G
HEORGHEG
LUHOVSCHI1, C
RISTINAG
LUHOVSCHI1, F
LAVIUB
OB1, S
ILVIAV
ELCIOV1,
V
IRGINIAT
RANDAFIRESCU1, L
IGIAP
ETRICA1, G
HEORGHEB
OZDOG1, D
ANIELC
IOCA2Immune Processes at the Level of the Juxtaglomerular
Apparatus and Their Relationship
with Hypertension and Immune−Mediated
Nephropathies – State of the Art
Procesy immunologiczne w obrębie aparatu przykłębuszkowego
i ich związek z nadciśnieniem tętniczym i chorobami nerek
o podłożu immunologicznym
1 Nephrology Department, Victor Babes University of Medicine and Pharmacy, Timisoara, Romania 2Center for Immunophysiology and Biotechnologies, Victor Babes University of Medicine and Pharmacy,
Timisoara, Romania, and the Austrian Academy of Sciences, Immunology Division, Innsbruck, Austria
Adv Clin Exp Med 2009, 18, 6, 623–639 ISSN 1230–025X
REVIEWS
© Copyright by Wroclaw Medical University
Abstract
The paper presents aspects of immune processes at the level of the juxtaglomerular apparatus (JGA) and their re− lationship with hypertensive pathology and immune−mediated glomerular nephropathies. In various immune−me− diated diseases of the JGA, such as, among others, systemic lupus erythematosus (SLE), IgA nephropathy, and dia− betes mellitus, markers of immune processes are present. During immune processes that affect the extraglomeru− lar mesangium by means of immune deposits or immune cells, the transmission of information from the level of the macula densa to the afferent arteriole can be perturbed, with alteration of the tubulo−glomerular feedback and with probable perturbation of the mechanisms of blood pressure regulation. Blood vessels, predominantly the af− ferent arteriole, participate in immune processes such as vasculitis, acute antibody−mediated graft rejection, and ne− phrosclerosis, which are all associated with hypertension. The occurrence of JGA vessel lesions during immune− mediated diseases which themselves affect the JGA could amplify the severity of hypertension in these diseases. Because in some diseases, such as diabetes mellitus, diminution of the lumen of JGA vessels has been found, the suspicion arises that lesions of the respective afferent arterioles may cause Goldblatt−like phenomena, and because the lesion is diffuse, we are faced with a sum of Goldblatt phenomena, which may cause a lesion similar to that en− countered in renal artery stenosis. The extraglomerular mesangium of the JGA communicates with the intraglome− rular mesangium. Some studies plead for the fact that the cells of the extraglomerular mesangium are involved in regenerative processes of the intraglomerular mesangium, which is damaged in glomerulonephritis. CD34 markers encountered at the level of stem cells have been evidenced at the level of the extraglomerular mesangium. Intrare− nal RAS could play an important role in the arterial hypertension pathogenesis of AHT (arterial hypertension) and in glomerular nephropathy. It can be an alternative to or a completion of the role of the JGA (Adv Clin Exp Med 2009, 18, 6, 623–639).
Key words:JGA, immunopathology, vascular disease, glomerulopathy.
Streszczenie
The juxtaglomerular apparatus (JGA) is a spe− cialized contact region between the glomerulus and the thick cortical ascending limb. It consists of afferent and efferent arterioles, extraglomerular mesangium, and the macula densa. The JGA plays an active role in the maintenance of ion homeosta− sis and the control of blood pressure [1]. There− fore, the JGA has been most often studied from the perspective of its involvement in arterial hyperten− sion (AHT).
The kidney is an organ that is frequently in− volved in immune pathological processes. These processes can affect the glomeruli, the intersti− tium, and the vessels and cause immune−mediated glomerulonephritis, immune−mediated tubulo−in− terstitial nephritis, and immune−mediated vasculi− tis, respectively. The juxtaglomerular region is in close proximity to the glomerulus and has a close connection to the nephron. The extraglomerular mesangium communicates with the intraglomeru− lar mesangium; the afferent arteriole and the effe− rent arteriole, together with the glomerular capilla− ries and the interstitial vessels, form a vascular net− work, and the distal tubule participates, by means of the macula densa, in the formation of the juxta− glomerular apparatus.
Although the immune−mediated diseases that involve the components of the nephron are well studied, less is known about what is happening at the level of the JGA during immune processes. Morphopathological results usually do not men− tion pathological processes taking place at the JGA, although this region has a pivotal role in blood pressure (BP) control. At the same time, ma− ny of the immune−mediated renal diseases are ac− companied by arterial hypertension. An involve− ment of the JGA during pathological processes of the nephron seems likely. Moreover, the pathoge− netic mechanisms of arterial hypertension in im− mune−mediated glomerular nephropathies and va− sculitis have not yet been completely explained, and the possible involvement of the JGA needs to be studied.
This paper presents and analyzes the main da− ta in the literature concerning JGA involvement during the primary immunological diseases of the kidney or those involving the kidney secondarily. Concurrently, the paper analyzes the possible con− sequences of JGA involvement during immune− mediated diseases upon BP regulation and AHT production, respectively. To neglect the involve− ment of this important area of the nephron in im− mune processes is, in our opinion, unjustified. Al− so, to postpone approaching this problem until all pathological mechanisms have been elucidated would be unproductive.
Data Regarding
the Structure and Function
of the JGA and Implications
in Pathology
The JGA consists of afferent and efferent arte− rioles, extraglomerular mesangium, and the macu− la densa. The macula densa is a specialized epithe− lial cell plaque in the wall of the thick ascending limb located at the site where the limb joins the extraglomerular mesangium of the glomerulus [2]. The extraglomerular mesangium is a solid block of cells and matrix filling the space between the ma− cula densa and both arterioles, extending into the entrance funnel, and occluding the glomerular tuft [3]. The extraglomerular mesangium is composed of ramified, heavily branched cells surrounded by matrix and coupled to each other and to vascular cells by gap junctions. It represents a physiologi− cal liaison between macula densa cells and arterio− les, smooth muscle cells, and the arteriolar granu− lar myoepitheloid cells. in particular, the extraglo− merular mesangium interconnects all structures of the glomerular entrance [3].
According to Rosivall, one of the unique mor− phological characteristics of the JGA is the absence of capillaries and lymph vessels in the extraglome−
przeszczepu mediowane przez przeciwciała, miażdżycopochodne stwardnienie nerki, z których wszystkie są zwią− zane z nadciśnieniem. Występowanie uszkodzeń naczyń w JGA w czasie chorób o podłożu odpornościowym, które same mogą naruszać JGA, mogłoby zwiększyć nasilenie nadciśnienia w tych chorobach. Ponieważ w niektórych chorobach, takich jak cukrzyca, wykryto zmniejszenie światła naczyń JGA, podejrzewa się, że zmiany w odpowie− dnich doprowadzających tętniczkach mogą wywołać zmiany podobne do zjawiska Goldblatta, a ponieważ zmiany są rozlane, występuje suma zjawisk Goldblatta, które mogą wywoływać uszkodzenia podobne do spotykanych w zwężeniu tętnicy nerkowej. Mezangium zewnętrzne JGA komunikuje się z mezangium wewnętrznym. Niektóre badania powołują się na to, że komórki mezangium zewnętrznego są zaangażowane w procesy regeneracyjne me− zangium wewnętrznego, które jest uszkodzone w kłębuszkowym zapaleniu nerek. Markery CD34 występujące w komórkach macierzystych zostały wykryte także w mezangium zewnętrznym. Zwężenie tętnicy nerkowej może odgrywać istotną rolę w patogenezie nadciśnienia tętniczego i kłębuszkowego zapalenia nerek. Może być alterna− tywą lub uzupełnieniem roli JGA (Adv Clin Exp Med 2009, 18, 6, 623–639).
rular mesangium, i.e. in the area between the affe− rent and efferent arterioles. This area is composed of densely packed mesangial cells amidst an inter− stitial fluid with basal laminal material [4]. The va− scular component of the JGA includes the terminal portion of the afferent arteriole and the proximal po− rtion of the efferent arteriole [5]. In the walls of the afferent arterioles are granular cells, i.e. modified smooth muscle cells present in groups (of four to fi− ve cells) in the terminal portion of the afferent arte− riole and, rarely, also in the efferent arteriole. The number of granular cells varies greatly because fila− ment−rich smooth muscle cells can undergo meta− plasia to granular cells and produce renin that is stocked in the granules [6]. The synthesis and rele− ase of renin from juxtaglomerular cells are control− led by humoral factors, changes in arteriolar pressu− re, the activity of sympathetic nerves, and signaling by the macula densa. Renin synthesis is also stimu− lated by prostaglandins and is inhibited through a negative feedback loop by angiotensin II, high blood pressure, salt, and volume overload [7]. The granular cells are in contact with the extraglomeru− lar mesangium, adjacent smooth muscle cells, and arteriolar endothelium and are coupled to these structures by gap junctions.
