of Medically Important Black Yeast Species
Guido Heinrichs,aG. Sybren de Hoog,band Gerhard Haasea
Institut für Medizinische Mikrobiologie, RWTH Aachen University Hospital, Aachen, Germany,aand Centraalbureau voor Schimmelcultures KNAW Fungal Biodiversity
Centre, Utrecht, and Institute of Biodiversity and Ecosystem Dynamics, Amsterdam, The Netherlandsb
Herpotrichiellaceous black yeasts and relatives comprise severe pathogens flanked by nonpathogenic environmental siblings. Reliable identification by conventional methods is notoriously difficult. Molecular identification is hampered by the sequence variability in the internal transcribed spacer (ITS) domain caused by difficult-to-sequence homopolymeric regions and by poor taxonomic attribution of sequences deposited in GenBank. Here, we present a potential solution using short barcode identifiers (27 to 50 bp) based on ITS2 ribosomal DNA (rDNA), which allows unambiguous definition of species-specific fragments. Start-ing from proven sequences of ex-type and authentic strains, we were able to describe 103 identifiers. Multiple BLAST searches of these proposed barcode identifiers in GenBank revealed uniqueness for 100 taxonomic entities, whereas the three remaining identifiers each matched with two entities, but the species of these identifiers could easily be discriminated by differences in the remaining ITS regions. Using the proposed barcode identifiers, a 4.1-fold increase of 100% matches in GenBank was achieved in comparison to the classical approach using the complete ITS sequences. The proposed barcode identifiers will be made
accessible for the diagnostic laboratory in a permanently updated online database, thereby providing a highly practical, reliable, and cost-effective tool for identification of clinically important black yeasts and relatives.
B
lack yeasts and their filamentous relatives that belong to the ascomycete familyHerpotrichiellaceae(orderChaetothyriales) comprise a large group of pathogenic fungi. The infections caused by these fungi frequently occur in immunocompetent humans and range from mild cutaneous to severe, deep, or disseminated. The mycoses are difficult to treat, and systemic cases have a high fatality rate. For example,Cladophialophora bantianaand Rhino-cladiella mackenzieiare neurotropic agents classified as biosafety level 3 organisms (5). Some neurotropic species, likeExophiala dermatitidis, are common in wet habitats of indoor environments, such as in bathrooms (14), in steam baths (15), and on the rubber seals of dishwashers (19). Other black yeast species are associated with infections in cold-blooded animals (6). In total, the order contains 44 species that have been proven to be involved in infec-tions of vertebrate hosts (5) and ranges among the most species-rich clinically relevant groups in the fungal kingdom.Identification of black yeasts by morphological criteria alone is mostly impossible because of variable expression of morphologi-cal traits (synanamorphs) during complex, pleomorphic life cy-cles or low degrees of differentiation. Conversely, nearly identical morphological structures can be observed in phylogenetically dis-tantly related species (20). Since comparable physiological data are also lacking for most species and test results may be variable within the species, these traits do not allow reliable species iden-tification either.
Genetic identification of herpotrichiellaceous black yeasts is rou-tinely based upon sequences of the ribosomal DNA (rDNA) internal transcribed spacer (ITS) followed by a similarity search in public da-tabases. However, one should keep in mind that GenBank was not designed for this purpose. Nilsson et al. (16) noted that 27% of the ITS sequences deposited in the International Nucleotide Sequence Database (INSD; GenBank, EMBL, and DDBJ) had been incorrectly annotated to the species level, partly due to a general neglect of type materials defining the species. The need for a fast, easy, and reliable identification tool can be best illustrated by the sibling species
