faecalisV583 1 2 0
3 623 33.06 3.37 4
10 E. faecalis3932 32 2 585 16.06 1.63 3
3 631 4.87 0.46 1
11 E. faecalis3697 16 2 582 19.38 2.05 3
3 631 13.67 1.33 1
12 E. faecalis3696 32 2 576 25.18 3.39 4
3 0
a
Peaks 1 and 4 were identified for all lanes (internal markers) and are not included. b
Based on a comparison of peak areas. The number is the result of three independent experiments; here the data for a single experiment are given.
on May 15, 2020 by guest
http://jcm.asm.org/
23S allele might not in all cases be sufficient to confer linezolid
nonsusceptibility (14, 15). The findings for E. faecium 3698
demonstrated that even in clinical practice, a single mutated
allele is capable of mediating linezolid resistance (MIC, 8g/
ml). There is reasonable concern whether such isolates would be unambiguously identified by melting-curve analysis after real-time PCR using the LightCycler (32) or by sequencing pooled PCR products of all 23S alleles (15).
In conclusion, we established a Taqman PCR assay with two labeled probes detecting linezolid-resistant and -susceptible 23S alleles. Our assay design could easily be applied to other
genera, likeStaphylococcus, but slightly different
oligonucleo-tides would have to be used (e.g., forS. aureus, one nucleotide
mismatch for the two probes). With the help of LabChip
tech-nology, we were able to address the expected number of mu-tated alleles, which correlated well with the corresponding
linezolid MICs forE. faecalis, E. faecium, andS. aureus(14, 15;
T. A. Wichelhaus, S. Besier, V. Brade, and A. Ludwig, abstr. KMP021 from the 55th Annu. Mtg. of the DGHM, Int. J. Med.
Microbiol.293[Suppl. 36]:375–376, 2003) (Table 2). This result
illustrates again that modern techniques like multiplex PCR, real-time PCR, and DNA chip technology are appropriate tools to predict and/or confirm corresponding resistance phe-notypes (3, 16, 26).
We acknowledge skillful technical assistance by Bianca Hildebrandt and Carola Konstabel. We thank R. Patel, F. J. Allerberger, and E. Halle for providing the linezolid-resistant strains for our analyses. FIG. 2. Detection of the G or T allele type of 23S ribosomal DNA by real-time PCR using two labeled probes. In vitro gene dosage experiments vary the number of wild-type versus mutated alleles inE. faecium(A) andE. faecalis(B) (see text for details). Only results for the FAM-LIZ-TQ-R probe are shown. For better visibility, only one representative per allele mix is shown. Labels indicate the numbers of mutated alleles relative to the overall number of 23S ribosomal DNA copies per genome (E. faecium, 6;E. faecalis, 4). Delta Rn, difference of fluorescence signals of a given template and the no-template control.
on May 15, 2020 by guest
http://jcm.asm.org/
REFERENCES
1.Auckland, C., L. Teare, F. Cooke, M. E. Kaufmann, M. Warner, G. Jones, K. Bamford, H. Ayles, and A. P. Johnson.2002. Linezolid-resistant enterococci: report of the first isolates in the United Kingdom. J. Antimicrob. Chemother.
50:743–746.
2.Baba, T., F. Takeuchi, M. Kuroda, H. Yuzawa, K. Aoki, A. Oguchi, Y. Nagai, N. Iwama, K. Asano, T. Naimi, H. Kuroda, L. Cui, K. Yamamoto, and K. Hiramatsu.2002. Genome and virulence determinants of high virulence community-acquired MRSA. Lancet359:1819–1827.
3.Call, D. R., M. K. Bakko, M. J. Krug, and M. C. Roberts.2003. Identifying antimicrobial resistance genes with DNA microarrays. Antimicrob. Agents Chemother.47:3290–3295.
4.Dutka-Malen, S., S. Evers, and P. Courvalin.1995. Detection of glycopep-tide resistance genotypes and identification to the species level of clinically relevant enterococci by PCR. J. Clin. Microbiol.33:24–27. (Erratum,33:
1434.)
5.Ellerbrok, H., H. Nattermann, M. Oezel, L. Beutin, B. Appel, G. Pauli.2002. Rapid and sensitive identification of pathogenic and apathogenicBacillus anthracisby real-time PCR. FEMS Microbiol. Lett.214:51–59.