Other cells of the JGA are also coupled to each other and to vascular cells by gap junctions [5]. Gap junctions are made up of connexins. Connexin proteins play a role in cell−to−cell communication via intercellular gap junction channels or transien− tly open hemichannels [8]. Renin−producing cells predominantly express connexin 40 and, to a lesser extent, connexin 37. Connexin 40 appears to be cri− tically involved in the calcium−dependent control of renin secretion and is also probably involved in renin synthesis. Recently, the expression of conne− xin 40 in the JGA has been demonstrated in mice, where it is involved in the regulation of renin secre− tion and blood pressure. Hanner et al. reported the important role played by connexin 45 in the propa− gation of JGA vascular signals and in the regulation of renin release and blood pressure [8]. Connexin 45 could regulate calcium wave propagation via di− rect gap junction coupling [8].
The Functions of the JGA
The JGA exhibits a complex array of functions which regulate the process of filtrate formation and renal hemodynamics [9]. Its main functions are: 1) control and adaptation of the glomerular fil− tration rate (GFR) to early distal tubular NaCl (tu− buloglomerular feedback) and 2) the synthesis and release of renin. Both of these functions relate to changes in luminal NaCl concentration and NaCl transport by the macula densa. The JGA is a spe−
cialized contact region between the glomerulus and the cortical thick ascending limb that plays an active role in the maintenance of ion homeostasis and the control of blood pressure. Macula densa cells are renal sensor elements that detect changes in distal tubular fluid composition and transmit si− gnals to the glomerular vascular elements. Macula densa cells detect changes in the luminal sodium chloride concentration through a complex series of ion transport−related intracellular events. NaCl en− ters via a Na:K:2Cl cotransporter and Cl exits th− rough a basolateral channel [10]. The JGA is an important regulatory site of renal salt and water conservation and systemic blood pressure homeo− stasis [4]. Nitric oxide released from the macula densa can modulate tubuloglomerular feedback in response to increased tubular NaCl by counterac− ting vasoconstriction, by reducing macula densa transport, or by interacting with local prostaglan− din synthesis. In addition, nitric oxide may also adjust renin production. The glomerular hyperfil− tration seen in diabetic nephropathy is related to the JGA. Thus, increased expression of endothelial cell nitric oxide synthase (eNOS) in afferent and glomerular endothelial cells is involved [11].
The role of the JGA in pathology is complex: • microvascular autoregulatory responsive− ness to changes in perfusion pressure: ATII has a role in the regulation of renal hemodynamics, tu− bular function regulation, and tubuloglomerular feedback regulation,
• the activation of the renin−angiotensin−al− dosterone pathway through increased production of renin has been found in renal artery stenosis,
• the activation of this system is also present in other forms of arterial hypertension,
• the activation of the renin−angiotensin sy− stem through increased renin production is present in other diseases, such as nephritic syndrome, he− art failure, and hepatic cirrhosis,
• increased production of renin has been ob− served in patients with JGA tumors,
• the therapeutic blocking of renin produc− tion can be achieved by using some drugs. Recen− tly, the blocking of renin synthesis through Aliski− ren has been reported. Aliskiren accumulates in re− nin secretory granules and binds plasma pro−renin, thus allowing long− lasting blocking of the renin− angiotensin system [12],
• connexins are involved in the propagation of JGA vascular signals and blood pressure regula− tion [8],
• involvement of the JGA in human immune pathology.
Data from the literature indicate the involve− ment of some elements in diseases with an immu− ne component, such as SLE, IgA nephropathy, and diabetes mellitus. As mentioned above, damage to the arterioles of the JGA in diseases associated with immune processes suggests a relationship be− tween them. Arterioles, predominantly the afferent ones, are involved in immune processes such as vasculitis, acute antibody−mediated graft rejection, and nephrosclerosis [14].
The complexities of AHT pathology are cur− rently being studied, but with very little reference to JGA involvement during the immune pathologi− cal processes that affect it. It is surprising how few studies in the literature mention the involvement of the JGA and, specifically, the involvement of its vessels during immune−mediated vascular or renal diseases. The aim of the present paper is to present an overview of the current immunopathological data at the JGA level and its relationship to immu− ne glomerular and vascular pathology and to AHT. This paper mainly refers to three aspects of the in− volvement of the JGA to disease: 1) immune pro− cesses involving the extraglomerular mesangium, 2) the role of the extraglomerular mesangium in regenerative processes of the intraglomerular me− sangium, and 3) the relationship between JGA ves− sels, immunology and AHT. To fulfill these objec− tives, this paper presents information on some diseases in which the presence of immune proces− ses has been reported at the level of the JGA. The association of these diseases with AHT has been mentioned. Not only those pathological conditions in which the involvement of the JGA has been cle− arly demonstrated will be dealt with, but also situ− ations in which its participation in immune proces− ses has been signaled. Some of the data presented will need to be clarified in the future. Because of the paucity of knowledge of the immune patholo− gy of the JGA, well−established data have been emphasized, while controversial data have been avoided. In the last part of this article some future directions derived from the data presented in this paper are discussed with the goal of attracting some attention to this problem, which should lead to new studies in this field of pathology in which so little is known.
Immune Processes Involv−
ing the Extraglomerular
Mesangium
The Involvement of Humoral
Immunity at the Level of the
Extraglomerular Mesangium
Immune−mediated glomerular diseases are fre− quently accompanied by immune deposits at the glomerular level. The deposition of immune com− plexes and deposits in the JGA has been less extensively studied. Due to the ability of mesan− gial cells, which belong to the microphage system, to capture immune complexes, the question is rai− sed of whether extraglomerular mesangial cells play a similar role.
In systemic lupus erythematosus (SLE), immu− ne deposits in the JGA were reported by Silva et al., who showed discrete electron−dense “immune− type” deposits in the JGA in patients with SLE [15]. In addition, Hvala et al. reported an electron microscopic fingerprint and other organized depo− sits in renal biopsies obtained from SLE patients. Besides mesangial and interstitial deposition and deposition alongside tubules and the peritubular capillary basement membrane, fingerprint deposits have also been demonstrated on the arteriolar wall and in the JGA in biopsies [16]. Mechanisms of cellular and humoral immunity have an important role in SLE. Many authors have reported immune abnormalities in lupus nephritis [17]. Mesangial immune deposits as well as deposits at the level of the glomerular basement membrane underlie the development of mesangial proliferative nephritis and membranous nephropathy. The concomitant pre− sence of immune deposits at the level of the extraglo− merular and intraglomerular mesangium shows a si− multaneous involvement in immune injury.
In a study of the juxtaglomerular apparatus in IgA nephropathy, Hara et al. reported that IgA de− posits are rare in the juxtaglomerular apparatus de− spite a large amount of IgA deposits in the mesan− gium. Concomitant C3 deposits have been found in the mesangium, in the lacis region, and in the af− ferent and efferent arteriolar walls [18]. In IgA ne− phropathy, which is a mesangial proliferative ne− phritis, there are circulating immune complexes as well as deposits at the mesangial level. The find− ings of Hara et al. show concomitant involvement of the glomerulus and the JGA in immune proces− ses in IgA nephropathy, similar to that found in lu− pus nephritis [18].
from patients with glomerulonephritis with mesan− gial IgM deposits. Biopsies also showed mesan− gial IgA deposition, but IgA was not seen in the macula densa. They suggest that mesangial IgM deposits may reflect non−specific macromolecular transport rather than an immune reactant. The pre− sence of immune markers in the JGA is not syno− nymous with the activation of immune mechani− sms at this level, and a trapping of immune com− plexes at the cellular level of the juxtaglomerular apparatus cannot be excluded [19].
Although it has not yet been demonstrated in humans, it cannot be excluded that the presence of immune deposits at the level of the JGA affects this zone and alters the functionality of the JGA.