Clado-phialophora bantianaandCladophialophora psammophila:C. banti-anais an agent of fatal brain disease, whereasC. psammophilais as-sociated with aromatic hydrocarbons, proven to be avirulent in animal tests (1). With the molecular developments over the last de-cade, increasing numbers of novel species and reclassifications have been proposed, among which were numerous opportunistic patho-gens. In principal, usage of ITS sequence data is reliable for identifi-cation (20). Reference sequences have barcode status, i.e., sequences are open access and have been deposited with trace data and eventu-ally with original specimens. Of note, obvious sequence variability occurring mainly in homopolymeric regions due to formation of stutter products in the PCR (4) may interfere with unambiguous attribution of query sequences to a given species. Short barcode frag-ments are therefore needed which are invariable within the species and show stable indels or differences in nucleotide composition, e.g., due to compensatory base exchanges with neighboring species. The selection of barcode fragments with such properties may provide an elegant and straightforward approach to identify herpotrichiella-ceous black yeasts and their relatives.
MATERIALS AND METHODS
Strains and culture conditions. Sequences of ex-type and authentic strains of species with clinical relevance and closely related environmental
species ofHerpotrichiellaceaewere chosen for analysis. In addition, species
were included that had repeatedly been isolated from clinical samples. This resulted in a selection of 68 strains (Table 1), which are preserved at the CBS-KNAW Fungal Biodiversity Centre, Utrecht, The Netherlands. In
Received1 March 2012Returned for modification29 March 2012
Accepted5 July 2012
Published ahead of print11 July 2012
Address correspondence to Gerhard Haase, [email protected]. Copyright © 2012, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JCM.00574-12
on May 16, 2020 by guest
http://jcm.asm.org/
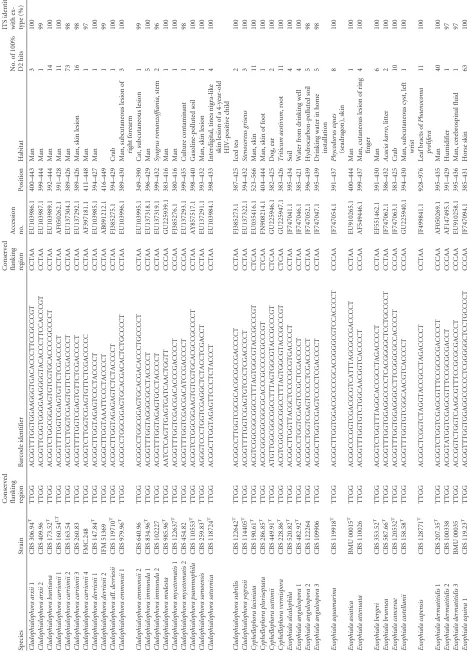
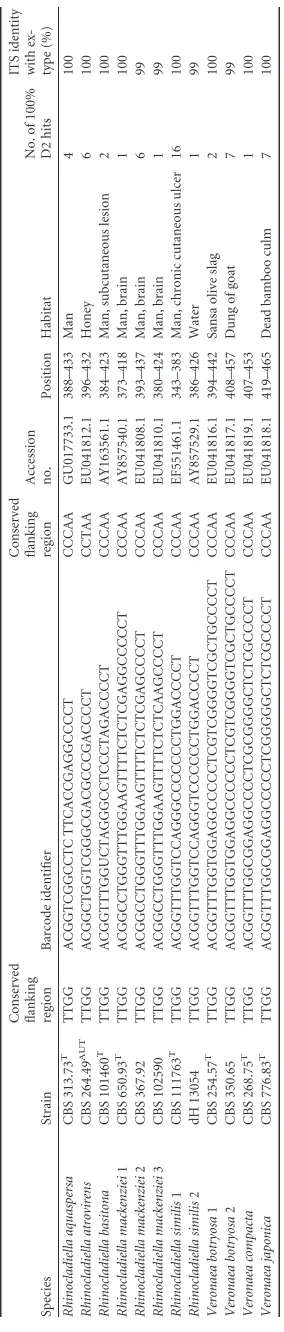
TABLE 1 Compilation and description of 103 proposed barcode identifiers Species Strain
Conserved flanking region
Barcode
identifier
Conserved flanking region Accession no. Position Habitat No. of 100% D2 hits ITS identity with ex-type (%) Cladophialophora arxii 1 CBS 306.94 T TTGG ACGGTTTGGTGGAAGGGTGACACCCTTCCGCCCGT CCTAA EU103986.1 400–443 Man 3 100 Cladophialophora arxii 2 CBS 409.96 TTGG ACGGTTCGGTGGGGAAGGGGTACACCCTTCCACCCGT CCTAA EU103987.1 399–444 Man 1 99 Cladophialophora bantiana CBS 173.52 T TTGG ACGGTCTGGCGGAAGTGTCGTGCACCCCGCCCCT CCTAA EU103989.1 392–444 Man 14 100 Cladophialophora carrionii 1 CBS 160.54 LT TTGG ACGGTTTTGGTCGAGTGTTCTCGACCCCT CCTAA AF050262.1 391–428 Man 11 100 Cladophialophora carrionii 2 CBS 163.54 TTGG ACGGTCTTGGTCGAGTGTTCTCGACCCCT CCTAA EU137304.1 389–426 Man 73 98 Cladophialophora carrionii 3 CBS 260.83 TTGG ACGGTTTTGGTCGAGTGTTCTCGACCCCT CCTAA EU137292.1 389–426 Man, skin lesion 16 98 Cladophialophora carrionii 4 FMC.248 TTGG ACGGTCTTGGTCGAGTGTTCTCGACCCCC CCTAA AF397181.1 411–448 Man 1 97 Cladophialophora devriesii 1 CBS 147.84 T TTGG ACGGCTTGGTAGAGTCCCTACCCCT CCTAA EU103985.1 394–427 Man 1 100 Cladophialophora devriesii 2 IFM 51369 TTGG ACGGCTTGGTAAATTCCCTACCCCT CCTAA AB091212.1 416–449 1 99 Cladophialophora aff. devriesii CBS 119710 T TTGG ACGGCTTGGTAGAGAGTCTCTACCCCT CCTAA FJ385275.1 394–429 Crab 1 100 Cladophialophora emmonsii 1 CBS 979.96 T TTGG ACGGCCTGGCGGAGTGCACGACACTCTGCCCCT CCTAA EU103996.1 389–430 Man, subcutaneous lesion of right forearm 3 100 Cladophialophora emmonsii 2 CBS 640.96 TTGG ACGGCCTGGCGGAGTGCACGACACCCTGCCCCT CCTAA EU103995.1 349–390 Cat, subcutaneous lesion 1 99 Cladophialophora immunda 1 CBS 834.96 T TTGG ACGGTTTGGTAGGGCGCCTACCCCT CCTAA EU137318.1 396–429 Man 5 100 Cladophialophora immunda 2 CBS 102227 TTGG ACGGTTTGGTAGAGTGCCTACCCCT CCTAA EU137319.1 399–432 Syagrus romanzoffiania ,stem 2 96 Cladophialophora modesta CBS 985.96 T TTGG AATCTCAGTTGAGTGATCAACTGGTT CCCAA GU225939.1 383–416 Man 1 100 Cladophialophora mycetomatis 1 CBS 122637 T TTGG ACGGTTTGGTCGACGACACCCGACCCCT CCCAA FJ385276.1 380–416 Man 1 100 Cladophialophora mycetomatis 2 CBS 454.82 TTGG ACGGTTTGGTCGACGACATCCGACCCCT CCCAA EU137293.1 399–435 Culture contaminant 1 98 Cladophialophora psammophila CBS 110553 T TTGG ACGGTCTGGCGGAAGTGTCGTGCACGCCGCCCCT CCTAA AY857517.1 398–440 Gasoline-polluted soil 1 100 Cladophialophora samoensis CBS 259.83 T TTGG AGGGTCCCTGGTCGAGGCTCTACCTCGACCT CCTAA EU137291.1 393–432 Man, skin lesion 1 100 Cladophialophora saturnica CBS 118724 T TTGG ACGGCTTGGTAGAGTTCCCTCTACCCT CCTAA EU103984.1 398–433 Interdigital, tinea nigra-like skin lesion of a 4-year-old HIV-positive child 4 100 Cladophialophora subtilis CBS 122642 T TTGG ACGGCCTTGGTCGCGCACGCGCCGACCCCT CCTAA FJ385273.1 387–425 Iced tea 2 100 Cladophialophora yegresii CBS 114405 