0095-1137/09/$08.00⫹0 doi:10.1128/JCM.00551-09
Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Aspergillus
Section
Fumigati
Typing by PCR-Restriction
Fragment Polymorphism
䌤
Janet F. Staab,
1S. Arunmozhi Balajee,
2and Kieren A. Marr
1*
Johns Hopkins University School of Medicine, Baltimore, Maryland,1and Mycotic Diseases Branch, Centers for Disease Control and
Prevention, Atlanta, Georgia2
Received 18 March 2009/Returned for modification 14 April 2009/Accepted 21 April 2009
Recent studies have shown that there are multiple clinically important members of theAspergillussection
Fumigatithat are difficult to distinguish on the basis of morphological features (e.g.,Aspergillus fumigatus,A. lentulus, andNeosartorya udagawae). Identification of these organisms may be clinically important, as some species vary in their susceptibilities to antifungal agents. In a prior study, we utilized multilocus sequence
typing to describeA. lentulus as a species distinct fromA. fumigatus. The sequence data show that the gene
encoding-tubulin,benA, has high interspecies variability at intronic regions but is conserved among isolates
of the same species. These data were used to develop a PCR-restriction fragment length polymorphism
(PCR-RFLP) method that rapidly and accurately distinguishesA. fumigatus,A. lentulus, andN. udagawae, three
major species within the sectionFumigatithat have previously been implicated in disease. Digestion of thebenA
amplicon with BccI generated unique banding patterns; the results were validated by screening a collection of
clinical strains and by in silico analysis of thebenAsequences ofAspergillusspp. deposited in the GenBank
database. PCR-RFLP of benA is a simple method for the identification of clinically important, similar
morphotypes ofAspergillusspp. within the sectionFumigati.
Aspergillus fumigatuscontinues to be the most common
eti-ological agent of invasive aspergillosis. Identification ofA.
fu-migatushas historically been based on morphological features;
the organism is typified by green conidia produced in chains basipetally from uniseriate phialides (15). However, recent studies have shown that species identification on the basis of morphology alone is problematic; some isolates are pigment-less or sporulate poorly, and growth conditions can influence the morphology, making identification difficult (9). Moreover,
some A. fumigatus-related species have similar phenotypic
characteristics, making misidentification common when mor-phological examination alone is used (4, 5, 7).
Molecular phylogenetic typing methods based on the DNA sequences of gene sets have begun to complement and some-times supersede traditional phenotypic species identification methods (for a review, see reference 6). For example, the conserved ribosomal internal transcribed spacer and the D1 and D2 (domains of the large ribosomal subunit) regions have been used to identify molds to the genus and species levels. At times, these regions are inadequate for the intrasection iden-tification of species, and comparative sequence analyses of other loci are required to make an accurate identification. Recently, sequence data have been used to uncover new and previously described species among clinical isolates morpho-logically identified as “A. fumigatus” in clinical microbiology laboratories (3–5, 7, 10, 11). These studies frequently used partial sequence data from two or three genes to generate phylogenies that infer genetic relatedness and species bound-aries. We previously identifiedA. lentulusas a unique species
on the basis of its phylogenetic relationship toA. fumigatusand other Aspergillus spp. using multilocus sequence typing (MLST) (4). As MLST is not readily performed in clinical microbiology laboratories, we sought to develop a rapid mo-lecular typing method based on PCR of evolutionary informa-tive DNA regions. In the present study, we devised a species identification scheme based on thebenA sequence to rapidly identify similar morphotypes in the section Fumigati (A.
fu-migatus, A. lentulus, andNeosartorya udagawae) using
PCR-restriction length fragment polymorphisms (RFLPs). We vali-dated this method by screening a collection of clinical and environmental isolates and by using DNA analysis computer software (i.e., in silico analysis) to examine additional A.
lentulusand N. udagawae benAsequences deposited in the
GenBank database.
