Journal of Biological Sciences and Medicine
Available online at www.jbscim.com
ISSN: 2455-5266
1
Research Article
Open Access
Optimization of Process Parameters in Submerged Fermentation
for Lipase Production from Soil Fungal Isolate
Arun Kumar Sharma
1, Sapna Kumari
1, Vinay Sharma
1*and Jyoti Saxena
21
Department of Bioscience and Biotechnology, Banasthali University, Rajasthan, India
2
Department of Biochemical Engineering, Bipin Tripathi Kumaon Institute of Technology, Dwarahat, Uttrakhand.
*Corresponding author: [email protected]
ARTICLE INFO ABSTRACT
Article History:
Received
13 May 2017
Accepted
20 June 2017
Available online
30 June 2017
Lipases are very important industrial enzymes. They are used in number of industries including paper, detergent, food, dairy, pharmaceutical, diagnostic centre, cosmetic etc. By considering immense applications of lipases, an attempt was made to optimize lipase production in submerged fermentation (SmF) from a fungal isolate. In the present study, various parameters such as carbon source, nitrogen source, pH, temperature and incubation time were optimized for a lipolytic fungal strain named as S2St1, previously isolated from mustard field soil sample, for lipase production in SmF. Maximum activity of lipase (157.28±4.26 U/ml/min) was obtained at 28 ºC, after 7 days of incubation in the medium containing mixture of fructose (0.5% w/v) + coconut oil (1% v/v) as carbon source and peptone (4% w/v) + (NH4)2SO4 (0.1% w/v) as nitrogen
source. The present study reveals that coconut oil was found to be best among other lipid carbon source for lipase production, which indicates that presence of inducer (oil) is required for maximum lipase activity. Lipase activity was also found better at 50 ºC (65.77±5.39 U/ml/min), indicating superior thermo-stability of fungal strain for lipase production.
Key words:
Submerged fermentation;
Optimization; Lipases; Carbon source; Nitrogen source
Copyright: © 2017 Sharma et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Lipases are biotechnologically imperative group of enzymes which act on the ester bonds present in triacylglycerol and liberate fatty acids and glycerol (Abrunhosa et al. 2013). Lipases not only catalyze hydrolysis reaction but also capable of catalyzing range of reactions such as esterification, transesterification (alcoholysis, aminolysis, acidolysis, interesterification). They catalyze hydrolytic reaction in aqueous environment and other reactions (esterification, transesterification) in non-aqueous environment (Charles and James 2011).
2 treatment for removal of lipid clogged drains and in the medical industry for estimation of blood lipid contents (Verma and Prakash 2014).
Lipases are produced from a variety of living organisms such as plants, animals, bacteria, fungi, yeasts, actinomycetes. Amongst all, microbial sources of lipase have gained industrial attention in past few years because of unique properties and chemical stability (Andualema and Gessesse 2012). Fungal lipase production has been reported in different species of Geotrichum,
Candida, Aspergillus, Humicola, Mucor,
Pencillium and Rhizopus genus (Gopinath et al. 2002; Cihangir and Sarikaya 2004; Prabhakar et al. 2012). Lipase producing microbes can be found in different habitats such as industrial effluents, edible oil processing industries, dairies, soil contaminated with edible oil, diesel oil, oilseeds, and decaying food (Sztajer and Zboinska 1988).
Enhancement of lipase production requires strain improvement and optimization studies. Lipase production is normally influenced by culture medium ingredients and cultivation conditions (Immanuel et al. 2008). By varying culture medium ingredients, an increase in amount of lipase enzyme can be maintained. In addition, by considering enormous applications of lipases, an attempt has been made to increase lipase production in submerged fermentation (SmF) from lipolytic soil fungal isolate by optimization studies.
Materials and methods
Microorganism
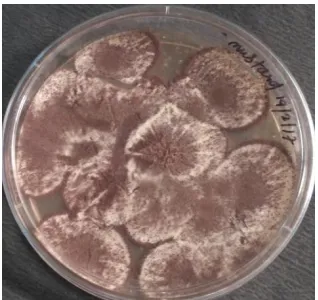
Lipolytic soil fungal isolate (named as S2St1, isolated from mustard field soil sample) (Fig. 1) was obtained from Department of Bioscience and Biotechnology, Banasthali University, Rajasthan. Culture was maintained on potato dextrose agar (PDA) slants.