6.European Committee on Antimicrobial Susceptibility Testing (EUCAST) of the European Society of Clinical Microbiology and Infectious Diseases (ESCMID).2001. Linezolid breakpoints. Clin. Microbiol. Infect.7:283–284. 7.Gonzales, R. D., P. C. Schreckenberger, M. B. Graham, S. Kelkar, K. DenBesten, and J. P. Quinn.2001. Infections due to vancomycin-resistant
Enterococcus faeciumresistant to linezolid. Lancet357:1179.
8.Herrero, I. A., N. C. Issa, and R. Patel.2002. Nosocomial spread of linezolid-resistant, vancomycin-resistantEnterococcus faecium. N. Engl. J. Med.346:
867–869.
9.Johnson, A. P., L. Tysall, M. W. Stockdale, N. Woodford, M. E. Kaufmann, M. Warner, D. M. Livermore, F. Asboth, and F. J. Allerberger.2002. Emerg-ing linezolid-resistantEnterococcus faecalisandEnterococcus faecium iso-lated from two Austrian patients in the same intensive care unit. Eur. J. Clin. Microbiol. Infect. Dis.21:751–754.
10.Jones, R. N., P. Della-Latta, L. V. Lee, Douglas, and J. Biedenbach.2002. Linezolid-resistantEnterococcus faecium isolated from a patient without prior exposure to an oxazolidinone: report from the SENTRY antimicrobial surveillance program. Diagn. Microbiol. Infect. Dis. 42:137–139. 11.Klein, D.2002. Quantification using real-time PCR technology: applications
and limitations. Trends Mol. Med.8:257–260.
12.Kloss, P., L. Xiong, and D. L. Shinabarger.1999. Resistance mutations in 23S rRNA identify the site of action of the protein synthesis inhibitor lin-ezolid in the ribosomal peptidyl transferase center. J. Mol. Biol.294:93–101. 13.Lapierre, P., A. Huletsky, V. Fortin, F. J. Picard, P. H. Roy, M. Ouellette, and M. G. Bergeron.2003. Real-time PCR assay for detection of fluoro-quinolone resistance associated withgrlAmutations inStaphylococcus au-reus. J. Clin. Microbiol.41:3246–3251.
14.Lobritz, M., R. Hutton-Thomas, S. Marshall, and L. B. Rice.2003. Recom-bination proficiency influences frequency and locus of mutational resistance to linezolid inEnterococcus faecalis. Antimicrob. Agents Chemother.47:
3318–3320.
15.Marshall, S. H., C. J. Donskey, R. Hutton-Thomas, R. A. Salata, and L. B. Rice.2002. Gene dosage and linezolid resistance inEnterococcus faecium
andEnterococcus faecalis. Antimicrob. Agents Chemother.46:3334–3336. 16.Monecke, S., I. Leube, and R. Ehricht.2003. Simple and robust array-based
methods for the parallel detection of resistance genes ofStaphylococcus aureus. Genome Lett.2:106–118.
17.Mutnick, A. H., V. Enne, and R. N. Jones.2003. Linezolid resistance since 2001: SENTRY Antimicrobial Surveillance Program. Ann. Pharmacother.
37:769–774.
18.Noskin, G. A., F. Siddiqui, V. Stosor, D. Hacek, and L. R. Peterson.1999. In vitro activities of linezolid against important gram-positive bacterial patho-gens including vancomycin-resistant enterococci. Antimicrob. Agents Che-mother.43:2059–2062.
19.Paulsen, I. T., L. Banerjei, G. S. A. Myers, K. E. Nelson, R. Seshadri, T. D. Read, D. E. Fouts, J. A. Eisen, S. R. Gill, J. F. Heidelberg, H. Tettelin, R. J. Dodson, L. Umayam, L. Brinkac, M. Beanan, S. Daugherty, R. T. DeBoy, S. Durkin, J. Kolonay, R. Madupu, W. Nelson, J. Vamathevan, B. Tran, J. Upton, T. Hansen, J. Shetty, H. Khouri, T. Utterback, D. Radune, K. A. Ketchum, B. A. Dougherty, and C. M. Fraser.2003. Role of mobile DNA in the evolution of vancomycin-resistantEnterococcus faecalis. Science299:
2071–2074.