MATERIALS AND METHODS
Isolates.Three hundred eighteen totalAspergillusisolates were used in this study. They included clinical and environmental isolates in our culture collection originating from the University of Washington and the Fred Hutchinson Cancer Research Center. We testedA. fumigatus,A. lentulus,Neosartorya udagawae,A. fumigatusvar.ellipticus,N. pseudofischeri, andN. fischeri. Sixty-four strains were obtained from the Centers for Disease Control and Prevention (Atlanta, GA), and 12 were obtained from the University of Texas in San Antonio (Fungus Testing Laboratory, San Antonio, TX).N. fischeri(NRRL 4075) andA. fumigatus
var.ellipticus(NRRL 5109) were obtained from S. W. Peterson (U.S. Depart-ment of Agriculture, Peoria, IL). ReferenceA. fumigatusstrains Af293 and B-5233 were obtained from David Denning (University of Manchester, Manches-ter, United Kingdom) and June Kwon-Chung (National Institutes of Health, Bethesda, MD), respectively. The collection included three poorly sporulating strains that were previously identified asA. fumigatus(strains FH1, FH6, and FH219 (5) and that upon retesting with an expanded MLST scheme were found to beA. lentulus(data not shown) (7). The isolates were routinely maintained on potato dextrose agar (Difco, Becton Dickinson and Co.) at 37°C and frozen at ⫺80°C in 8% skim milk for longer-term storage. All aspergilli were identified by morphology and/or previous MLST (3, 5) studies.
Genomic DNA preparation.Chromosomal DNA was prepared from mycelia grown in Sabouraud dextrose broth (Sigma, St. Louis, MO) at 37°C for 48 h with * Corresponding author. Mailing address: Johns Hopkins University
School of Medicine, 720 Rutland Ave., Ross-1064, Baltimore, MD 21205. Phone: (443) 614-9141. Fax: (443) 614-8518. E-mail: kmarr4 @jhmi.edu.
䌤Published ahead of print on 29 April 2009.
2079
on May 16, 2020 by guest
http://jcm.asm.org/
shaking. Mycelia were harvested between sheets of Miracloth (Calbiochem, La Jolla, CA), and approximately 200 mg (wet weight) was used for DNA extraction by use of an Epicentre kit (MasterPure yeast DNA purification kit; Epicentre Biotech), as modified by Jin et al. (13). DNA quality and yield were determined by agarose gel electrophoresis by standard methods (17).
PCR-RFLP of a region of-tubulin.Previously reported data (4) together with the results of an in-house MLST screen (S. A. Balajee and K. A. Marr, unpub-lished results) were used to develop a PCR-RFLP based on a 492-bp region of
A. fumigatus benA(GenBank accession number AFUA_2G14990) that spans three introns near the 5⬘region of the orthologous genes inEmericella nidulans
(anamorph,A. nidulans) andA. flavus(8). Amplification ofbenAofAspergillus
spp. was performed with approximately 75 ng of genomic DNA, forward primer tub1 (5⬘-AATTGGTGCCGCTTTCTGG-3⬘), and reverse primertub2 (5⬘-A GTTGTCGGGACGGAATAG-3⬘) (4) as follows: the reactions were performed in a volume of 50l consisting of PCR buffer (25 mM TAPS [N -Tris(hydroxy-methyl)methyl-3-aminopropanesulfonic acid; pH 9.3], 50 mM KCl, 2 mM MgCl2,
1 mM 2-mercaptoethanol); 0.2 mM each of dATP, dGTP, dCTP, and dTTP; 0.2 pmol (each) primer; and 2.5 of TaKaRa ExTaqpolymerase (Takara Bio Inc., Japan). After an initial denaturation step of 94°C for 2 min, the reactions were cycled 30 times at 94°C for 30 s, 55°C for 30 s, and 72°C for 45 s in a Techne TC-512 thermal cycler (Techne Inc.). The reactions were terminated with a final incubation at 72°C for 5 min following the last cycle. The amplification ofbenAwas verified by agarose (1.5%) gel electrophoresis of a portion (5l) of each reaction mixture. Digestion of the amplicons was performed with 10l of the reaction mixtures in a 50-l reaction volume containing 1⫻NE Buffer 1 (10 mM Bis-Tris-propane-HCl, 10 mM MgCl2, 1 mM dithiothreitol [pH 7.0]), 100g/ml bovine
serum albumin, and 1.0 U of BccI enzyme (New England Biolabs) in a water bath at 37°C for 1 h. The resulting DNA fragments (25l) were separated in a 3.5% agarose gel and visualized by staining with ethidium bromide, according to standard methods (17). The gel electrophoresis images were captured with a GelDoc-ItTS imaging system (UVP, LLC, Upland, CA), and manipulated with ImageJ processing and analysis software (http://rsb.info.nih.gov/ij/).