The Involvement of Cellular
Immunity at the Level of the
Juxtaglomerular Apparatus
in Diabetes Mellitus
The presence of bone marrow−derived immune cells at the level of the JGA constitutes an impor− tant indication of their involvement in immune−me− diated renal diseases. Cellular immune−mediated mechanisms are present in many immune−mediated diseases that affect the kidney; however, concomi− tant involvement of the JGA has only been mentio− ned sporadically. Diabetes mellitus (DM), compli− cated by diabetic nephropathy, has been studied with respect to the participation of cellular immu− ne−mediated mechanisms (T cells) at the level of the JGA.
According to Moriya et al., type 1 diabetes mellitus is an autoimmune disorder associated with T cell−mediated injury to multiple endocrine tissues. In diabetic nephropathy secondary to type 1 diabetes mellitus, T−cell infiltration of the JGA has been found. This finding raises the hypothesis that immunological injury of the JGA in type 1 diabetes mellitus could alter the clinical consequ− ences of diabetic nephropathy. The presence of T cells in the JGA could be associated with chan− ges in local renin−angiotensin activity [20], which is suggestive of autoimmune processes, according to Poulsen [21]. The presence of an immune T−cell infiltrate in diabetic nephropathy at the level of the JGA mesangium could also perturb the functiona− lity of the JGA and the transmission of information through the extraglomerular mesangium zone from the macula densa to the afferent arteriole in a manner similar to the deposition of immune complexes.
Autoimmune Phenomena
and the Juxtaglomerular
Apparatus
Autoimmune phenomena are frequently invo− lved in pathology. The presence of structures with antigenicity similar to that seen in other zones of the organism (endothelial cells, adhesion molecu− les) at the level of the JGA raises the question of whether autoimmune mechanisms that affect other zones in the organism may affect the JGA as well. Autoimmune phenomena at the level of vessels in different diseases, such as vasculitis, could cause arteriolar lesions in the JGA, with alteration of pressor mechanisms at the glomerular level. Auto− antibodies against intercellular adhesion mole− cule−1 (ICAM−1) and E−selectin are present in au− toimmune vasculitis [22, 23]. There is just one re− port which implicates autoimmune mechanisms at the level of the JGA. Mavrikakis et al. found a po− sitive correlation between rheumatoid factor le− vels, microhematuria, and plasma renin activity in patients with rheumatoid arthritis [24].
Although current knowledge of immune pa− thophysiology at the level of the JGA is rather scarce in comparison with that of its role in the glomerulus or other segments of the nephron, its presentation is important. As long as mesangial cells are included in the macrophage/microphage system and the extraglomerular mesangial cells communicate with cells of the glomerular mesan− gium where they can migrate, it is difficult to se− parate them from the immune system.
The Consequences of the
Presence of Immune Processes
for the Functionality of the
Extraglomerular Mesangium
The extraglomerular mesangium has a strate− gic disposition between the macula densa and the afferent arteriole. At the same time, cells are loca− ted at the vascular pole of the glomerulus, while
Fig. 1.The juxtaglomerular apparatus and immunopathological processes
the cells of the extraglomerular mesangium form a syncytium and are connected with glomerular mesangial cells via gap junctions [25]. Gap junc− tion−mediated intercellular communication in the JGA provides a pathway for signal transduction and coordination of multicellular functions [26]. Blood vessels are missing in the extraglomerular mesangium. Messenger systems between the ma− cula densa and the afferent arteriole are involved in the transfer of signals from the macula densa to mesangial cells and from mesangial cells to the af− ferent arteriole. There are several candidate mole− cules that may function in this capacity [4]. Great attention is given to phospholemman, a single− span membrane protein homologous to the Na K− ATPase gamma subunit. Its location in the juxta− glomerular apparatus suggests that it is involved in
tubuloglomerular feedback [27]. Rosivall et al. have shown that a significant and dynamic flow exists in the JGA which may help filter the renin released into the renal interstitium (endocrine function). It may also modulate tubuloglomerular feedback and transmit signals to the JGA (hemo− dynamic function) [14].
The presence of some pathological processes at this level could perturb this extremely fine and sensitive communication system that lies at the ba− sis of tubuloglomerular feedback. The exact me− chanism through which the presence of immune cells and immune complexes in the JGA perturbs the tubuloglomerular feedback is not well known. Mentioned above was the transmission of informa− tion from the macula densa to the extraglomerular mesangium in terms of tubuloglomerular feedback
Table 1. The extraglomerular mesangium and mesangial regenerative processes
Tablica 1. Mezangium zewnętrzne oraz regeneracyjne procesy mezangialne
Observations Pathogenesis Study type Authors
(Uwagi) (Implikacje patogenetyczne) (Rodzaj (Autorzy)
studium)
Migration of cells migration of cells from hilar area and extraglomerular anti−Thy 1.1 Hugo et al. 1997 (Migracja komórek) mesangium into the juxtaglomerular apparatus model
migration of mesangial cells is dependent on PDGF−BB and anti−Thy 1.1 Haisley et al.
partially on bFGF model 1999
CD34 CD34−positive cells in the extraglomerular mesangium and human Gluhovschi et al.
glomerular mesangium biopsies 2008
CD34 in mesangial cells corresponds to alpha smooth muscle human Naruse et al.
actin (ASMA) expression biopsies 1999
Origin of 11–12% of glomerular cells were derived from transplanted Ito et al. 2003 mesangial cells bone marrow
(Powstanie komórek circulating stem cells engraft into the kidney and differentiate in situhybrid− Poulson et al.
mezangialnych) into renal parenchymal cells ization 2003
bone marrow derived cells differentiate into glomerular in vitrostudies Imasawa et al.
mesangial cells (mice and rats) 2001, Iwatani et
al. 1999 hematopoietic stem cells differentiate into glomerular experimental Masuya et al.
mesangial cells model 2003
transplanted mesenchymal stem cells accelerate glomerular experimental Kunter et al.
healing model 2006
progenitor cell migration from the juxtaglomerular apparatus ischemia− Baud et al. 2005
and also bone marrow reperfusion
model
Transmission of blocking the tubulo−glomerular feedback by administering in vitrostudy Ren et al. 2002 information from anti−thy1.1 and complement to the JGA (rabbits)
macula densa to Significant and dynamic flow exists in the JGA experimental Rosivall et al.
afferent and efferent model 2006
arteriole
through channels that are formed between extra− glomerular mesangial cells [4]. The channels formed enable gap junction−mediated intercellular communication in the JGA, which provides a pathway for signal transduction. Disruption of cell−to−cell communication in the JGA results in altered preglomerular vascular tone and renin se− cretion [26]. The exact mechanism through which immune deposits and cells that infiltrate the JGA affects the tubuloglomerular feedback and blood pressure regulation is not yet clarified. However, their presence in the JGA is very likely involved in the dysregulation of this extremely fine−tuned me− chanism.
To demonstrate this phenomenon, Ren et al. performed an experimental study which showed the possible involvement of lesions secondary to im− mune processes in JGA function. They administe− red an anti−Thy 1−1 antibody and complement to in
vitroperfused JGAs isolated from male New Zea−
land white rabbits. The tubuloglomerular feedback was completely eliminated in this system. They suggested that both intra− and extraglomerular me− sangia and gap junctions were necessary for tubulo− glomerular feedback [28]. The alteration of these structures, secondary to immune−induced lesions, indicates that the immune processes in the JGA may have significant pathological consequences.
In humans, the presence of immune deposits in immune glomerular nephropathies (lupus ne− phritis, IgA nephropathy) or the cellular immune infiltrates (e.g. diabetic nephropathy) could also act as obstacles to the flux between the macula densa and the afferent arterioles and have nega− tive effects on the functionality of the extraglo− merular mesangium and the transmission of the informational message between afferent arterio− les and the macula densa. They could also inter− fere with the regulation of blood pressure. This phenomenon could be compared with traffic bar− riers in crowded streets of a large city, in which an apparently well−organized circulation is per− turbed. Although the involvement of immune processes in the extraglomerular mesangium in producing arterial hypertension in human immu− ne−mediated glomerular diseases has not yet been demonstrated, the possibility could be interesting for future studies.
The Extraglomerular
Mesangium
in the Regenerative
Processes of the Glomerular
Mesangium
The immune pathology of the JGA must be considered in its complexity, with reference to all of its components and the possible relationships between them. The involvement of the extraglo− merular mesangium in regenerative processes of the glomerular mesangium is presented here by both experimental studies and clinical data. Stem cells are still insufficiently delimited in the neph− ron and they intervene in regenerative processes. Their interrelation with glomerular and extraglo− merular mesangia must be discussed.