T TTGG ACGGTTTTGGTCGAGTGTCCCTCGACCCCT CCTAA EU137322.1 394–432 Stenocereus griseus 3 100 Cyphellophora laciniata CBS 190.61 T TTGG ACGTCGGCGGCGGCCTTTAGTGGCGTACCGCCCGT CTCAA EU035416.1 523–566 Man, skin 11 100 Cyphellophora pluriseptata CBS 286.85 T TTGG ACGCGGCGGCGGCGCACCCGCCCCCGCCCGT CTCAA FN908214.1 404–444 Man, skin of foot 1 100 Cyphellophora suttonii CBS 449.91 T TTGG ATGTTGGCGGCGGCCTTTAGTGGCGTACCGCCCGT CTCAA GU225946.1 382–425 Dog, ear 2 100 Cyphellophora vermispora CBS 228.86 T TTGG ACGTCGGCGGCGGCCTTTAGTGGCGTACCGCCCGT CTCAA GU225947.1 383–426 Triticum aestivum ,root 11 100 Exophiala alcalophila CBS 520.82 T TTGG ACGGCTTGGTTACGCTCCCCGCGTGACCCCT CCTAA JF747041.1 395–434 Soil 4 100 Exophiala angulospora 1 CBS 482.92 T TTGG ACGGCCTGGTCGAGTCCGCTCGACCCCT CCTAA JF747046.1 385–421 Water from drinking well 4 100 Exophiala angulospora 2 CBS 122264 TTGG ACGGCCTGGTCGAGTCCCCTCGACCCCT CCTAA JF747052.1 396–439 Hydrocarbon-polluted soil 5 98 Exophiala angulospora 3 CBS 109906 TTGG ACGGCTTGGTCGAGTCCCCTCGACCCCT CCTAA JF747047.1 395–439 Drinking water in home installation 59 8 Exophiala aquamarina CBS 119918 T TTGG ACGGCTTGGTGGACGCCCCGCACGGGGCGTCCACCCCT CCCAA JF747054.1 391–437 Phycodurus eques (seadragon), skin 8 100 Exophiala asiatica BMU 00015 T TTGG ACGGCCCGGTCGACGCGTCATTTAGCGCCGACCCCT CCTAA EU910265.1 400–444 Man 1 100 Exophiala attenuata CBS 110026 TTGG ACGGTTTGGTGTCTGGCAACGGTCACCCCT CCCAA AF549446.1 399–437 Man, cutaneous lesion of ring finger 4 100 Exophiala bergeri CBS 353.52 T TTGG ACGGTCTGGTTTAGGCACCGCCTAGACCCCT CCTAA EF551462.1 391–430 Man 6 100 Exophiala brunnea CBS 587.66 T TTGG ACGGTTTGGTGGAGGCCCCTCACGGGGCTCCTGCCCCT CCTAA JF747062.1 386–432 Acacia karro ,litter 2 100 Exophiala cancerae CBS 120532 T TTGG ACGGTTTGGTGGAGGGCCTTCGGGCGCCACCCCT CCCAA JF747063.1 383–425 Crab 10 100 Exophiala castellanii CBS 158.58 T TTGG ACGGTTTGGTGTCGGCAACGTCACCCCT CCCAA GU225940.1 394–430 Man, subcutaneous cyst, left wrist 1 100 Exophiala capensis CBS 128771 T TTGG ACGGCTCGGTCTAGGTACCGCCTAGACCCCT CCTAA JF499841.1 928–976 Leaf bracts of Phaenocoma prolifera 11 100 Exophiala dermatitidis 1 CBS 207.35 T TTGG ACGGTCTGGTCGAGCGTTTCCGCGCGACCCCT CCCAA AF050269.1 395–435 Man 40 100 Exophiala dermatitidis 2 CBS 100338 TTGG ACGGTATGGTCGAGCGTTTCCGCGCGACCT CCCAA AF147495.1 391–429 Humidifier 1 97 Exophiala dermatitidis 3 BMU 00035 TTGG ACCGGTCTGGTCAAGCGTTTCCGCGCGACCCT CCCAA EU910258.1 395–436 Man, cerebrospinal fluid 1 97 Exophiala equina 1 CBS 119.23 T TTGG ACGGTTTGGTGGAGGCCCCCTCGGGGGCTCCTGCCCCT CCCAA JF747094.1 385–431 Horse skin 63 100
on May 16, 2020 by guest
http://jcm.asm.org/
[image:2.585.63.532.72.727.2]Exophiala equina 2 CBS 122263 TTGG ACGGTTTGGTGGAGGCCCCCTCGGGGGTTCCTGCCCCT CCCAA JF747099.1 385–431 9 98 Exophiala equina 3 CBS 122270 TTGG ACGGTTTGGTGGAGGTCCCCTCGGGGGCTCCTGCCCCT CCCAA JF747101.1 384–430 Human foot 9 98 Exophiala exophialae CBS 668.76 T TTGG ACGGTTTGGTCCCGGGACGCCCCTGGACCCCT CCCAA AY156973.1 385–425 Straw in burrow of Dasypus septemcinctus 3 100 Exophiala halophila CBS 121512 T TTGG ACGGCTTGGTCGCGCCCCCCGGGCACCGACCCCT CCTAA JF747108.1 394–436 Man, axillary 3 100 Exophiala heteromorpha CBS 232.33 T TTGG ACGGTCTGGTCGAGCCCGCCTCGACCCCT CCCAA AY857524.1 389–426 Wood pulp 6 100 Exophiala jeanselmei 1 CBS 507.90 T TTGG ACGGTTTGGTCTCGGGTCCGACCCCCCTTGACCCCT CCCAA AY156963.1 389–433 Man 14 100 Exophiala jeanselmei 2 CBS 677.76 TTGG ACGGTTTGGTCAAAGGGTCCGACCCCCCTAGACCCCT CCCAA AY163553.1 386–431 Skin, abscess of foot 2 96 Exophiala lacus CBS 117497 T TTGG ACGGTTTGGCGGAGACCTGTTACAGGCCTCCACCCCT CCCAA JF747110.1 384–429 Water 1 100 Exophiala lecanii-corni CBS 123.33 T TTGG ACGGCCTGGCGTCGGCGACGACCCCACCT CCCAA AY857528.1 409–446 Man 10 100 Exophiala mesophila 1 CBS 402.95 T TTGG ACGGCTTGGTGTCAGCGATGTCACCCCT CCTAA GU225941.1 396–432 Silicone seal, in shower room of hospital 16 100 Exophiala mesophila 2 CBS 121511 TTGG ACGGCTTGGTGTCAGCAATGTCACCCCT CCTAA JF747122.1 397–433 Nasal tissue 1 97 Exophiala moniliae CBS 520.76 T TTGG ACGGTCTGGTTAGGCGACTGACCCCT CCTAA GU225948.1 390–424 Quercus ,twig 6 100 Exophiala nishimurae CBS 101538 T TTGG ACGGCTTGGTCTAGGTCGTCGCCTAGACCCCT CCCAA AY156966.1 a 392–432 Bark 1 100 Exophiala oligosperma 1 CBS 725.88 T TTGG ACGGTCTGGTCCGGGGACCTCAAACCCCCTGGACCCCT CCCAA AY163551.1 397–443 Man, tumor of sphenoidal cavity 25 100 Exophiala oligosperma 2 UTHSC95-2041 TTGG ACGGTTTGGTTCGGGGACCTCAAACCCCCTGGACCCCT CCCAA EF025415.1 388–434 Man 2 96 Exophiala oligosperma 3 UTHSC 91–870 TTGG ACGGTTTGGTCCGGGGACCTCAAAGCCCCCTGGACCCCT