Separate experiments investigated the star activity (nonspecific digestion out-side a restriction enzyme’s recognition site) of BccI that became apparent during the large-scale screening ofAspergillusisolates. Digestion of thebenAamplicons with⬎5 U of BccI in a 50-l volume resulted in restriction fragments whose sizes did not correlate with the predicted sizes (data not shown). Incubation of theA. fumigatus benAamplicon with XhoI (New England Biolabs) served as control for reactions with excess units of restriction enzyme. The latter digestions were performed with 10l of the strain Af293benAamplicon in a 50-l reaction
mixture containing 1⫻NE Buffer 2 (50 mM NaCl, 10 mM Tris-HCl, 10 MgCl2
1 mM dithiothreitol [pH 7.9]) and 10 U of XhoI at 37°C for 1 h. Digestion of the Af293benAamplicon with XhoI cleaved the DNA once to produce two frag-ments of 426 bp and 66 bp, respectively.
DNA data manipulations. ThebenA sequences fromA. fumigatus Af293 (GenBank accession number AFUA_2G14990),A. lentulusFH5 (GenBank accession number AY738513 [4]), andN. udagawaeCDC58/FH83 (GenBank accession number DQ058391 [5]) were aligned by using the ClustalW algorithm within MacVector (version 8.0) software (MacVector, Inc., Cary, NC). Restriction maps of thebenAsequences were also generated with MacVector software. Other
benAsequences deposited in the GenBank database were used for the in silico analysis of BccI sites (MacVector software) within the amplified region (GenBank accession numbers are given in parentheses): thebenAsequences ofA. lentulus
isolates (total of 21) NRRL 35553 (EF669826), NRRL 35552 (EF669825), NRRL 35551 (EF669824), CBS 612.97 (DQ534081), IFM 41090 (AB248073), IFM 47063 (AB248074), CM-4428 (EU310870), CM-4426 (EU310869), CM-4420 (EU310868), CM-4387 (EU310868), CM-4387 (EU310866), CM-4370 (EU310865), CM-3583 (EU310853), CM-3537 (EU310851), CM-3364 (EU310850), CM-1290 (EU310839), CM-3134 (EU310842), CM-4330 (EU310864), CM-3599 (EU310854), CM-3538 (EU310852), and CM-4415 (EU310867) andN. udagawaeisolates (total of 6) IBT 23363 (DQ534161), KACC 41683 (DQ534102), KACC F3759 (DQ534102), CBS 154.89 (DQ534080), CBM FA-0703 (AB248303), and FA-0702 (AB248302).
RESULTS
Previously (5), we developed a PCR-RFLP screen based on rodlet A to discriminate betweenA. fumigatusand potentialA.
lentulus isolates. However, that RFLP is based on a single
polymorphism within therodAamplicon, and it does not dis-tinguish otherAspergillusspp. fromA. fumigatus. MLST data evaluating seven loci (calmodulin, carboxypeptidase 5,
glycer-aldehyde 3-phosphate dehydrogenase, class III chitinase G, rodlet A,-tubulin, and catalase) revealed thatbenA(the gene encoding -tubulin) produced the most useful phylogenetic data (4, 5; unpublished data). We elected to utilize BccI to digest thebenAamplicon in a PCR-RFLP scheme based on the availability of the enzyme, the sizes of the restriction fragments, and the number of polymorphic sites encoded by more than 1 nucleotide difference among species; thebenAhomology align-ment betweenA. fumigatus,A. lentulus, andN. udagawaeand the BccI restriction digest map are shown in Fig. 1A and B, respec-tively. ThisbenAPCR-RFLP is predicted to be useful for distin-guishing most known section Fumigati members, including A.