Fig. 1: Lipolytic fungal isolate (S2St1)
Lipase production in SmF
For lipase production, 1 ml of fungal spore suspension (made from 6 days old slat culture of fungi) was transferred in 100 ml of autoclaved production medium containing (g/1000 ml): bacteriological peptone, 40; olive oil, 10 ml; MgSO4.7H2O, 1; KH2PO4, 1; (NH4)2SO4, 1;
sucrose, 5 and pH was adjusted to 6.5 (Xia et al. 2011). Inoculated flasks were placed in a shaker incubator at 28 ºC, 150 rpm for 3 days. At the end of incubation lipase activity was determined from mycelium free culture supernatant.
Lipase assay
Activity of lipase was measured by spectrophotometric technique using p-nitrophenyl palmitate as substrate as explained previously by Winkler and Stuckmann (1979).
Protein quantification
Total protein contents were determined by method given by Lowry (1951).
Optimization of culture conditions for increased lipase production
3 Effect of different carbon sources on lipase production
To study the impact of various carbon sources on lipase production by isolate S2St1, original carbon sources (olive oil (1% v/v) and sucrose (0.5% w/v)) of production medium were replaced by combination of other carbon sources such as soybean oil + glucose, almond oil + lactose, coconut oil + fructose, and mustard oil + starch at a concentration of 1% v/v oil and 0.5% w/v carbohydrate source. One flask was kept as control containing sucrose + olive oil as carbon sources. Spore suspension was prepared and inoculated in 100 ml of production medium containing different combination of carbon sources followed by incubation for 4 days at 28 ºC, 150 rpm. Mycelium free supernatant was recovered from fermentation broth after 3 days and 4 days of incubation and used for estimation of lipase activity and protein content (Sharma et al. 2016a).
Effect of different nitrogen sources on lipase production
Best carbon sources from previous experiment were used for preparation of culture medium for this study. Original nitrogen sources (peptone (4% w/v) and (NH4)2SO4 (0.1 % w/v)) of
production medium were replaced by combination of other nitrogen sources such as casein + (NH4)2SO4, peptone + (NH4)2SO4, beef
extract + KNO3, yeast extract + (NH₄)₂S₂O₈ and
skim milk + ammonium acetate at a concentration of 4% w/v organic nitrogen and 0.1% inorganic nitrogen source. One flask was kept as control containing sucrose + olive oil as carbon sources and peptone + (NH4)2SO4 as nitrogen sources
(Sharma et al. 2016b). Flasks were inoculated with spore suspension and lipase activity and protein content were determined at 3 days and 4 days of incubation in culture supernatant.
Effect of different pH on lipase production
Previously optimized carbon and nitrogen sources were used for media preparation in this study. pH of the production medium (containing fructose +
coconut oil and peptone + (NH4)2SO4) was
adjusted in the range of 4 to 10 and lipase production was estimated at 3 days and 4 days of incubation (Sharma et al. 2015a). One flask was kept as control containing sucrose + olive oil as carbon sources and peptone + (NH4)2SO4 as
nitrogen sources and pH was 6.5. Lipase activity and protein content were determined at 3 days and 4 days of incubation.
Effect of different temperatures on lipase production
Previously optimized carbon sources, nitrogen sources and pH were used for media preparation in this study. Fermentation broth (fructose + coconut oil and peptone + (NH4)2SO4) was
prepared and after inoculation with spore suspension, flasks were incubated at different temperatures: 28 ºC, 37 ºC and 50 ºC (Sharma et al. 2015b). Three control flasks were also prepared (sucrose + olive oil and peptone + (NH4)2SO4) and incubated at 28 ºC, 37 ºC and 50
ºC, respectively. Lipase activity and protein content were determined at 3 days of incubation.