CCCAA EF551458.1 354–401 Man, hand 8 99 Exophiala oligosperma 4 IFM 41701 TTGG ACGGTTTGGTCCGGGGACCTCAAAAACCCCCTGGACCCCT CCCAA AY163548.1 392–440 Soil 1 97 Exophiala opportunistica CBS 109811 TTGG ACGGTTTGGTGGAGACCCCCTTCGCGGGCTTCTACCCCT CCCAA JF747123.1 384–431 Drinking water 1 100 Exophiala pisciphila CBS 537.73 T TTGG ACGGTTTGGTGGAGGCCCCCTCGCGGGGCCACCGCCCCT CCCAA AF050272.1 390–437 Ictalurus punctatus 17 100 Exophiala salmonis CBS 157.67 T TTGG ACGGTTTGGTGGAGGCCCCTTGCGGGCATCTGCCCCT CCCAA AF050274.1 512–557 Salmo clarkii 4 100 Exophiala sideris CBS 121818 T TTGG ACGGTTTGGTCCAGGTCACCTGGACCCCT CCTAA HQ452311.1 391–428 Berry, Sorbus aucuparia 26 100 Exophiala spinifera CBS 899.68 T TTGG ACGGTTTGGTCCCGGGACCCCCCTGGACCCCT CCCAA AY156976.2 384–424 Man, nasal granuloma 37 100 Exophiala “siphonis” UTHSC88-471 TTGG ACGGTCTGGTCGAGCTGCTCGACCCCT CCCAA EF025401.1 391–426 Man 6 100 Exophiala xenobiotica 1 CBS 118157 T TTGG ACGGTTTGGTTTAGGTACCCCTAGACCCCT CCTAA DQ182587.1 383–421 Oil sludge 32 100 Exophiala xenobiotica 2 CBS 119306 TTGG ACGGTTTGGTTTAGGCACCCCTAGACCCCT CCTAA EF025409.1 377–415 Man 1 97 Exophiala xenobiotica 3 CBS 117665 TTGG ACGGTTTGGTTTAGGTACCCCTAGATCCCT CCTAA DQ182588.1 383–421 Man, tissue 7 99 Exophiala xenobiotica 4 CBS 117676 TTGG AGGTCTTGGTGTAGGGTTCCCCCCTCCACCCCT CCTAA DQ182592.1 388–429 Man, finger 1 93 Exophiala xenobiotica 5 CBS 117641 TTGG AGGTCTTGGTGTAGGGTCCCCCCTACACCCCT CCTAA DQ182591.1 388–428 Man, knee cyst 1 94 Fonsecaea erecta 1 dH 20513 TTGG ACGGTCCGGTGGAGAGTCATCCTTTCCACCCGT CCTAA 391–432 1 100 Fonsecaea erecta 2 dH 20502 TTGG ACGGTCCGGTGGAGAGTCATCCTTTTCACCCGT CCTAA 428–469 0 b 99 Fonsecaea minima dH 20511 TTGG ACGGTTCGGTGGAGAGTCACACCTCCCACCCGT CCTAA 396–437 2 100 Fonsecaea monophora 1 CBS 269.37 T TTGG ACGGCTTGGTGGAGTAAGTTCACACTTTCCACCCCT CCTAA AY857511.1 401–445 Man 20 100 Fonsecaea monophora 2 CBS 121732 TTGG ACGGCTTGGTGGAGCAAGTTCACACTTTCCACCCCT CCTAA EF513759.1 400–444 Man 20 99 Fonsecaea monophora 3 IFM 4889 TTGG ACGGCTTGGTGGAGTAAGTTCACGCTTTCCACCCCT CCTAA AB091204.2 423–467 Man 33 97 Fonsecaea monophora 4 IFM 54446 TTGG ACGGCTTGGTGGAGTAGGTTCACGCTTTCCACCCCT CCTAA AB251881.1 400–444 Cervical lymph nodes 1 97 Fonsecaea multimorphosa CBS 980.96 T TTGG ACGGCTCGGTGGACTCCTTCCACCCGT CCTAA JF267657.1 392–427 Cat 0 b 100 Fonsecaea nubica 1 CBS 269.64 T TTGG ACGGTTTGGTGGAGCGAGTTCACACTTTCCACCCCT CCTAA EU938592.1 400–444 Man, skin 6 100 Fonsecaea nubica 2 CBS 121733 TTGG ACGGCTTGGTGGAGCGAGGTCACACTTTCCACCCCT CCTAA EF513756.1 447–491 Man 2 98 Fonsecaea nubica 3 CBS 557.76 TTGG ACGGCTTGGTGGAGCGAGTTCACACTTTCCACCCCT CCTAA EU938593.1 399–443 52 99 Fonsecaea pedrosoi 1 CBS 271.37 T TTGG ACGGCTTGGTGGAGCGAGTTCACACTTTCCACCCCT CCTAA AB114127.1 423–467 Man 52 100 Fonsecaea pedrosoi 2 CBS 122741 TTGG ACGGCTTGGTGGAGCGAGTTCACACTTTCCACCCCCT CCTAA EU938589.1 400–445 Man, foot 1 98 Knufia epidermidis CBS 120353 T TTGG ACCCAAGTTTTTGGCTATTAAAAACTTGGT CCTAA EU730589.1 392–430 Man, foot 2 100 Phaeoannelomyces elegans CBS 110172 TTGG ACGGTTTGGTGGAGGCCCCCTCGGGGGCTCCTGCCCCT CCCAA EF551549.1 348–394 Railway tie 63 100 Phialophora americana CBS 840.69 TTGG ACGGATTTTGGTCGTGTAACAACGACCCCT CCTAA AF050283.1 387–425 Decaying timber 14 100 Phialophora europaea CBS 129.96 T TTGT AGCGGCCCCCCCGCTCTT CCCAA EF551553.1 386–412 Man, toe 12 100 Phialophora reptans CBS 113.85 T TTGG ACCCGGCGGGAGCTTCGCGCCCCCGCCCGT CTCAA EU514699.1 389–427 4 100 Phialophora verrucosa 1 CBS 273.37 TTGG ACGGATCTTGGTCGTGTGATCGCGACCCCT CCTAA AF050281.1 389–427 Man 2 100 Phialophora verrucosa 2 IMTSP.800 TTGG ACGGATCCTGGTCGTGTGATCGCGACCCCT CCTAA AF397135.1 410–448 1 99 Phialophora verrucosa 3 CBS 286.47 TTGG ACGGATTTTGGTCGTGTGATATCGACCCCT CCTAA AF050282.1 397–429 Fruit rachis, Musa sapientum 59 6 Phialophora verrucosa 4 CBS 839.68 TTGG ACGGATTTTGGTCGTGTGATACCGACCCCT CCTAA DQ404353.1 434–472 Kiwifruit elephantiasis 2 98 Rhinocladiella anceps 1 CBS 181.65 INT TTGG AAGGCCTGGTGTCCCCCCGGGGACACCCCT CCCAA EU041805.1 385–423 Soil under Thuja plicata 2 100 Rhinocladiella anceps 2 CBS 157.54 TTGG AAGGCCTGGTGTCTCC CCCCGGGGATACCCCT CCCAA EU041804.1 417–457 Fagus sylvatica ,stem 4 95 (Continued on following page)
on May 16, 2020 by guest
http://jcm.asm.org/
case of missing sequences, sequencing was performed. All sequences of the complete ITS regions of these strains have been deposited in GenBank. Fungi were cultured in 5 ml 2% Sabouraud’s glucose broth (Merck, Darmstadt, Germany) in cultivation tubes (Sarstedt, Nümbrecht, Ger-many) for at least 5 days at 28°C.