fumigatus,A. lentulus,N. udagawae, andN. pseudofischeri(Table
1). However, the digestion ofbenAamplicons with BccI fromN.
fischeriorA. fumigatusvar.ellipticusis not expected to
differen-tiate these organisms fromA. fumigatus(Table 1).
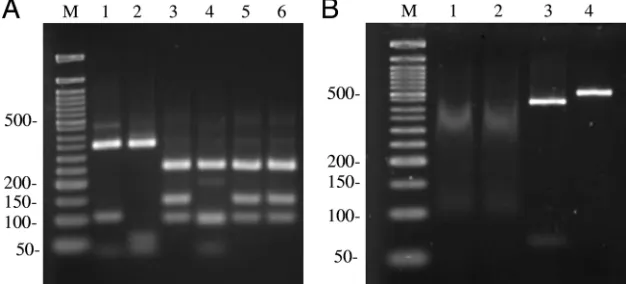
Digestion of the benA amplicons produced the expected DNA fragments for each Aspergillus species (Fig. 2A). The
benAdigestion pattern for each organism was stable, as judged with independent DNA samples prepared from biological rep-licates (at least two independent cultures; data not shown). ThebenAPCR-RFLP was further tested with 284A. fumigatus
isolates, 15 A. lentulus isolates, and 10N. udagawae/A.
uda-gawaeisolates (A. udagawaeis the anamorph ofN. udagawae;
seven of the nine isolates were previously classified asA.
uda-gawae[5]) and was found to generate results 100% concordant
with the predicted DNA banding patterns for each species. We also tested three additionalN. pseudofischeriisolates (isolates FH240, FH242, and FH274 [3]) and confirmed the digestion fragment pattern predicted by thebenAsequence data.
During the large-scale screening of our isolates, we found the need to optimize the BccI digestions. The enzyme exhibits a high level of star activity and tends to lose activity over time (P. Zhang, production manager, New England Biolabs, per-sonal communication). Fresh enzyme used at the indicated concentration (see Materials and Methods; 1 U/50 l diges-tion) for 1 h at 37°C consistently produced complete digestion of thebenA amplicons. The number of units per digest was increased to 5 U as the enzyme aged for several months during storage at⫺20°C, as determined by the detection of an intact amplicon, in addition to digestion products. The use of⬎5 U of BccI per digest increased the star activity and generated DNA fragments reminiscent of degradation (Fig. 2B).
All of our A. lentulus and N. udagawae clinical isolates originated from sites within the United States. To determine if the benABccI polymorphic sites are conserved in other geographically distinct isolates and to increase the robust-ness of our analysis, we examined A. lentulus (21 entries)
and N. udagawae (6 entries) benA sequences deposited in
the GenBank database. This in silico analysis ofA. lentulus
andN. udagawaesequences revealed that the BccI
polymor-phisms were conserved within each species and that the predicted DNA fragment sizes were in 100% agreement with the experimental results generated with our isolates (data not shown). This suggests that the BccI polymor-phisms are genetically stable and characteristic of each spe-cies and are thus a useful marker for spespe-cies discrimination
ofA. fumigatusmorphotypes.
on May 16, 2020 by guest
http://jcm.asm.org/
DISCUSSION
In this study, we have developed a PCR-RFLP method that can be employed to differentiateAspergillusspecies within the sectionFumigati, with the BccI polymorphisms at a 5⬘region of
benAdistinguishing between 285A. fumigatusstrains (clinical
and environmental), 37A. lentulusstrains, and 17N. udagawae
strains and sequences examined. Although the majority of cases of invasive aspergillosis appear to be attributed to A.