Effect of incubation period on lipase production
Previously optimized conditions (fructose + coconut oil, peptone + (NH4)2SO4, pH 8.0,
temperature 28 ºC) were used for preparation of culture media. Spore suspension was inoculated in fermentation broth followed by incubation for 6 days at optimized temperature. Lipase activity and protein content were determined at following days of incubation: day 1, 2, 4, 5 and 7.
Results
Lipase production in SmF
Lipolytic fungal isolate (named as S2St1, isolated from mustard field soil sample) demonstrated 98.33±4.53 U/ml/min at 3rd day of incubation and
150.76±2.06 U/ml/min at 4th day of incubation in the medium containing olive oil and sucrose as carbon sources and peptone and (NH4)2SO4 as
4 lipase gene hence, activity of lipase was increased. It indicated inducible nature of lipases.
Influence of carbon sources on lipase production
The highest activity of lipase was obtained at day 3 (124±4.65 U/ml/min) and day 4 (157.48±2.90 U/ml/min) of incubation when the medium was fed with the mixture of fructose and coconut oil as compared to other carbon sources (Table 1). Fructose is a monosaccharide that supported the growth of the fungus, while coconut oil was efficiently used by the fungus for maximum lipase production. Lipase activity was lower in presence of lactose and almond oil. At day 3, least activity was obtained with starch + mustard oil. It was probably due to the fact that starch was not efficiently used by the fungus for growth hence, the lipase activity declined.
Our results suggest that presence of single carbon source (carbohydrate) in production medium is not sufficient for higher lipase production, addition of lipidic carbon source (oil) is also required which act as an inducer for lipase production.
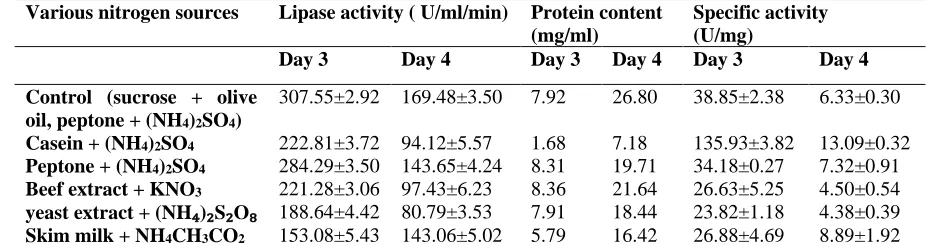
Influence of nitrogen sources on lipase production
Combination of peptone and (NH4)2SO4 was
found optimum nitrogen source and increased lipase activity to 284.29±3.50 U/ml/min at 3rd day
of incubation and 143.65±4.24 U/ml/min at 4th day of incubation (Table 2). In the present study, all the nitrogen sources were efficiently used by the fungus for growth and accumulation of lipase. Our study suggests that combination of inorganic and organic sources of nitrogen is required for growth of the fungus and production of the lipase.
Table 1: Influence of carbon sources on specific activity and lipase production from lipolytic fungal isolate (S2St1).
Carbon source optimization Lipase activity ( U/ml/min)
Protein content (mg/ml)
Specific activity (U/mg)
Day 3 Day 4 Day 3 Day 4 Day 3 Day 4
Control (Olive oil + sucrose) 98.33±4.53 150.76±2.06 21.47 18.96 4.75±0.66 8.39±0.75
Soyabaen oil + glucose 46.56±3.23 104.04±3.10 21.25 20.47 2.20±0.38 5.10±0.94
Almond oil + lactose 42.27±1.28 45.13±3.12 21.11 20.4 1.98±0.54 2.21±0.21
Coconut oil + fructose 124±4.65 157.48±2.90 21.99 19.66 5.64±0.13 8.04±0.68
Mustard oil + starch 39.40±3.85 106.37±4.29 20.61 19.53 1.90±0.65 5.43±0.37
Nitrogen sources (peptone + (NH4)2SO4): constant; pH: constant; temperature: constant.
Table 2: Influence of nitrogen sources on specific activity and lipase production from lipolytic fungal isolate (S2St1).