DNA extraction, PCR, sequencing.Tubes were centrifuged for 10
min at 3,180⫻gat 4°C. Aliquots of about 0.5 g were removed and washed
with 500l aqua dest. in 2-ml cups. One hundred microliters of lysis
buffer (50 mM Tris HCl [pH 7.2], 50 mM EDTA, 3% SDS) was added, and the biomass was crushed with a micropestle for 1 min. The cups were
incubated at 99°C for 2 min, 300l lysis buffer was added, and cups were
incubated at 80°C for an additional 10 min. After extraction with 400l
extraction mix (50:48:2 [percent volume]
phenol-chloroform-isoamylal-cohol) and 20 min of centrifugation at 20,000⫻g, DNA was precipitated
from the aqueous phase with 40l 3 M sodium acetate (pH 5.2) and 400
l isopropanol and separated by centrifugation with 20,000⫻gfor
an-other 20 min. The pellet was washed with 70% ethanol and dissolved in 50
l aqua dest. PCR was performed with 500 ng target DNA, ITS5 (5=-GG
AAGTAAAAGTCGTAACAAGG-3=) and NL-4 (5=-GGTCCGTGTTTCA
AGACGG-3=) as primers (Sigma, Taufkirchen, Germany), and
Taq-Poly-merase (Roche, Mannheim, Germany) using the following program: 94°C initially for 1 min, 55°C for 1 min, and 72°C for 3 min for 35 cycles, with a terminal elongation step at 72°C for 5 min (Labcycler, Sensoquest, Göt-tingen, Germany). Sequencing was done with 200 ng PCR product on an ABI Prism 310 Genetic Analyzer (Applied Biosystems, Delaware) with ITS5 and NL-4 as sequencing primers.
BLAST searches and parameters.BLAST searches (BLASTN 2.2.25⫹) were performed at the NCBI website with the (i) 68 barcode identifier candidate sequences (range, 27 to 50 bp; strategy A), (ii) ITS2 sequence segments (range, 183 to 249 bp; strategy B), and (iii) complete ITS1, 5.8S, and ITS2 sequence segments (range, 477 to 595 bp; strategy C) using the GenBank database as of 31 October 2011. For all three search strategies, BLAST algorithm parameters were set as follows. General parameters were as follows: max target sequences, 100; short queries, automatically adjust parameters for short input sequences; expect threshold, 10; word size, 28. Scoring parameters were as follows: match/mismatch scores, 1,
⫺2; gap costs, linear. Filters and masking were as follows: filter, low
com-plexity regions; mask, mask for lookup table only. The search was re-stricted to fungi (taxid 4751). Revealed sequences with less than 100% query coverage were not taken into account.
RESULTS
Determination of the barcode candidates.ITS sequences (n⫽
68) were downloaded and processed using BioEdit (version 7.0.9.1) (9). The rDNA spacer ITS1 and ITS2 regions were delim-ited manually using a Web-based tool (11). Sequences were veri-fied for suitable regions meeting the requirements of a barcode fragment with low intraspecies variability and with a significant gap toward the neighboring species. This fragment is termed a “barcode identifier” since it contains only the signature sequence (hypervariable region) (10) of the barcode since the term “bar-code” is typically used in conjunction with an entire gene. It be-came evident that domain two (13), located in the ITS2 rDNA, was a promising candidate for this purpose. Within the Herpotrichiel-laceae, this region was 27 to 50 bp in length, had highly conserved flanking regions at the 5=and 3=ends (Table 1), and showed great variability in the central portion, even in case of closely related taxa. Therefore, this region was chosen for further analysis on the basis of assumed high interspecies and low intraspecies variability. To detect intraspecies variation within each of the 68 entities, a search with every species name as well as respective synonyms was performed in GenBank with restriction to internal transcribed spacer sequences. Resulting sequences covering at least 85% of the
TABLE
1
(Continued)
Species
Strain
Conserved flanking region
Barcode
identifier
Conserved flanking region Accession no.
Position
Habitat
No.
of
100%
D2
hits
ITS
identity
with
ex-type
(%)
Rhinocladiella
aquaspersa
CBS
313.73
T
TTGG
ACGGTCGGCCTC
TTCACCGAGGCCCCT
CCCAA
GU017733.1
388–433
Man
4
100
Rhinocladiella
atrovirens
CBS
264.49
AUT
TTGG
ACGGCTGGTCGGGCGACGCCCGACCCCT
CCTAA
EU041812.1
396–432
Honey
6
100
Rhinocladiella
basitona
CBS
101460
T
TTGG
ACGGTTTGGUCTAGGGCCTCCCTAGACCCCT
CCCAA
AY163561.1
384–423
Man,
subcutaneous
lesion
2
100
Rhinocladiella
mackenziei
1
CBS
650.93
T
TTGG
ACGGCCTGGGTTTGGAAGTTTTCTCTCGAGGCCCCCT
CCCAA
AY857540.1
373–418
Man,
brain
1
100
Rhinocladiella
mackenziei
2
CBS
367.92
TTGG
ACGGCCTGGGTTTGGAAGTTTTCTCTCGAGCCCCT
CCCAA
EU041808.1
393–437
Man,
brain
6
99
Rhinocladiella
mackenziei
3
CBS
102590
TTGG
ACGGCCTGGGTTTGGAAGTTTTCTCTCAAGCCCCT
CCCAA
EU041810.1
380–424
Man,
brain
1
99
Rhinocladiella
similis
1
CBS
111763
T
TTGG
ACGGTTTGGTCCAGGGCCCCCCCTGGACCCCT
CCCAA
EF551461.1
343–383
Man,
chronic
cutaneous
ulcer
16
100
Rhinocladiella
similis
2
dH
13054
TTGG
ACGGTTTGGTCCAGGGTCCCCCCTGGACCCCT
CCCAA
AY857529.1
386–426
Water
1
99
Veronaea
botryosa
1
CBS
254.57
T
TTGG
ACGGTTTGGTGGAGGCCCCCTCGTCGGGGTCGCTGCCCCT
CCCAA
EU041816.1
394–442
Sansa
olive
slag
2
100
Veronaea
botryosa
2
CBS
350.65
TTGG
ACGGTTTGGTGGAGGCCCCCCTCGTCGGGGTCGCTGCCCCT
CCCAA
EU041817.1
408–457
Dung
of
goat
7
99
Veronaea
compacta
CBS
268.75
T
TTGG
ACGGTTTGGCGGAGGCCCCTCGCGGGGCTCTCGCCCCT
CCCAA
EU041819.1
407–453
1
100
Veronaea
japonica
CBS
776.83
T
TTGG
ACGGTTTGGCGGAGGCCCCCTCGGGGGCTCTCGCCCCT
CCCAA
EU041818.1
419–465
Dead
bamboo
culm
7
100
aDeposited
sequence
belongs
to
strain
CBS
110628.