fumigatus, other related and often misidentified Aspergillus
[image:3.585.59.523.65.605.2]spp. have been uncovered as causes of invasive infections (4, 5,
FIG. 1. PCR-RFLP ofbenAamplicons. (A) Homology alignment of thebenAamplicons from each species with the ClustalW algorithm. Potential BccI sites (5⬘-CCATCNNNNN-3⬘) are underlined. Restriction sites centered atA. fumigatusnucleotides 106 and 253 have two site polymorphisms. Species are designated at the left (Af,A. fumigatus; Al,A. lentulus; Nu,N. udagawae). (B) Restriction digest maps of each species. BccI sites are shown at the top of each amplicon.
on May 16, 2020 by guest
http://jcm.asm.org/
12); the true burden of these infections remains to be fully elucidated. However, accurate identification may have clinical relevance, as different species are not equally susceptible to antifungal drugs in vitro. For example,A. lentulusexhibits low and variable in vitro susceptibilities to itraconazole, voricon-azole, amphotericin B, and the echinocandins (4, 7, 18). In addition, others have reported decreased in vitro susceptibili-ties ofA. lentulusisolates to ravuconazole, posaconazole, and terbinafine (1).
The recognition thatAspergillusspecies identification solely on the basis of morphological features is insufficient has pushed the development of molecular methods for the purpose of species identification. The current strategy of using se-quenced-based identification methods relies upon the accuracy of banked sequence data together with the quality of the data generated from the unknown isolate. Once the sequence data are aligned, a homology score is used to make the identifica-tion; and this score may be influenced by the quality of the sequences, the length of the sequence data, and the software used to make the alignment. However, the use of a restriction site polymorphism within a conserved gene for species identi-fication does not depend upon sequence quality and, most importantly, does not rely upon arbitrary cutoff homology scores. Furthermore, same-species homology score cutoff
val-ues have not been standardized to date, and there is a lack of consensus on how best to use sequence homology data to identify a species (2). Thus, sequence homology-based meth-ods for the identification of fungi may be approaching the “gold standard,” but a critical parameter for the determination of relatedness, the percent standard homology value, has not been standardized. Other molecular methods that do not rely on pure sequence data can complement or altogether circum-vent the need for the sequencing of genetic loci.
We previously developed a molecular identification method that uncoveredA. lentuluson the basis of sequence data for five genetic loci that included-tubulin (benA) (4).-Tubulin has proven to be useful for phylogenetic relatedness studies of
Aspergillusand related species (3–5, 8, 11, 16), because it
ap-pears to be a slowly evolving, conserved gene with a high degree of interspecies variability. Recently, an editorial ad-dressing the molecular identification ofAspergillusspp. recom-mended the use of comparative sequence analysis of-tubulin for species identification, once isolates are assigned to a spe-cies complex or section (e.g., the sectionFumigati) on the basis of the nuclear ribosomal internal transcribed region and/or traditional morphological identification methods (2). We have taken this identification method a step further and simplified it by avoiding sequence-based homology caveats. The PCR-RFLP described here takes advantage of BccI restriction site polymorphisms withinbenAthat are unique toA. fumigatus,A.
lentulus, andN. udagawae. This methodology can be used
to-gether with other identification methods to confirm the iden-tities of A. fumigatusisolates and discriminate those isolates
from A. lentulus and N. udagawae. However, one limitation
that we encountered was the inability to distinguishA.
fumiga-tusfromN. fischeriandA. fumigatusvar.ellipticus. This is not
surprising, as N. fischeri and A. fumigatus var. ellipticus are closely related toA. fumigatusphylogenetically (8) and perhaps do not warrant separate species designations.N. fischeri has been reported to cause less than five human infections, andA.
fumigatusvar.ellipticushas not been associated with disease in
[image:4.585.43.282.90.182.2]humans; thus, the clinical importance of these isolates remains undefined. Another related but apparently unusual causative
TABLE 1. Restriction size fragments generated by digestingbenA amplicons from different aspergilli
Organism GenBank
accession no.