Various nitrogen sources Lipase activity ( U/ml/min) Protein content (mg/ml)
Specific activity (U/mg)
Day 3 Day 4 Day 3 Day 4 Day 3 Day 4
Control (sucrose + olive oil, peptone + (NH4)2SO4)
307.55±2.92 169.48±3.50 7.92 26.80 38.85±2.38 6.33±0.30
Casein + (NH4)2SO4 222.81±3.72 94.12±5.57 1.68 7.18 135.93±3.82 13.09±0.32
Peptone + (NH4)2SO4 284.29±3.50 143.65±4.24 8.31 19.71 34.18±0.27 7.32±0.91
Beef extract + KNO3 221.28±3.06 97.43±6.23 8.36 21.64 26.63±5.25 4.50±0.54
yeast extract + (NH₄)₂S₂O₈ 188.64±4.42 80.79±3.53 7.91 18.44 23.82±1.18 4.38±0.39
Skim milk + NH4CH3CO2 153.08±5.43 143.06±5.02 5.79 16.42 26.88±4.69 8.89±1.92
5 Influence of pH on lipase production
The maximum activity of lipase (179.55±5.06 U/ml/min) was obtained at day 3 of incubation when the pH of fermentation broth was adjusted to 8.0 (Table 3). At day 3, lipase activity was maximum at pH 8.0 followed by 7.0, 6.5, 5.0, 4.0, 9.0 and 10.0. Lipase activity was higher at neutral pH (7.0), slightly acidic pH (6.5, 5.0, and 4.0) and slightly alkaline pH (8.0) but it was drastically declined when the pH of production medium was adjusted in alkaline range (pH 9.0-10.0). The activity was higher in the acidic pH range (4.0 and 5.0) than in alkaline pH range (9.0-10.0), indicating that acidic environment promote the growth of the fungus and accumulation of lipase.
Our results suggest that lipase production is very much affected by external pH fluctuations.
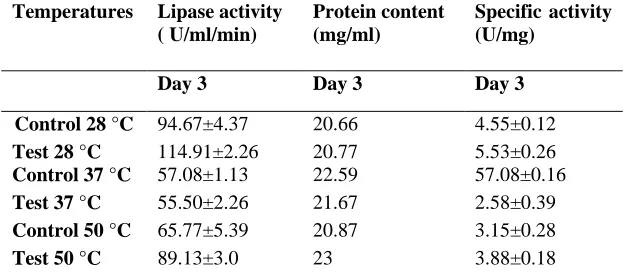
Influence of temperature on lipase production
Among all temperatures, highest activity of test (114.91±2.26 U/ml/min) and control (94.67±4.37 U/ml/min) was obtained when incubation temperature was 28 ºC. Optimum temperature was 28 ºC followed by 50 ºC and 37 ºC (Table 4). Lipase activity was higher at 50 ºC (89.13±3.0 U/ml/min) than 37 ºC (55.50±2.26 U/ml/min) indicating that fungus is capable of synthesizing good quantity of lipase at higher temperature also. Our results suggest that fungus (S1St2) can be used for lipase production at higher temperature. It indicates the thermostable nature of the fungus.
Table 3: Influence of external pH on specific activity and lipase production from lipolytic fungal isolate (S2St1).
Various pH Lipase activity ( U/ml/min)
Protein content (mg/ml)
Specific activity (U/mg)
Day 3 Day 4 Day 3 Day 4 Day 3 Day 4
Control (6.5) 165.13±6.27 172.88±3.98 20.58 20.63 8.03±0.53 8.38±0.37
pH 4 44.39±2.3 77.82±4.5 21.86 19.79 2.04±0.92 3.94±0.71
pH 5 58.71±4.93 74.27±5.2 20.19 19.39 2.91±0.30 13.82±0.50
pH 7 173.58±6.71 181.67±5.87 20.66 19.37 8.39±0.28 9.41±0.82
pH 8 179.55±5.06 173.53±5.33 18.88 18.34 9.55±0.89 9.52±1.07
pH 9 21.28±2.57 45.38±2.58 22.72 22.60 0.93±0.11 2.01±0.44
pH 10 12.09±0.44 57.18±4.6 24.02 23.78 0.48±0.37 2.40±0.18
Control: sucrose + olive oil, peptone + (NH4)2SO4; Test: pH 4 to 10 (fructose + coconut oil, peptone +
(NH4)2SO4).