bSequence
not
deposited
at
the
time
of
analysis.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:4.585.90.232.64.730.2]length of the respective ex-type strain’s entire ITS region were added to the initial sequence alignment. Deviating barcodes with the same species denotation (i.e., proven to be conspecific in the literature by genealogical concordance analysis) and exhibiting
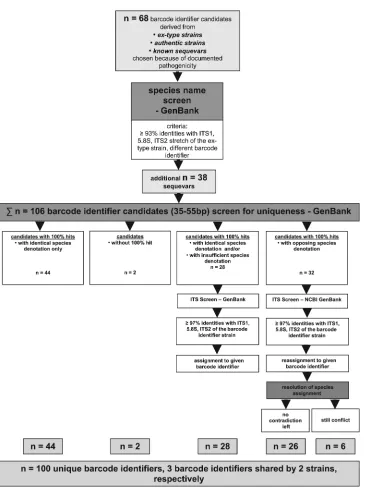
ⱖ93% sequence identity (Table 1) in the remaining ITS region of the specific ex-type strain were treated as sequevars of the respec-tive species. The original ex-type or authentic strain’s name was then denoted with “1,” and all following sequevars were indicated by increasing numbers according to decreasing sequence similar-ity of the identifier region (e.g.,E. dermatitidis,Table 1). For 22 of the original 68 barcode identifiers, 38 tentative sequevars (2 to 5 per species) were revealed, resulting in a total of 106 barcode iden-tifier candidates (Table 1). A flow diagram of the procedure of barcode identifier selection is given inFig. 1.
Screen for uniqueness of the barcode identifiers.An NCBI
BLAST search with the barcode identifier candidates (n⫽106) of 27 to 50 bp in length (search strategy A) was performed using the parameters described above. Only hits with 100% query coverage were considered for further analysis. In 44 cases, the barcode identifier was found to be unique, providing only a single species denotation. In 28 cases, 100% hits with insuffi-ciently identified source organisms (e.g., “Exophiala unde-scribed species” or “uncultured ascomycete”) were obtained. Since all these exhibitedⱖ97% sequence identity of the entire ITS region with the ex-type isolate of the respective black yeast species, they were assigned to the species name of the particular barcode identifier. In two cases, viz. Fonsecaea erecta 1 and Fonsecaea multimorphosa, no match could be obtained because FIG 1Determination and analysis of the barcode identifiers.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:5.585.113.478.58.550.2]ITS sequences of these novel species had not been deposited in GenBank at the time of analysis.
In 29 cases, 100% hits were obtained with source organisms other than the species defined by the ex-type isolate. This was reevaluated by a thorough search of recent literature with eventual taxonomic novelties. All but three conflicts could be traced back to obsolete or simply erroneous species denotations of the respective GenBank entries, which had not been updated or corrected. In those cases, respective sequences were reassigned to the particular species or sequevars. The three remaining conflicting sequences with different species sharing the same barcode identifier were compared with the entire ITS domain. Of note,Cyphellophora laciniataandCyphellophora vermisporacould be distinguished by 1.3% ITS diversity,Fonsecaea pedrosoisequevar 1 andFonsecaea nubicasequevar 3 by 2.6%, andExophiala equinasequevar 1 and Phaeoannellomyces elegansby 13.0%, with differences occurring mainly in the ITS1 region.Figure 1illustrates the described ap-proach which led to 100 unique barcode identifiers and 3 barcode identifiers shared by two species or sequevars each (n⫽106 initial barcode identifier candidates).
Performance of BLAST search strategies. Three BLAST
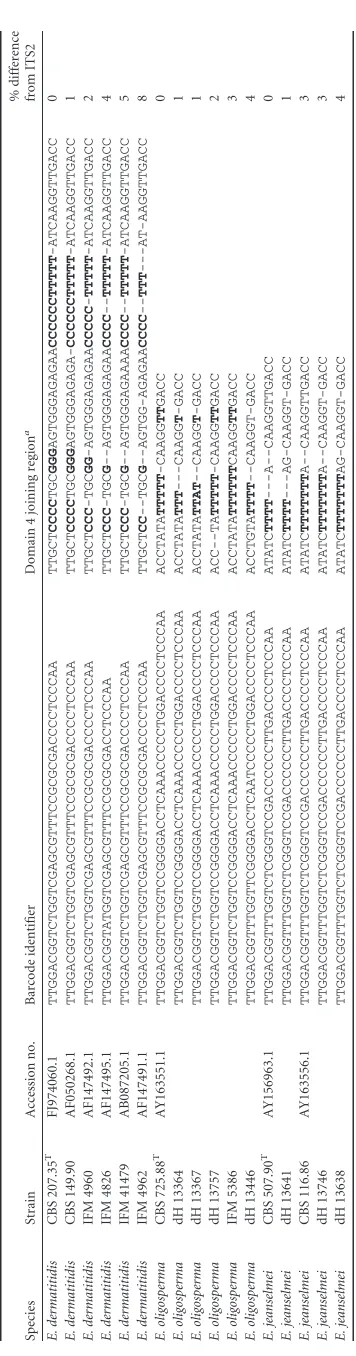
searches were performed with the 103 barcode identifiers (strategy A), the ITS2 (strategy B), and the near-complete ITS domain (strategy C) using GenBank with the settings as described above. The first and second best hits were documented (data not shown). BLAST searches A, B, and C were compared for resulting 100% sequence matches. When the barcode identifiers (strategy A) were used, about four times more 100% hits were found than with the entire ITS domain (strategy C), underlining the diagnostic strength of the use of barcode identifiers. With increasing length of the fragment used in a BLAST search, the total number of 100% hits with concordant species names decreased, as well as the total number of 100% hits with ambiguous or inconsistent species names (A¡B¡C). Detailed values are given inTable 2. This phe-nomenon is due to accumulating sequence errors in longer se-quences occurring mainly in homopolymeric regions due to for-mation of stutter products in the PCR (4) (Table 3).