Experimental Studies
The extraglomerular mesangium communica− tes with the glomerular mesangium at the level of the hilus. Hugo et al. revealed the extraglomerular origin of mesangial cells and underlined the im− portance of this juxtaglomerular origin after their injury. They also demonstrated the role played by the juxtaglomerular apparatus in the repair of glo− merular lesions. In experimental GNs mediated through anti−Thy 1.1 antibodies in rats, the same authors showed that injection of an anti−Thy 1.1 antibody that binds the Thy 1.1 antigen on rat mesangial cells eliminated the mesangial cell po− pulation. Thy−1−positive cells in the JGA were se− questered from the circulation and survived [13]. Serial biopsies have shown the progressive migra− tion and proliferation of these cells until the entire glomerulus is repopulated. According to Hugo et al., this migration of mesangial cells from the extraglomerular mesangium argues in favor of a role for the juxtaglomerular apparatus in the maintenance of the mesangial cell population, a normal mesangial cell pool after injury [13]. The same authors introduced the term “reserve mesan− gial cells” that may characterize the extraglomerular and hilar Thy 1.1−postive cells [14]. Haseley et al. suggest that these cells may also play an important role in the repair of mesangial lesions. It is believed that the migration of the mesangial cells is depen− dent on PDGF−BB and partially on bFGF [29].
Clinical Studies in Humans
sangium as well as in the glomerular mesangium [30]. CD34 is also present on stem/progenitor cells and on mesangial cells that have undergone trans− differentiation [31]. Naruse reported that CD34 expression can serve as a marker of transfor− med/activated mesangial cells in glomerular dis− ease. In normal glomeruli, mesangial cells were negative for CD34, but in glomerulonephritis, CD34 was expressed in mesangial cells, which were identified by alpha−smooth muscle actin (ASMA) expression [32]. The presence of CD34− positive cells in the extraglomerular mesangium may indicate that the migration of cells from the extraglomerular mesangium can be associated with the presence of stem/regenerative cells. It may also indicate that extraglomerular mesangial cells undergo a process of trans−differentiation that is related to their migration to the glomerular me− sangium. In addition, the presence of CD34−posi− tive cells in the glomerular mesangium has a simi− lar significance. CD34 expression at this site may indicate trans−differentiation of mesangial cells or the presence of stem/regenerative cells.
Stem Cells and Regenerative
Processes
Bone marrow is considered by Ito et al. to be a reservoir of repopulating mesangial cells during glomerular remodeling. In restored glomeruli, 11 to 12% of glomerular cells were derived from transplanted bone marrow [33]. One of the most debated problems is that of stem cells at the level of the kidney, and specifically in the nephron. The renal papilla and, recently, Bowman’s capsule have been found to be stem cells niches [34, 35]. It is known that a number of glomerular mesangial cells originate in the bone marrow. Moreover, some circulating stem cells can reach the kidney and differentiate into resident cells. The relation− ship between stem cells and the extraglomerular mesangium is not yet known, but because of its involvement in glomerular regenerative processes, it cannot be eluded.
Experimental studies:By in situhybridization,
Poulsom et al. demonstrated that circulating stem cells frequently engraft in the kidney and differen− tiate into renal parenchymal cells [36]. Bone mar− row−derived cells have the potential to differentia− te into glomerular mesangial cells in mice and rats [37–39]. Furthermore, Masuya et al. demonstrated that a hematopoietic stem cell is capable of diffe− rentiating into a glomerular mesangial cell type [40]. Transplanted mesenchymal stem cells acce− lerate glomerular healing in experimental glome− rulonephritis in rats [41]. These studies underscore the possibility that certain types of stem cells, such
as mesenchymal stem cells, reach the nephron by a direct vascular pathway, without passing through the extraglomerular mesangium.
Clinical studies: El Nahas suggested that phe−
notypic changes characteristic of the immune re− sponse include the migration of hematopoietic progenitor cells into injured glomeruli, which is suggestive of the involvement of stem cells. Ac− cording to El Nahas, these cells appear to be in− volved in the normal turnover of mesangial cells and in the response of the mesangium to injury [42]. Repopulating mesangial cells seem to enter the glomerulus through the juxtaglomerular zone [14, 43]. PDGF−BB is an important factor for the conversion of bone marrow−derived stem cells in− to mesangial cells [44]. Baud et al. propose that in glomerulonephritis with mesangiolysis, mesangial regeneration involves both progenitor cell migra− tion from the JGA and bone marrow−derived cells [45]. As the activity and presence of stem cells in the kidney are not yet completely understood, stu− dies concerning the JGA and its role in regenera− ting lesions at the level of the nephron are neces− sary. Also, the trans−differentiation processes at the level of the glomerulus also requires further elucidation.
The JGA, through the extraglomerular mesan− gium, seems to play an important role in regenera− tive processes at the level of the glomerulus that take place following immune−mediated lesions in the glomerulus. The importance of the participa− tion of the JGA in immune processes in the glome− rulus has not yet been defined. Stem cells also have a probable role in this process. The finding of stem−cell markers in the extraglomerular mesan− gium amplifies the significance of the involvement of this region in the immunopathology of glomeru− lar nephropathies.
Vascular Immune Pathology
and the Juxtaglomerular
Apparatus
main hypotensive drugs, such as ACEIs and ARBs, is reduced or avoided in cases of advanced chronic kidney disease, such as in the case of ad− vanced renal insufficiency.
Immune−mediated vascular pathology is of great relevance and is being approached in many studies. Immune processes are demonstrated to be involved in atherosclerosis and hypertension. Be− sides the lesions in arterioles of the entire vascular bed seen in these diseases, the JGA arterioles are selectively affected during some pathological pro− cesses, such as in humoral acute rejection and du− ring treatment with calcineurin inhibitors. The le− sions in arterioles in vasculitis and diabetes melli− tus are important and have been reported to affect the JGA arterioles.
Immune−Mediated Arteriolar
Lesions That Primarily Affect
The Kidney
In humoral acute rejection, important patholo−
gical changes take place that affect arterioles, especially the afferent arteriole [46]. Antibody− mediated rejection is pathologically characterized by capillary endothelial swelling, arteriolar fibri− noid necrosis, and fibrin thrombi in glomerular ca− pillaries [47]. The presence of arteriolar fibrinoid necrosis is a sensitive factor in the diagnosis of acute humoral rejection [48, 49]. During acute graft rejection, mechanisms of humoral immunity can involve the arterioles of the JGA, contributing to the latter’s complex pathogenesis.
Vascular Lesions Caused by
Immunomodulatory Drugs that
Affect the Kidney
Treatment with calcineurin inhibitors. The
most characteristic lesion of chronic calcineurin inhibitor nephrotoxicity is arteriolar hyaline thic− kening (if nodular) [50]. Obliterative vasculopathy of the afferent arteriole and tubulo−interstitial fi− brosis in grafts have been interpreted to be a sign of cyclosporin nephrotoxicity and may explain at least the severity of hypertension. Vascular lesions at the level of the JGA caused by immunomodula− tory drugs can be complex; they are due not only to the rejection, but also to the cyclosporin therapy and the possible additive effects of the two [51]. In both situations, arteriolar lesions can lead to an al− teration of the irrigation at the level of the afferent arteriole and its consequences and to the activation of pressor mechanisms at the glomerular level.
Vascular Lesions with Immune
Involvement that Affect
the Circulatory System
of the Whole Body,
Including the Kidney
Arterial Hypertension
Renal vascular damage which is mainly reflec− ted in the vessels of the JGA and especially the af− ferent arteriole is found in arterial hypertension. In benign nephrosclerosis, the lesion consists of affe− rent arteriole hyalinization that is frequently seg− mental [52]. There is a narrowing of the lumina of arterioles and small arteries, caused by thickening
Table 2. Cellular and humoral immune mechanisms at the level of the extraglomerular mesangium
Tabela 2.Komórkowe i humoralne mechanizmy immunologiczne na poziomie mezangium zewnętrznego
Disease Observed lesion Study type Authors
(Rodzaj choroby) (Zaobserwowane porażenie) (Rodzaj studium) (Autorzy) SLE electron−dense ”immune−type” deposits in the JGA human biopsies Silva et al. 1986
fingerprint deposits in the JGA human biopsies Hvala et al. 2000 IgA nephropathy rare IgA deposits in the JGA human biopsies Hara et al. 1988 (Nefropatia IgA)
GN with mesangial IgM deposits in the macula densa human biopsies Thompson
IgM deposits and Evans 2001
(Kłębkowe zapalenie nerek ze złogami IgM)
and hyalinization of the wall that is secondary to the hyaline vascular narrowing [53]. Lesions in benign nephrosclerosis can affect both afferent and efferent arterioles. They are also associated with atherosclerotic processes in other nephropathies, such as IgA nephropathy. According to Razga et al., efferent arteriolar lesions can result from an− giotensin II (AII)−mediated auto−regulatory effe− rent vasoconstriction that is exerted to maintain glomerular filtration pressure [54].