Table 4: Influence of temperatures on specific activity and lipase production from lipolytic fungal isolate (S2St1).
Temperatures Lipase activity ( U/ml/min)
Protein content (mg/ml)
Specific activity (U/mg)
Day 3 Day 3 Day 3
Control 28 °C 94.67±4.37 20.66 4.55±0.12
Test 28 °C 114.91±2.26 20.77 5.53±0.26
Control 37 °C 57.08±1.13 22.59 57.08±0.16
Test 37 °C 55.50±2.26 21.67 2.58±0.39
Control 50 °C 65.77±5.39 20.87 3.15±0.28
Test 50 °C 89.13±3.0 23 3.88±0.18
6 Influence of incubation period on lipase production
The lipase activity was gradually increased with the rise of incubation period from 48 h (25.67±3.19 U/ml/min), 72 h (49.77±4.91 U/ml/min), 120 h (120.19±3.76 U/ml/min) and reached to maximum (157.28±4.26 U/ml/min) at 7 days of incubation (Table 5). Specific activity was also higher (8.32±1.20 U/mg) at 7 days of incubation. Specific activity is the amount of lipase within the mixture of contaminating proteins. Our results suggest that fungus slowly utilizes the culture medium ingredients, therefore able to accumulate large amount of lipase even at 7th day of incubation.
Discussion
Dahiya and Purkayastha (2011) reported that highest lipase production (4.25±0.020 U/ml) was obtained with olive oil as a carbon source followed by coconut oil (2.5±0.030 U/ml). Supakdamrongkul et al. (2010) have revealed highest lipase activity from fungus using castor oil (30.5±0.6 U mL−1), followed by coconut oil (20.8±0.4 U mL−1), olive oil (20.8±0.4 U mL−1) and cottonseed oil (20.6±0.4 U mL−1).
Niaz et al. (2013) reported almond meal as best substrate for optimum lipase activity (12.25±0.11 U/mL/min) from Trichophyton sp. Sopuruchukwu et al. (2015) reported sucrose at a concentration of 1.5 % as best carbon source for optimum production of lipase (7.7 U/ml/min) from Fusarium sp. Prasad (2013) observed
maximum lipase activity (6.102 U/ml) in the presence of glucose and starch as carbon sources.
Prasad (2013) observed maximum lipase activity (8.441 U/ml) in presence of casein as nitrogen sources. Maia et al. (1999) have documented maximum lipase activity (10500 U/L) from Fusarium sp. in presence of peptone (3% w/v) and olive oil (0.5v/v) in the medium. Ire and Ike (2014) reported highest lipase production from Aspergillus carbonarius with peptone (2% w/v) and glucose (2% w/v) in production medium. Peptone was also reported as best nitrogen source by Ulker et al. (2011) for
Trichoderma harzianum.
Lopes et al. (2016) reported ammonium sulfate as best nitrogen source for optimum lipolytic activity from Yarrowia lipolytica. Sopuruchukwu et al. (2015) reported ammonium chloride as best nitrogen source at a concentration of 2.5 % for optimum production of lipase (23.6 U/ml/min) from Fusarium sp. Ammonium chloride was also reported as best nitrogen source by D’annibale et al. (2006) for maximum lipase production from
Penicillium citrinum. Dahiya and Purkayastha (2011) reported that ammonium nitrate produced maximum lipase activity (15.6±0.036 U/ml).
In agreement with the present results, Maia et al. (1999) and Supakdamrongkul et al. (2010) reported maximum lipase production from fungus at pH 8.0. An optimum pH of 7.0 was reported by Adinarayana et al. (2003) for Aspergillus sp. and Costa and Peralta (1999) for Penicillium sp.