DISCUSSION
[image:6.585.334.512.52.730.2]Black yeast fungi and their filamentous relatives in the Herpo-trichiellaceaecomprise harmless environmental species, some of which are closely related to severely pathogenic biosafety level 3 organisms (1). Pathogens and saprobes cannot be discriminated by conventional methods using micromorphology or physiology. A genetic approach using the ITS domain deposited in public databases for sequence analysis is regarded as the gold standard for routine identification, even though this may be hampered by poor quality of databases in terms of sequence quality, sequence length, TABLE 2Number of 100% sequence matches resulting from three different BLAST search strategies
BLAST search
strategy Segment blasted
Total no. of 100% matches
Total no. of 100% hits with concordant species name
Total no. of 100% hits with ambiguous or inconsistent species name
A Barcode identifier 944 496 448
B ITS2 410 243 167
C ITS1, 5.8S, ITS2 231 140 91
TABLE
3
Examples
of
homoploymeric
regions
in
the
domain
4
joining
fragment
and
the
resulting
sequence
divergence
referring
to
the
respective
ex-type
strain
of
three
Exophiala
species
Species
Strain
Accession
no.
Barcode
identifier
Domain
4
joining
region
a
%
difference
from
ITS2
E.
dermatitidis
CBS
207.35
T
FJ974060.1
TTGGACGGTCTGGTCGAGCGTTTCCGCGCGACCCCTCCCAA
TTGCT
CCCC
TGC
GGG
AGTGGGAGAGAA
CCCCCCTTTTT
-ATCAAGGTTGACC
0
E.
dermatitidis
CBS
149.90
AF050268.1
TTGGACGGTCTGGTCGAGCGTTTCCGCGCGACCCCTCCCAA
TTGCT
CCCC
TGC
GGG
AGTGGGAGAGA-CCCCCCTTTTT
-ATCAAGGTTGACC
1
E.
dermatitidis
IFM
4960
AF147492.1
TTGGACGGTCTGGTCGAGCGTTTCCGCGCGACCCCTCCCAA
TTGCT
CCC
-TGC
GG
-AGTGGGAGAGAA
CCCCC
-TTTTT
-ATCAAGGTTGACC
2
E.
dermatitidis
IFM
4826
AF147495.1
TTGGACGGTATGGTCGAGCGTTTCCGCGCGACCTCCCAA
TTGCT
CCC
-TGC
G
--AGTGGGAGAGAA
CCCC
--TTTTT
-ATCAAGGTTGACC
4
E.
dermatitidis
IFM
41479
AB087205.1
TTGGACGGTCTGGTCGAGCGTTTCCGCGCGACCCCTCCCAA
TTGCT
CCC
-TGC
G
--AGTGGGAGAAAA
CCCC
--TTTTT
-ATCAAGGTTGACC
5
E.
dermatitidis
IFM
4962
AF147491.1
TTGGACGGTCTGGTCGAGCGTTTCCGCGCGACCCCTCCCAA
TTGCT
CC
--TGC
G
--AGTGG-AGAGAA
CCCC
--TTT
---AT-AAGGTTGACC
8
E.
oligosperma
CBS
725.88
T
AY163551.1
TTGGACGGTCTGGTCCGGGGACCTCAAACCCCCTGGACCCCTCCCAA
ACCTATA
TTTTT
-CAAGG
TT
GACC
0
E.
oligosperma
dH
13364
TTGGACGGTCTGGTCCGGGGACCTCAAACCCCCTGGACCCCTCCCAA
ACCTATA
TTT
---CAAGG
T
-GACC
1
E.
oligosperma
dH
13367
TTGGACGGTCTGGTCCGGGGACCTCAAACCCCCTGGACCCCTCCCAA
ACCTATA
TTAT
--CAAGG
T
-GACC
1
E.
oligosperma
dH
13757
TTGGACGGTCTGGTCCGGGGACCTCAAACCCCCTGGACCCCTCCCAA
ACC--TA
TTTTT
-CAAGG
TT
GACC
2
E.
oligosperma
IFM
5386
TTGGACGGTCTGGTCCGGGGACCTCAAACCCCCTGGACCCCTCCCAA
ACCTATA
TTTTTT
CAAGG
TT
GACC
3
E.
oligosperma
dH
13446
TTGGACGGTTTGGTTCGGGGACCTCAATCCCCCTGGACCCCTCCCAA
ACCTGTA
TTTT
--CAAGGT-GACC
4
E.
jeanselmei
CBS
507.90
T
AY156963.1
TTGGACGGTTTGGTCTCGGGTCCGACCCCCCTTGACCCCTCCCAA
ATATC
TTTT
---A--CAAGGTTGACC
0
E.
jeanselmei
dH
13641
TTGGACGGTTTGGTCTCGGGTCCGACCCCCCTTGACCCCTCCCAA
ATATC
TTTT
---AG-CAAGGT-GACC
1
E.
jeanselmei
CBS
116.86
AY163556.1
TTGGACGGTTTGGTCTCGGGTCCGACCCCCCTTGACCCCTCCCAA
ATATC
TTTTTTT
A--CAAGGTTGACC
3
E.
jeanselmei
dH
13746
TTGGACGGTTTGGTCTCGGGTCCGACCCCCCTTGACCCCTCCCAA
ATATC
TTTTTTT
A--CAAGGT-GACC
3
E.
jeanselmei
dH
13638
TTGGACGGTTTGGTCTCGGGTCCGACCCCCCTTGACCCCTCCCAA
ATATC
TTTTTTT
AG-CAAGGT-GACC
4
aBolding
represents
variable
homopolymeric
regions.
on May 16, 2020 by guest
http://jcm.asm.org/
and taxonomic or other updates. Nilsson et al. (16) found that more than a quarter of the ITS sequences in general databases had been incorrectly annotated; 2.5% of all sequences had more than 1% IUPAC ambiguities, 82% lacked an explicit reference to a voucher specimen, and 99.2% of all fungal sequences had never been updated after first submission. The databases are not and were not intended to be appropriate for the identification of spe-cies. Smaller, dedicated, and validated databases are increasingly implemented (e.g.,Fusarium, http://www.cbs.knaw.nl/fusarium
/BioloMICS.aspx;Trichoderma,http://www.isth.info/), but these
as yet cover only a fraction of the fungal kingdom. Furthermore, the ITS2 region of theHerpotrichiellaceaecomprises homopoly-meric regions, particularly poly(T), which may result in artificial sequence variance of up to 8% of base positions (Table 3). The barcode identifier approach eliminates these uncertainties effec-tively (Table 2).
To overcome the obstacles described above, we here present an identification approach based on barcode identifiers validated by comparison with ex-type strains. A 27- to 50-bp fragment located in the ITS2 is selected. We have shown that 100 out of 106 species or sequevars could be distinguished unequivocally using the pro-posed 103 barcode identifiers. Three barcode identifiers were shared by the remaining six taxonomic entities with two entities each. Of note, they could be easily distinguished by sequence dif-ferences in the ITS1 region. The results of species diversities in the barcode identifier region are given inTable 1and are open-access available to professional users in diagnostic microbiology labs
(www.blackyeast.org). This highly practicable tool for
identifica-tion ofHerpotrichiellaceaeis an important step toward rapid on-line identification of black yeasts and relatives, which always have been regarded as a diagnostic nightmare (3).