Malignant nephrosclerosis is characterized by fibrinoid necrosis of the afferent arteriole, which frequently extends into the glomerulus. Thus Di− kow and Ritz considered that the hallmark of ma− lignant hypertension is fibrinoid necrosis of the af− ferent arterioles [55]. The proliferation of the me− dia and intima causes a narrowing of the arteriolar lumen, with hemodynamic consequences [56]. The diffuse narrowing of the arteriolar lumen pro− duces glomerular ischemia, which affects pressor mechanisms with activation of the renin−angioten− sin aldosterone system. Malignant hypertension is usually associated with remarkably high levels of renin, angiotensin, and aldosterone [53].
Phenomena similar to those described by Gold− blatt in renal artery stenosis also take place at the level of every affected nephron, and through their summation a similar effect can be obtained. The se− condary involvement of the vascularization of the JGA can amplify arterial hypertension produced by vascular disease, resulting in a vicious circle.
Immune mechanisms are involved in the pa− thogenesis of vascular injury during the course of arterial hypertension, namely in benign and mali− gnant nephrosclerosis. Immune processes that come together with benign and malignant nephro− sclerosis by affecting the JGA arterioles could have a complementary role in their production. Until now, observations have been limited to some experimental studies. It could be useful to study these processes in humans as well.
Studies performed on animals have also reveal− ed changes in the afferent arteriole. Skov and Mu− lvany showed that renal afferent arterioles are structurally narrowed in young and adult sponta− neously hypertensive rats [57]. In genetically hy− pertensive rats, Ledingham and Laverty demon− strated that losartan and enalapril reduce blood pressure and the left ventricular mass, cause remo− deling of afferent arterioles, and lower the M/L ra− tio below normal levels [58].
An interesting experimental study performed by Tapia et al. investigated the effects of suppres− sing inflammation with mycophenolate mofetil on glomerular hemodynamics, arteriolar structural changes, and renal histologic injury in rats with subtotal renal ablation (with preservation of affe−
rent arteriolar function). Inflammation−associated arteriolopathy of the afferent arteriole could con− tribute to glomerular hemodynamic disturbances that participate in the progression of renal disease [59].
Arteriolar Injury
in Diabetes Mellitus
Hyalinization of juxtaglomerular arterioles is prominent in advanced diabetic nephropathy and may have important functional consequences, even in the early phase of diabetic nephropathy concomitant with early glomerulopathy [60]. Posi− tive correlation of the media thickness of afferent and efferent arteriolar profiles with albumin excre− tion rate and negative correlation with glomerular filtration rate (GFR) were observed in type I dia− betes mellitus with microalbuminuria [61]. In dia− betes, the differential expression of connexins in preglomerular and postglomerular vessels was al− so found [62]. According to Sugimoto et al. there is increased expression of endothelial cell nitric oxide synthase (eNOS) in afferent and glomerular endothelial cells, which contributes to glomerular hyperfiltration in diabetic nephropathy [63]. Mo− reover, neuronal oxide synthase (NOS1) plays an important role in the pathogenesis of hemodyna− mic changes in diabetes. It has been shown that ni− tric oxide modulates vascular tone in pre−glomeru− lar arterioles [64]. In diabetes mellitus, the role of immune mechanisms in the production of vascular lesions has been extensively studied, while less is known about their role at the level of the JGA.
Renal Arteriolar Injury in Vasculitis
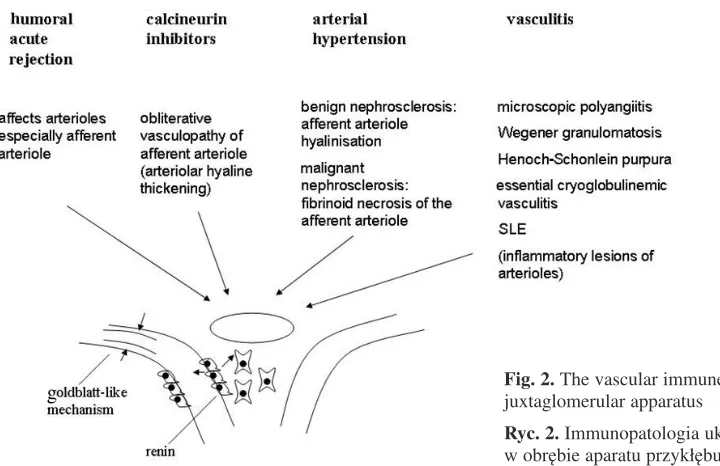
Injury of the arterioles of the JGA can impact immune−mediated diseases that are accompanied by arterial hypertension, such as vasculitis. Renal vessels are injured in vasculitis. Medium−sized vessels are mainly involved in macroscopic poly− angiitis (medium−sized vessel vasculitis), while small vessels, especially arterioles, are affected in microscopic polyangiitis and Wegener’s granulo− matosis (small−sized vessel vasculitis). In vasculi− tis, immune mechanisms are very important, with anti−neutrophil cytoplasmic antibodies (ANCAs) playing a major role. It is possible that ANCAs al− so affect JGA arterioles.
JGA vessels in the pathology of hypertension and renal lesions in this disease needs to be assessed.
Vasculitis is sometimes associated with severe hypertension. The relationship between hyperten− sion and arteriolar lesions of the JGA is not yet clear. The JGA vessel lesions during immune−me− diated diseases which affect the JGA could ampli− fy the severity of hypertension in these diseases. The treatment used to address the immune mecha− nisms involved in these diseases could have a be− neficial effect on the vessels of the JGA as well. Effects on the JGA arterioles in both vascular dise− ases that involve the entire vascular system and those that selectively involve the JGA arterioles may lead to the diminution of the lumens of these vessels. This reduction of lumen size may have consequences not only in terms of nephron func− tionality but also on AHT regulation in the entire organism. Studies performed by Gulmann et al. have revealed a negative correlation between the change in the luminal area of the afferent arteriole and the mean diastolic blood pressure in diabetic nephropathy [16].
It is a well known fact that diminution of the size of the renal artery, that is, renal artery stenosis, causes severe AHT. Experimentally, Goldblatt achieved severe hypertension by clamping the re− nal artery, reducing the blood flow in the renal cir− culation, including at the level of JGA arterioles, with increased production of renin. Activation of the renin−angiotensin− aldosterone system causes severe AHT. By analogy, the diminution of the size of the JGA vessels secondary to vascular lesions in the above−mentioned diseases (arterial hyperten− sion, atherosclerosis, vasculitis, etc.) may create
the conditions for deficient irrigation at the neph− ron level that renal artery stenosis produces. There− fore, instead of obtaining the Goldblatt effect thro− ugh the diminution of renal artery diameter, one could realize a sum of the Goldblatt effect through reduction of the circulatory flux in the afferent ar− terioles of the approximately two million nephrons. If achieving the experimental model of Gold− blatt is much easier than proving such a model at the level of JGA arterioles, many observations in pathology actually reveal lesions of the JGA arte− rioles, especially the afferent arterioles. Under some pathological conditions there have been ob− servations regarding the relationship between ischemia produced by vascular lesions in different diseases and the activation of the renin−angioten− sin−aldosterone system. Tapia et al. mention that the proliferation of media and intima in malignant hypertension causes the reduction of arteriolar lu− men size [59]. This reduction could lead to ische− mia, with activation of the renin−angiotensin−aldo− sterone system. Malignant hypertension is associa− ted with increased values of renin, angiotensin, and aldosterone [53]. Thus Gulmann et al. studied diabetes mellitus and managed to prove the rela− tionship between changes in the luminal area of the afferent arteriole and blood pressure (mean diastolic blood pressure) in diabetic nephropathy. More experimental and clinical studies are needed to support this finding.
However, the present authors consider it use− ful to present the main diseases in which lesions of the JGA arterioles, which may lead to diminished irrigation in these arterioles, have been reported. These diseases are accompanied by AHT, and
Fig. 2.The vascular immune pathology of the juxtaglomerular apparatus
a contribution by the reduction of the flux in the JGA arterioles cannot be excluded.