Amplicon
size (bp)a Predicted DNA
fragment sizes (bp)
A. fumigatus AFUA_2G14990 492 249, 144, 99 A. lentulus AY738513 492 348, 105, 39 N. udagawae DQ058391 492 347, 60, 46,b39b
N. pseudofischeri AF057325 483 250, 99,b94,b39
N. fischeri AF057322 488 247, 142, 98 A. fumigatusvar.
ellipticus
AF057314 493 249, 145, 99
aSequences were edited at their 5⬘and 3⬘ends to reflect the predicted size of
the amplicon generated with the oligonucleotides used in the PCR.
bFragments not resolved by agarose gel electrophoresis.
FIG. 2. (A) Gel electrophoresis of the fragments generated from the digestion of thebenAamplicons with BccI. Lane M, 50-bp DNA ladder (New England Biolabs); lane 1,A. lentulus; lane 2,N. udagawae; lane 3,A. fumigatus; lane 4,N. pseudofischeri; lane 5,N. fischeri; lane 6,A. fumigatus var.ellipticus. The molecular sizes of the DNA standards (in bp) are indicated at the left. (B) Nonspecific (star activity) digestion of BccI when it is used in excess. Gel electrophoresis of theA. fumigatus benAamplicon digested with 10 U of BccI (lanes 1 and 2) or 10 U of XhoI (lane 3) for 1 h at 37°C. Digestion of thebenAamplicon with excess XhoI generated the expected 426- and 66-bp DNA fragments. Lane 4, intactA. fumigatus benAamplicon (492 bp); lane M, 50-bp DNA ladder. The molecular sizes of the DNA standards (in bp) are indicated at the left.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:4.585.135.448.526.668.2]agent of aspergillosis, N. pseudofischeri, which displays de-creased in vitro susceptibilities to voriconazole and amphoter-icin B (3), was readily identified by using BccIbenA polymor-phisms (Table 1 and Fig. 1A).
Other previously unknown agents of aspergillosis have emerged from the analysis of atypical A. fumigatus clinical isolates that were reclassified by molecular identification meth-ods (1, 14, 18). TheseA. fumigatus-related organisms include
A. arvii, A. fumisynnematus, A. viridinutans, A. fumigatiaffinis,
andN. hiratsukae, which may be of some clinical importance,
as several of these species display reduced susceptibilities to multiple antifungal drugs (1, 18). Because these organisms appear to be rare etiologic agents of aspergillosis, the sequence data from these isolates are limited and not available in suffi-cient numbers for the robust evaluation of our benA PCR-RFLP method at this time.
The true prevalence of these closely related, non-A. fumiga-tusorganisms as causes of human disease is unknown, and we do not yet understand the potential importance of their vari-able in vitro susceptibility profiles. However, an increased number of studies have reported these isolates, particularlyA.
lentulus, among clinical culture collections (1, 18). The method
described herein will be useful both for microbiology labora-tories and for investigations evaluating the prevalence and significance of these newly described opportunistic pathogens.
ACKNOWLEDGMENTS
This study was supported by a National Institutes of Health grant (R21 AI067971) to K.A.M.
We thank Leon W. Razai for technical assistance.
REFERENCES
1.Alcazar-Fuoli, L., E. Mellado, A. Alastruey-Izquierdo, M. Cuenca-Estrella, and J. L. Rodriguez-Tudela.2008.AspergillussectionFumigati: antifungal susceptibility patterns and sequence-based identification. Antimicrob. Agents Chemother.52:1244–1251.
2.Balajee, S. A., A. M. Borman, M. E. Brandt, J. Cano, M. Cuenca-Estrella, E. Dannaoui, J. Guarro, G. Haase, C. C. Kibbler, W. Meyer, K. O’Donnell,
C. A. Petti, J. L. Rodriguez-Tudela, D. Sutton, A. Velegraki, and B. L. Wickes.2009. Sequence-based identification ofAspergillus,Fusarium, and
Mucoralesspecies in the clinical mycology laboratory: where are we and where should we go from here? J. Clin. Microbiol.47:877–884.