20.Perry, C. M., and B. Jarvis.2001. Linezolid—a review of its use in the management of serious gram-positive infections. Drugs61:525–551. 21.Prystowsky, J., F. Siddiqui, J. Chosay, D. L. Shinabarger, J. Millichap, L. R.
Peterson, and G. A. Noskin.2001. Resistance to linezolid: characterization of mutations in rRNA and comparison of their occurrences in vancomycin-resistant enterococci. Antimicrob. Agents Chemother.45:2154–2156. 22.Rubinstein, E., S. K. Cammarata, T. H. Oliphant, R. G. Wunderink, and the
Linezolid Nosocomial Pneumonia Study Group. 2001. Linezolid (PNU-100766) versus vancomycin in the treatment of hospitalized patients with nosocomial pneumonia: a randomized, double-blind, multicenter study. Clin. Infect. Dis.32:402–412.
23.Ruggero, K. A., L. K. Schroeder, P. C. Schreckenberger, A. S. Mankin, and J. P. Quinn. 2003. Nosocomial superinfections due to linezolid-resistant
Enterococcus faecalis: evidence for a gene dosage effect on linezolid MICs. Diagn. Microbiol. Infect. Dis.47:511–513.
24.Sander, P., L. Belova, Y. G. Kidan, P. Pfister, A. S. Mankin, and E. C. Bo¨ttger.2002. Ribosomal and non-ribosomal resistance to oxazolidinones: species-specific idiosyncrasy of ribosomal alterations. Mol. Microbiol.46:
1295–1304.
25.Sinclair, A., C. Arnold, and N. Woodford.2003. Rapid detection and esti-mation by pyrosequencing of 23S rRNA genes with a single nucleotide polymorphism conferring linezolid resistance in enterococci. Antimicrob. Agents Chemother.47:3620–3622.
26.Strommenger, B., C. Kettlitz, G. Werner, and W. Witte.2003. Multiplex PCR assay for simultaneous detection of nine clinically relevant antibiotic resis-tance genes inStaphylococcus aureus. J. Clin. Microbiol.41:4089–4094. 27.Swaney, S. M., H. Aoki, M. C. Ganoza, and D. L. Shinabarger.1998. The
oxazolidinone linezolid inhibits initiation of protein synthesis in bacteria. Antimicrob. Agents Chemother.42:3251–3255.
28.Torres, M. J., A. Criado, M. Ruiz, A. C. Llanos, J. C. Palomares, and J. Aznar.2003. Improved real-time PCR for rapid detection of rifampin and isoniazid resistance inMycobacterium tuberculosisclinical isolates. Diagn. Microbiol. Infect. Dis.45:207–212.
29.Tsiodras, S., H. S. Gold, G. Sakoulas, G. M. Eliopoulos, C. Wennersten, L. Venkataraman, R. C. Moellering, and M. J. Ferraro.2001. Linezolid resis-tance in a clinical isolate ofStaphylococcus aureus. Lancet358:207–208. 30.Walker, R. A., N. Saunders, A. J. Lawson, E. A. Lindsay, M. Dassama, L. R.
Ward, M. J. Woodward, R. H. Davies, E. Liebana, and E. J. Threlfall.2001. Use of a LightCyclergyrAmutation assay for rapid identification of muta-tions conferring decreased susceptibility to ciprofloxacin in multiresistant
Salmonella entericaserotype Typhimurium DT104 isolates. J. Clin. Micro-biol.39:1443–1448.
31.Willems, R. J., J. Top, D. J. Smith, D. I. Roper, S. E. North, and N. Woodford.2003. Mutations in the DNA mismatch repair proteins MutS and MutL of oxazolidinone-resistant or -susceptibleEnterococcus faecium. An-timicrob. Agents Chemother.47:3061–3066.
32.Woodford, N., L. Tysall, C. Auckland, M. W. Stockdale, A. J. Lawson, R. A. Walker, and D. M. Livermore.2002. Detection of oxazolidinone-resistant
Enterococcus faecalisandEnterococcus faeciumstrains by real-time PCR and PCR-restriction fragment length polymorphism analysis. J. Clin. Microbiol.
40:4298–4300.
33.Xiong, L., P. Kloss, S. Douthwaite, N. M. Andersen, S. Swaney, D. L. Shina-barger, and A. S. Mankin.2000. Oxazolidinone resistance mutations in 23S rRNA ofEscherichia colireveal the central region of domain V as the primary site of drug action. J. Bacteriol.182:5325–5331.