The vascular endothelium is a widely spread organ responsible for the regulation of the hemo− dynamic vascular remodeling and metabolic, syn− thetic, anti−inflammatory, and antithrombogenic processes [67]. Vascular lesions affect the arterio− lar endothelium, a fact that has been demonstrated in arterial hypertension and diabetes mellitus. En− dothelial dysfunction precedes renal injury. This has been demonstrated at the level of the afferent arteriole in spontaneously hypertensive rats fed a high−fat diet [68]. The involvement of the endo− thelium in immune processes at the level of the JGA was demonstrated experimentally by Juncos et al. They found variable degrees of endothelial lesions in the glomerular afferent arterioles of rab− bits perfused with medium containing the antibo− dy and complement [69].
Endothelial dysfunction could be regarded as the common mechanism that links CKD and car− diovascular diseases. Systemic blood pressure can be transmitted to the glomerular capillary network because of impairment of the autoregulatory me− chanisms existing in the afferent arteriole of the glomerulus. Glomerular hypertension and endo− thelial dysfunction are regarded as the common
mechanisms underlying the development of CKD [70]. The overproduction of nitric oxide (NO) has been reported in the diabetic kidney and is consi− dered to be involved in glomerular hyperfiltration. The enhanced NO synthesis by ecNOS in afferent arterioles and glomerular endothelial cells in re− sponse to the hyperglycemic state could cause pre− ferential dilation of afferent arterioles, which in− duces glomerular enlargement and glomerular hy− perfiltration [71].
At the level of the arterioles of the JGA, pres− sor substances such as angiotensin II or norepine− phrine can induce oxidative stress and consequent endothelial lesions. It has been demonstrated experimentally in rabbits that beta−1 receptors pro− tect the renal afferent arterioles of angiotensin−in− fused rabbits from norepinephrine−induced oxida− tive stress [72]. Oxidative stress plays an impor− tant role in producing endothelial lesions. It intervenes in the induction of vasopressor substan− ces. Thus infused angiotensin II in the afferent ar− teriole induces the formation of superoxide. Also, angiotensin II−infused rabbits develop oxidative stress in afferent arterioles upon administration of norepinephrine. Beta−1 receptors are protective against this effect, as demonstrated by the admini− stration of beta receptor stimulation [72].
Table 3. Vascular immune pathology of the JGA
Tabela 3. Immunologiczna patologia naczyń komórek okołokłębkowych
Disease Observed lesion Study type Authors
(Rodzaj choroby) (Zaobserwowane porażenie) (Rodzaj studium) (Autorzy) Acute humoral arteriolar fibrinoid necrosis human biopsies Mauiyyedi et al.
rejection 2002
(Ostre odrzucanie depositions at the level of peritubular capillaries of
humoralne) complement split product C4d human biopsies Feucht et al. 2005 Calcineurin inhibitor arteriolar hyaline thickening human biopsies Sis et al. 2006 treatment
(Leczenie inhibito− obliterative arteriopathy of the afferent arteriole human biopsies Busauschina et al.
rami kalcyneuryny) 2004
Arterial hypertension afferent arteriole hyalinization, benign experimental Wenzel et al. 1994
(Nadciśnienie tętnicze) nephrosclerosis model
fibrinoid necrosis of the afferent arteriole, malignant human biopsies Dikow and Ritz
nephrosclerosis 2003
narrowing of renal afferent arterioles adult spontaneously Skov and Mulvany hypertensive rats 2004
ACEIs cause remodeling of afferent arterioles genetically hyper− Ledingham and tensive rats Laverty 1998 Inflammation−associated arteriolopathy of the rats with subtotal Tapia et al. 2003
afferent arteriole renal ablation
Vasculitides Henoch Schonlein, injury of afferent arterioles, Jennette et al. 2005 (Zapalenie naczyń) severe hypertension
SLE – vasculitis involving afferent and efferent human biopsy Kozeny et al. 1986
Intrarenal RAS and the
Pathogenesis of Arterial
Hypertension in
Glomerular Nephropathies:
an Alternative to or
a Completion of the Role
of JGA in Producing
Arterial Hypertension
in These Diseases
The intrarenal RAS system could play an im− portant role in producing AHT in glomerular ne− phropathies [73]. Its elements are distributed at the level of the structures of the nephron. Thus in ma− ture kidneys, angiotensin II receptors have been localized in the luminal and basolateral mem− branes of several segments of the nephron, inclu− ding the glomeruli, arterial vasculature, vasa recta, arcuate arteries and juxtaglomerular cells, proxi− mal tubules, and the thick ascending limb of the loop of Henle [74]. Navar mentioned that angio− tensin II is compartmentalized in both a regional and a segmental manner. Its intrarenal disposition at the interstitial and tubular level must be mentio− ned [75]. Angiotensin II is compartmentalized in the renal interstitial fluid and the proximal tubular compartments at much higher concentrations than those existing in the circulation. According to Ko− bori, inappropriate activation of intrarenal RAS is an important contributor to the pathogenesis of hy− pertension and renal injury [73]. Intrarenal activa− tion of RAS is associated with local oxidative stress. This effect is ameliorated by ARB treat− ment. Intraglomerular activation of RAS has been evidenced in diabetic nephropathy. Activation of intrarenal RAS also occurs in various conditions of altered vascular volume. In some IgA nephropa− thies, Coppo mentions that local activation of RAS takes place in some cases [76]. Kobori et al. eva− luated 39 patients with IgA nephropathy and showed that immunoreactivity of intrarenal angio− tensinogen in IgA nephropathy was significantly increased compared with that in normal kidneys [73]. Mezzano et al. showed that immunohistoche− mical studies of renal biopsies from 20 patients with membranous nephropathy demonstrated ele− vated ACE and angiotensin II in tubular and inter− stitial cells [77].
A relationship between the intrarenal RAS sy− stem and AHT has not yet been demonstrated. In immune−mediated nephropathies, this link is po− ssible and needs to be investigated. The relation− ship of the RAS system with immune processes at
the level of the JGA must also be analyzed. The activation of intrarenal RAS has been observed in patients with essential hypertension [78].
Occasionally, the production of renin at the level of the JGA and the intrarenal level are in− creased concomitantly. Thus Tufro−McReddie fo− und in a rat model of cyclosporin A nephropathy that renin expression in the JGA was enhanced [74]. Intrarenal deposits of angiotensin II have been shown immunohistochemically in a model of chronic cyclosporine A toxicity in rats, which in− creased with time. This finding could suggest a complementary or alternative role of the JGA. Moreover, local intrarenal formation of angiotensin II was demonstrated by Admiraal et al. in renova− scular hypertension [79]. There is a concomitant increase in the production of renin in the JGA. The discussion can be raised regarding the complemen− tarity of the JGA with the RAS. Despite these re− sults, the role of the intrarenal RAS as an alternati− ve to or a completion of the role of JGA in produ− cing AHT is still debatable, and studies in humans to demonstrate this fact are difficult to perform. Al− though direct evidence of the augmentation of in− trarenal RAS in humans is relatively scarce, accu− mulating evidence including functional investiga− tions and studies of human biopsies suggest the augmentation of intrarenal RAS in human patients. From the data presented, it appears that the JGA is affected in diseases in which immune pro− cesses intervene, with a possible involvement of regulatory mechanisms of blood pressure. The in− trarenal RAS system is also frequently affected in these diseases. It is possible that in the production of AHT in these diseases, AHT is the consequence of the alteration of AHT regulation at the level of the JGA or at the level of the intrarenal RAS. In the meantime, these two mechanisms can be affec− ted concomitantly, and there may be a complemen− tarity between them. These phenomena are not yet elucidated, but deserve more attention in future studies that aim to solve these problems.
Prieto−Carasquero et al. mention that new understanding of the physiology and pathophysio− logy of intrarenal RAS provides a rationale for the use of ACE inhibitors and ARB combinations in the treatment of hypertension and prevention of the progression of kidney damage [80]. This conclusion can be extended to observations regarding the im− munopathology of the juxtaglomerular apparatus with respect to its involvement in AHT regulation.
Perspectives
which operate at the JGA level and may be pertur− bed as a consequence of immune processes that af− fect it. These include:
• the mechanism whereby the concomitant immune involvement of the JGA and the glomeru− lus participate in the production and progression of immune−mediated glomerular nephropathies,
• the injury of renin−producing cells during immune processes at the JGA level that could occur during arteriolar injury in vasculitis and other diseases,
• the relationship between immune processes in the JGA and nitric oxide production and prosta− glandin and cyclooxygenase activity,
• alteration of circulatory auto−regulation at the level of the kidney,
• vascular responses, including those of inju− red afferent and efferent arterioles, to hypotensive medication, including ACEIs and ARBs, which is especially important when the lesions advance. It may be possible in this situation to use another treatment, such as the direct blockade of renin with recent preparations.