3.Balajee, S. A., J. Gribskov, M. Brandt, J. Ito, A. Fothergill, and K. A. Marr. 2005. Mistaken identity:Neosartorya pseudofischeriand its anamorph mas-querading asAspergillus fumigatus. J. Clin. Microbiol.43:5996–5999. 4.Balajee, S. A., J. L. Gribskov, E. Hanley, D. Nickle, and K. A. Marr.2005.
Aspergillus lentulussp. nov., a new sibling species ofA. fumigatus.Eukaryot. Cell4:625–632.
5.Balajee, S. A., D. Nickle, J. Varga, and K. A. Marr.2006. Molecular studies reveal frequent misidentification ofAspergillus fumigatusby morphotyping. Eukaryot. Cell5:1705–1712.
6.Balajee, S. A., L. Sigler, and M. E. Brandt.2007. DNA and the classical way: identification of medically important molds in the 21st century. Med. Mycol. 45:475–490.
7.Balajee, S. A., M. Weaver, A. Imhof, J. Gribskov, and K. A. Marr.2004.
Aspergillus fumigatusvariant with decreased susceptibility to multiple anti-fungals. Antimicrob. Agents Chemother.48:1197–1203.
8.Geiser, D. M., J. C. Frisvad, and J. W. Taylor.1998. Evolutionary relation-ships in AspergillussectionFumigatiinferred from partial-tubulin and hydrophobin DNA sequences. Mycologia90:831–845.
9.Guarro, J., J. Gene´, and A. M. Stchigel.1999. Developments in fungal taxonomy. Clin. Microbiol. Rev.12:454–500.
10.Hong, S. B., S. J. Go, H. D. Shin, J. C. Frisvad, and R. A. Samson.2005. Polyphasic taxonomy ofAspergillus fumigatusand related species. Mycologia 97:1316–1329.
11.Hong, S. B., H. D. Shin, J. Hong, J. C. Frisvad, P. V. Nielsen, J. Varga, and R. A. Samson.2008. New taxa ofNeosartoryaandAspergillusinAspergillus
sectionFumigati. Antonie van Leeuwenhoek93:87–98.
12.Jarv, H., J. Lehtmaa, R. C. Summerbell, E. S. Hoekstra, R. A. Samson, and P. Naaber.2004. Isolation ofNeosartorya pseudofischerifrom blood: first hint of pulmonary aspergillosis. J. Clin. Microbiol.42:925–928.
13.Jin, J., Y. K. Lee, and B. L. Wickes.2004. Simple chemical extraction method for DNA isolation fromAspergillus fumigatusand otherAspergillusspecies. J. Clin. Microbiol.42:4293–4296.
14.Katz, M. E., A. M. Dougall, K. Weeks, and B. F. Cheetham.2005. Multiple genetically distinct groups revealed among clinical isolates identified as atyp-icalAspergillus fumigatus. J. Clin. Microbiol.43:551–555.
15.Latge, J. P.1999.Aspergillus fumigatusand aspergillosis. Clin. Microbiol. Rev.12:310–350.
16.Peterson, S. W.2008. Phylogenetic analysis ofAspergillusspecies using DNA sequences from four loci. Mycologia100:205–226.
17.Sambrook, J., E. Fritsch, and T. Maniatis.1989. Molecular cloning: a lab-oratory manual, 2nd ed. Cold Spring Harbor Lablab-oratory Press, Cold Spring Harbor, NY.
18.Yaguchi, T., Y. Horie, R. Tanaka, T. Matsuzawa, J. Ito, and K. Nishimura. 2007. Molecular phylogenetics of multiple genes onAspergillussection
Fumigatiisolated from clinical specimens in Japan. Nippon Ishinkin Gakkai Zasshi48:37–46.