The ITS2 region in fungi is regarded as relatively stable as shown by Krüger & Gargas (12). Domain 2 of the ITS2 region turned out as a promising candidate for barcode identifiers because it provides highly conserved flanking sequences with a highly variable section in between, which varies markedly in both length and nucleotide composition even between closely related taxonomic entities (Table 1). The sequevars outlined in this study in part may have a tentative character, because stud-ies on specstud-ies delimitation are ongoing. It is recommended to take the barcode identifier into account with the description of new black yeast species, in analogy to dermatophyte taxonomy and molecular identification (8). Of note, such identification approaches are artificial but straightforward and hence highly practical. The availability of an online, continuously updated platform with validated barcodes will contribute to stability in taxonomy and diagnostics of black yeasts and relatives. A pre-liminary analysis of domain 2 of the ITS2 region ofAspergillus, Candida, Cladosporium, Fusarium, and Trichophyton species revealed that the interspecies variance is too low to be used as a reliable barcode identifier (data not shown). Here, identifica-tion is achievable with other domains of the ITS region (17), -tubulin, translation elongation factor 1-␣, and other genes. For this reason, the presented barcode identifier approach is restricted so far to members of the orderChaetothyriales. Main human pathogens are located in the familyHerpotrichiellaceae, but the recent reclassification ofConiosporium epidermidisto Knufia epidermidis(18), a genus belonging to the Chaetothyri-aceae, indicates that the proposed barcode identifier tool is also applicable to the familyChaetothyriaceae.
We here introduce a highly practicable tool for species assign-ment of black yeast isolates in a routine diagnostic laboratory to overcome problems associated by using the (nearly) entire ITS rDNA for identification. To achieve reliable species identification, only ITS2 region sequences or even shorter fragments comprising the barcode identifier segment are required. Thereby, the short length of the barcode, here introduced as the barcode identifier (27 to 50 bp), enables the usage of straightforward pyrosequenc-ing techniques (2,17). Furthermore, metagenomic pyrosequenc-ing approaches like tag-encoded FLX amplicon pyrosequencpyrosequenc-ing (TEFAP) (7), where the quality of identification benefits from short barcode identifiers, becomes feasible. Rapid and cost-effec-tive identification and analysis ofHerpotrichiellaceaepresent in biofilms can be achieved (unpublished results). Resulting short sequences can easily be matched with the barcode identifiers pro-vided inTable 1. A searchable curated online database will be implemented.
In conclusion, we present an easy-to-use, reliable, highly prac-ticable, and cost-effective approach for rapid molecular identifi-cation of clinically importantChaetothyriales(5).
ACKNOWLEDGMENT
G. Heinrichs was supported by a research grant from Rheinenergie AG.
REFERENCES
1.Badali H, et al.2011.Cladophialophora psammophila, a novel species of Chaetothyriales with a potential use in the bioremediation of volatile
ar-omatic hydrocarbons. Fungal Biol.115:1019 –1029.
2.Borman AM, et al.2010. Rapid molecular identification of pathogenic yeasts by pyrosequencing analysis of 35 nucleotides of internal transcribed
spacer 2. J. Clin. Microbiol.48:3648 –3653.
3.Caligiorne RB, Licinio P, Dupont J, de Hoog GS.2005. Internal tran-scribed spacer rRNA gene-based phylogenetic reconstruction using algo-rithms with local and global sequence alignment for black yeasts and their
relatives. J. Clin. Microbiol.43:2816 –2823.
4.Clarke LA, Rebelo CS, Gonçalves J, Boavida MG, Jordan P.2001. PCR amplification introduces errors into mononucleotide and dinucleotide
repeat sequences. Mol. Pathol.54:351–353.
5.De Hoog GS, Guarro J, Gené Figueras JMJ.2011. Atlas of clinical fungi, electronic version 3.1. Centraalbureau voor Schimmelcultures/ Universitat Rovira i Virgili, Utrecht, Netherlands.
6.De Hoog GS, et al.2011. WaterborneExophialaspecies causing disease in
cold-blooded animals. Persoonia27:46 –72.
7.Dowd SE, et al.2008. Evaluation of the bacterial diversity in the feces of cattle using 16S rDNA bacterial tag-encoded FLX amplicon
pyrosequenc-ing (bTEFAP). BMC Microbiol.8:125–132.
8.Gräser Y, Scott J, Summerbell R. 2008. The new species concept in
dermatophytes—a polyphasic approach. Mycopathologia166:239 –256.
9.Hall TA.1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp.
41:95–98.
10. Iwen PC, Hinrichs SH, Ruppy ME.2002. Utilization of the internal transcribed spacer regions as molecular targets to detect and identify
hu-man fungal pathogens. Med. Mycol.40:87–109.
11. Keller A, et al.2009. 5.8S–28S rRNA interaction and HMM-based ITS2
annotation. Gene430:50 –57.
12. Krüger D, Gargas A. 2004. The basidiomycete genus Polyporus—an emendation based on phylogeny and putative secondary structure of
ri-bosomal RNA molecules. Feddes Repert.115:530 –546.
13. Krüger D, Gargas A.2008. Secondary structure of ITS2 rRNA provides taxonomic characters for systematic studies—a case in Lycoperdaceae
(Basidiomycota). Mycol. Res.112:316 –330.
14. Lian X, de Hoog GS.2010. Indoor wet cells harbour melanized agents of
cutaneous infection. Med. Mycol.48:622– 628.
15. Matos T, Haase G, Gerrits van den Ende AHG, de Hoog GS.2003. Molecular diversity of oligotrophic and neurotropic members of the black
yeast genusExophiala, with accent onE. dermatitidis. Ant. Leeuw.83:293–
303.
on May 16, 2020 by guest
http://jcm.asm.org/
16. Nilsson RH, et al.2006. Taxonomic reliability of DNA sequences in
public sequence databases: a fungal perspective. PLoS One1:e59 – e62.
doi:10.1371/journal.pone.0000059.
17. Pannanusorn S, Elings MA, Romling U, Fernandez V.2012. Pyrose-quencing of a hypervariable region in the internal transcribed spacer 2 to
identify clinical yeast isolates. Mycoses55:172–180.
18. Tsuneda A, Hambleton S, Currah RS.2011. The anamorph genusKnufia
and its phylogenetically allied species inConiosporium,Sarcinomyces, and
Phaeococcomyces. Can. J. Bot.89:523–536.
19. Zalar P, Novak M, de Hoog GS, Gunde-Cimerman N.2011. Dishwash-ers—a man-made ecological niche accommodating human opportunistic
fungal pathogens. Fungal Biol.115:997–1007.
20. Zeng JS, de Hoog GS.2008.Exophiala spiniferaand its allies: diagnostics
from morphology to DNA barcoding. Med. Mycol.46:193–208.