In AHT, regardless of etiology, the participa− tion of lesions in the JGA vessels, including im− mune−induced ones, must be considered. AHT ther− apy has been suggested in light of its action upon these vessels, which have functional and structural characteristics. It is important to study the role of the extraglomerular mesangium, which establishes a communication between the macula densa and the JGA arterioles. Pathological mechanisms, in− cluding immune−mediated ones, which impact the extraglomerular mesangium, could alter JGA functionality. This process requires further atten− tion. Therapeutic perspectives should be esta− blished based on this body of knowledge, because the mechanism of AHT in glomerular immune ne− phropathies is unknown.
Conclusions
Immune processes at the level of the JGA have been identified which can affect its functionality.
The deposition of immune complexes at the level of the extraglomerular mesangium can alter the transmission of information from the level of the macula densa to the level of the afferent arte− rioles. The same effect could be due to the presen− ce of a cellular infiltrate.
Experimental studies performed by Ren on JGAs isolated from animals have demonstrated this very well. It has not yet been demonstrated in humans, but the presence of these immune depo− sits and the presence of an immune infiltrate could play a similar role.
The JGA intervenes in regenerative processes in immune−mediated nephropathies through the cells of the extraglomerular mesangium.
The immune vascular pathology involves ves− sels of the JGA, primarily the afferent arterioles. The narrowing of its lumen as well as the reduc− tion of the irrigation of the glomerulus can cause, as in renal artery stenosis, the activation of pressor mechanisms, primarily the renin−angiotensin aldo− sterone system, at this level, in contrast to renal ar− tery stenosis, in which Goldblatt described this fact. In these diseases, the phenomena could take place in a multitude of affected nephrons.
The study of immune processes at the level of the JGA, although only in the early stages, should open perspectives in the understanding of immune mechanisms in immune−mediated glomerular ne− phropathies. It also draws attention to immune− mediated lesions at the level of the JGA arterioles, which may represent an important link in the pa− thogenesis of hypertension.
References
[1] Yao J, Oite T, Kitamura M:Gap junctional intercellular communication in the juxtaglomerular apparatus. Am J Physiol Renal Physiol 2009, 296(5), F939–946.
[2] Kriz W, Eiger M:Renal Anatomy in Johnson RJ, Feehally J, Comprehensive clinical nephrology. Ed. Mosby. Edinburgh 2ndEdition, 2003, 9–10.
[3] Eiger M, Sakai T, Kriz W:The vascular pole of the renal glomerulus of rat. Adv. Anat Embryol Cell Biol 1998, 139, 1–98.
[4] Rosivall L, Mirzahoseini S, Toma I, Sipos N, Peti−Peterdi J:Fluid flow in the juxtaglomerular interstitium visu− alised in vivo. Am J Physiol Renal Physiol 2006, 291, 5, F1241–1247.
[5] Bachmann S, Oberbaumer S: Structural and molecular dissection of the juxtaglomerular apparatus: New aspects for the role of nitric oxide. Kidney Int 1998, 54, S29–S33.
[6] Bonsib SM: Renal anatomy and histology. In: Jennette JC, Olson JL, Schwartz MM, Silva FG. Heptinstall’s Pathology of the kidney. 6 ed., Volter Kluver/ Lippincott Williams and Wilkins, 2006, 42–44.
[7] Schweda F, Frus U, Wagner C, Skott O, Kurtz A: Renin release. Physiology 2007, 22, 310–319.
[9] Peti−Peterdi J:Confocal imaging and function of the juxtaglomerular apparatus. Curr Opin Nephrol Hypertens 2005, 14(1), 53–57.
[10] Bell PD, Lapointe JY, Peti−Peterdi J:Macula densa cell signaling. Ann Rev Physiol 2003, 65, 481–500.
[11] Sugimoto H, Shikata K, Matsuda M, Kushiro M, Hayashi Y, Hiragushi K, Wada J, Makino H: Increased expression of endothelial cell nitric oxide synthesis (eNOS) in afferent and glomerular endothelial cells is involved in glomerular hyperfiltration of diabetic nephropathy. Diabetologia 1998, 41, 12, 1426–1434.
[12] Krop M, Garrelds IM, de Bruin RJ, van Gool JM, Fisher ND, Hollenberg NK, Jan Danser AH: Aliskiren accumulates in Renin secretory granules and binds plasma prorenin. Hypertension 2008, 52(6), 1076–1083.
[13] Hugo C, Shankland SJ, Bowen−Pope DF, Couser WG, Johnson RL: Extraglomerular origin of the mesangial cells after injury. J Clin Invest 1997, 100, 786–794.
[14] Jennette JC, Falk RJ: Kidney involvement in systemic vasculitis, in Greenberg A, Cheung AK, Falk RJ, Coffman TM, and Jennette, Primer on Kidney Diseases, National Kidney Foundation, Elsevier Saunders, Philadelphia 2005, 226–260.
[15] Silva FG, Eigenbrodt EH, Glass M, Taft E: An ultrastructural study of the renal juxtaglomerular apparatus and extraglomerular mesangium in patients with systemic lupus erythematosus. Am J Kidney Dis 1986, 7, 1, 47–57.
[16] Hvala A, Kobenter T, Ferluga D: Fingerprint and other organized deposits in lupus nephritis. Wien Klin Wochenschr 2000, 112, 15–16, 711–715.
[17] Masson LJ, Isenberg DA: The pathogenesis of systemic lupus erythematosus, in Davison AM, Grunfeld JP, Ponticelli C, Ritz E, Winearls CG, van Ypersele C. Oxford Textbook of Clinical Nephrology, 3rd ed. Oxford
University Press, Oxford, 2005.
[18] Hara M, Honda K, Matsuya S, Endo Y, Hara S, Suzuki Y: The juxtaglomerular apparatus in IgA nephropathy: An analysis of the transport and fate of IgA deposits at the glomerular hilus. Virchows Archiv 1988, 413, 5, 431–433.
[19] Thompson EM, Evans DJ: Association of mesangial IgM with IgM deposits in the macula densa: an indication of non−specific macromolecule transport rather than immune reactant? Nephrol Dial Transplant 2001, 16, 1910–1913.
[20] Moriya R, Manivel JC, Mauer M: Juxtaglomerular apparatus T−cell infiltration affects glomerular structure in Type 1 diabetic patients. Diabetologia 2004, 47, 1, 82–88.
[21] Poulsen EL, Burke BA, Vernier RL, Mallare MJ, Innes DJ Jr, Sturgill BC: Juxtaglomerular body abnormal− ities in youth−onset diabetic subjects. Kidney Int 1994, 45, 4, 1132–1139.
[22] Johnson PA, Alexander HD, McMillan SA, Maxwell PP:Upregulation of the endothelial cell adhesion mole− cule intercellular adhesion molecule−1 (ICAM−1) by autoantibodies in autoimmune vasculitis. Clin Exp Immunol 1997, 108, 234–242.
[23] Muller Kobold AC, van Wijk RT, Fransen CPM, Molema G, Kallenberg CG, Tervaert JW:In vitroupre− gulation of E selectin and induction of interleukin 6 in endothelial cells by autoantibodies in Wegener’s granulo− matosis and microscopic polyangeitis. Clin Exp Rheumatol 1999, 17, 433–440.
[24] Mavrikakis ME, Vaiopoulos G, Papantoniou B, Antoniades LG, Kostopoulos C, Papazoglou S, Lianos EA:
Plasma renin activity as a marker of renovascular injury in patients with rheumatoid arthritis. Clin Exp Rheumatol 1996, 14, 6, 613–617.
[25] Goligorsky MS: Frontiers in nephrology: viewing the kidney through the heart–endothelial dysfunction in chron− ic kidney disease. J Am Soc Nephrol 2007, 18(11), 2833–2835.
[26] Yao J, Oita T, Kitamura M:Gap junctional intercellular communication in the juxtaglomerular apparatus. Am J Physiol Renal Physiol 2009, 296, 5, F939–946.
[27] Wetzel R, Sweadner KJ:Phospholemman expression in extraglomerular mesangium and afferent arteriole of the juxtaglomerular apparatus. Am J Physiol Renal Physiol 2003, 285, 1, F121–129.
[28] Ren Y, Carretero OA, Garvin JL:Role of mesangial cells and gap junctions in tubuloglomerular feedback. Kidney Int 2002, 62, 525–531.
[29] Haseley LA, Hugo C, Reidy MA, Johnson RJ:Dissociation of mesangial cell migration and proliferation in experimental glomerulonephritis: Kidney Int 1999, 56, 3, 964–972.
[30] Gluhovschi C, Gluhovschi G, Potencz E, Herman D, Petrica L, Velciov S, Bozdog G, Bob F, Vernic C, Cioca D: What is the significance of CD34 mmunostaining in the extraglomerular and intraglomerular mesangium? Virchows Arch 2008, 453(4), 321–328.
[31] Baddour N, Adam AG, El Koraie AH, El Nahas EM:The repairative role of stem cells in human glomeru− lonephritis XXXIX Congress of the European Renal Association European Dialysis and Transplant Association. July 14–17 2002 Copenhagen, Denmark Nephrol Dial Transplant 2002, vol 17 Abstracts, Suppl. 1, 9–10.
[32] Naruse K, Fujieda M, Miyazaki E, Hayashi Y, Kuroda N, Nakayama N, Kiyoku H, Hiroi M, Karashige T, Enzan H: CD34 expression as a novel marker of transformed mesangial cells in biopsied glomerular disease. J Pathol 1999, 189, 105–111.
[33] Ito T: Stem cells of the adult kidney: where are you from? Nephrol Dial Transplant 2003, 18, 4, 641–644.
[34] Oliver JA, Maarouf O, Cheema FH, Martens TP, Al−Awquati Q:The renal papilla is a niche for adult kidney stem cells. J Clin Invest 2004, 114, 794–804.
[36] Poulsom R, Alison MR, Cook T, Jeffery R, Rzan E, Forbes SJ, Hunt T, Wyles S, Wright NA:Bone marrow stem cells contribute to healing of the kidney. J Am Soc Nephrol 2003, 14, 5, 48–554.
[37] Imasawa T, Utsunomiya Y, Kawamura T, Zhong Y, Nagasawa R, Okabe M, Maruyama N, Hosoya T, Ohno T:The potential of bone marrow−derived cells to differentiate to glomerular mesangial cells. J Am Soc Nephrol 2001, 12, 1401–1409.
[38] Imasawa T, Nagasawa R, Utsunomiya Y, Kawamura T, Zhong Y, Makita N, Muso E, Miyawaki S, Maruyama N, Hosoya T, Sakai O, Ohno T: Bone marrow transplantation attenuates murine IgA nephropathy: role of a stem cell disorder. Kidney Int 1999, 56, 1809–1817.
[39] Iwatani H, Ito I, Imai E, Suzuki A, Ueda N, Hori M:In vitroconversion of bone marrow cells into mesangial− like cells. J Am Soc Nephrol 2001, 16, 520 A.
[40] Masuya M, Drake CJ, Fleming PA, Reilly CM, Zeng H, Hill WD, Martin−Studdard A, Hess DC, Ogawa M:
Hematopoietic origin of glomerular mesangial cells. Blood 2003, 101, 6, 2215–2218.
[41] Kunter U, Rong S, Djuric Z, Boor P, Mueller−Newen G, Yu D, Floege J: Transplanted mesenchymal stem cells accelerate glomerular healing in experimental glomerulonephritis. J Am Soc Nephrol 2006, 17(8), 2202–2212.
[42] El Nahas AM: Plasticity of kidney cells: role in kidney remodeling and scarring. Kidney Int 2003, 64, 5, 1553–1465.
[43] Ito T, Suzuki A, Imai E, Okabe M, Hori M: Bone marrow is a reservoir of repopulating mesangial cells during glomerular remodeling. J Am Soc Nephrol 2001, 12, 2625–2635.
[44] Suzuki A, Ivatani H, Ito T, Imai E, Okabe M, Nakamura H, Isaka Y, Yamato M, Hori M:Platelet−derived growth factor plays a critical role to convert bone marrow cells into glomerular mesangial−like cells. Kidney Int 2004, 65, 15–24.
[45] Baud L, Haymann JP, Bellocq A, Fouqueray B:Contribution of stem cells to renal repair after ischemia/ reper− fusion. Bull Acad Natl Med 2005, 189, 4, 635–643.
[46] Racusen LC, Haas M:Antibody−mediated rejection in renal allografts. Lessons from Pathology. Clin J Am Soc Nephrol 2006, 1(3), 415–420.
[47] Vella JP, Danovitch GM:Syllabus Transplantation. Nephrology Self−Assessment Program 2006, 5, 4, 201–210.
[48] Mauiyyedi S, Crespo M, Bernard Collins A, Schneeberger EE, Pascual MA, Saidman SL, Tolkoff−Rubin NE, Williams WW, Delmonico FL, Cosimi AB, Colvin RB: Acute humoral rejection in kidney transplantation: II. Morphology, Immunopathology, and Pathologic Classification. J Am Soc Nephrol 2002, 13, 779–787.
[49] Feucht HE, Mihatsch MJ:Diagnostic value of C4d in renal biopsies. Curr Opin Nephrol Hypertens 2005, 14(6), 592–598.
[50] Sis B, Dadras F, Khoshjou F, Cockfield S, Mihatsch MJ, Solez K:Reproducibility studies on arteriolar hyaline thickening scoring in calcineurin inhibitor−treated renal allograft recipients. Am J Transplant 2006, 6, 1444.
[51] Busauschina A, Schnuelle P, van der Woude FJ:Cyclosporine nephrotoxicity. Transplant Proc 2004, 36 (2), 229S–233S.
[52] Wenzel UO, Helmchen U, Schoeppe W, Schwietzer G:Combination treatment of enalapril with nitrendipine in rats with renovascular hypertension. Hypertension 1994, 23(1), 114–122.
[53] Cotran RS, Kumar V, Robins SL.In Robins pathologic basis of disease 5thed. Ed Saunders Philadelphia 1994,
976–978.
[54] Razga Z, Ivanyi B, Zidar N, Ferluga D, Sonkodi S, Ormos J: Quantitative ultrastructural study of afferent and efferent arterioles in IgA glomerulonephritis and benign nephrosclerosis. Virchows Arch 1996, 429, 275–281.
[55] Dikow R, Ritz E:Cardiovascular complications in the diabetic patient with renal disease: an update in 2003. Nephrol Dial Transplant 2003, 18(10), 1993–1998.
[56] Tracy RE, Berenson GS, Cueto−Garcia LS, Wattigney WA, Barrett TY:Nephrosclerosis and aortic athero− sclerosis from age 6 to 70 years in the United States and Mexico. Virchows Archiv. Pathological Anatomy and Histopathology 1992, 420, 479–488.
[57] Skov K, Mulvany MJ: Structure of renal afferent arterioles in the pathogenesis of hypertension. Acta Physiol Scand 2004, 181(4), 397–405.
[58] Ledingham JM, Laverty R:Renal afferent arteriolar structure in the genetically hypertensive (GH) rat and the ability of losartan and enalapril to cause structural remodeling. J Hypertens 1998, 16, 1945–1952.
[59] Tapia E, Franco M, Sanchez−Lozada LG, Soto V, Avila−Casado C, Santamaría J, Quiroz Y, Rodríguez− Iturbe B, Herrera−Acosta J: Mycophenolate mofetil prevents arteriolopathy and renal injury in subtotal ablation despite persistent hypertension. Kidney Int 2003, 63(3), 994–1002.
[60] Osterby R, Bongstad HJ, Nyberg G, Walker JD, Viberti G: A quantitative ultrastructural study of juxta− glomerular arterioles in IDDM patients with micro and normoalbuminuria. Diabetologia 1995, 38, 11, 1320–1327.
[61] Gulmann C, Osterby R, Bongstad HG, Rutberg S: The juxtaglomerular apparatus in young type 1 diabetic patients with microalbuminuria. Effect of antihypertensive treatment. Virchows Arch 2001, 438, 6, 618–623.
[62] Zhang J, Hill CE:Differential connexin expression in preglomerular and postglomerular vasculature: accentua− tion during diabetes. Kidney Int 2005, 68(3), 1171–1185.
[63] Sugimoto H, Shikata K, Matsuda M, Kushiro M, Hayashi Y, Hiragushi K, Wada J, Makino H: Increased expression of endothelial cell nitric oxide synthase (eNOS) in afferent and glomerular endothelial cells is involved in glomerular hyperfiltration of diabetic nephropathy. Diabetologia 1998, 41(12), 1426–1434.