Table 5: Influence of incubation time on specific activity and lipase production from lipolytic fungal isolate (S2St1).
Carbon source: fructose + coconut oil; nitrogen source: peptone + (NH4)2SO4; pH: 8.0 and temperature: 28ºC.
Incubation time (days)
Lipase activity ( U/ml/min)
Protein content (mg/ml)
Specific activity (U/mg)
1 30.76±4.38 24.47 1.25±0.18
2 25.67±3.19 19.23 1.33±0.37
4 49.77±4.91 20.60 2.43±0.50
5 120.19±3.76 23.92 5.02±0.17
7 Ire and Ike (2014) revealed optimum pH of 6.0 for Aspergillus carbonarius. On the other hand, Sopuruchukwu et al. (2015) reported optimum lipase production from Fusarium sp. when pH of fermentation broth was in acidic range pH 2.5. Moderate lipase production was at pH 5.0-6.0, while lipase activity was declined in alkaline pH above 8.0.
Similar to our results, an optimum temperature of 30 ºC was reported by Sopuruchukwu et al. (2015) and Maia et al. (1999) for Fusarium sp. and Ire and Ike (2014) for Aspergillus carbonarius. Supakdamrongkul et al. (2010) have reported an optimum temperature of 25 ºC for lipolytic activity from fungus. An optimum temperature of 40 ºC was reported by Dahiya and Purkayastha (2011) and 45 ºC by Costa and Peralta (1999) for Penicillium sp.
Similar to our results, Costa and Peralta (1999) have reported optimum lipase activity from Penicillium sp after 7 days of incubation in presence of olive oil (5%). Sopuruchukwu et al. (2015) reported maximum lipase production from
Fusarium sp. after 120 h of incubation. An optimum incubation period of 96 h was reported by Adinarayana et al. (2003) for Aspergillus sp. and Ire and Ike (2014) for Aspergillus carbonarius. While Maia et al. (1999) reported highest lipase activity from Fusarium sp. after 72 h of incubation.
Conclusion
The current investigation has been taken up with a view of exploring the probabilities of using lipolytic fungal isolate as a source for production of industrially imperative lipase enzyme. Maximum production of lipase was observed at 28 ºC after 7 days of incubation at pH 8.0, when the medium was supplemented with mixture of fructose + coconut oil as carbon source and mixture of peptone + ammonium sulfate as nitrogen source. Results of optimization demonstrated that fungus is capable of producing increased amount of lipase under optimized conditions, therefore can be used as new source
of industrial lipase production. It is hoped that after some scale up studies, our fungal strain can be used for mass cultivation and production of large quantities of lipase in SmF.
Conflict of interest
Authors declare no conflict of interest.
Acknowledgements
Authors are grateful to Professor Aditya Shastri, Vice-Chancellor, Banasthali University for providing research facilities in the Department.
References
Abrunhosa L, Oliveira F, Dantas D, Goncalves C, Belo I (2013) Lipase production by Aspergillus ibericus using olive mill wastewater. Bioproc Biosyst Engin 36: 285-291
Adinarayana K, Bap Raju KVVSN, Zargar MI, Devi RB, Lakshmi PJ, Ellaiah P (2003) Optimization of Process Parameters for Production of Lipase in Solid State Fermentation by newly isolated Aspergillus species. I J Biotechnol 3: 65-69
Andualema B and Gessesse A (2012) Microbial lipases and their industrial applications: review. Biotechnology 11: 100-118
Charles ON and James CO (2011) Isolation of lipase producing fungi from palm oil Mill effluent (POME) dump sites at Nsukka. Braz Arch Biol Technol 54: 113-116
Cihangir N and Sarikaya E (2004) Investigation of lipase production by a new isolated of Aspergillus sp. World J Microbiol Biotech 20: 193-197
Costa MA and Peralta RM (1999) Production of lipase by soil fungi and partial characterization of lipase from a selected strain (Penicillium wortmanii). J Basic Microbiol 39: 11-15
D’Annibale A, Sermanni GG, Federici F, Petruccioli M (2006) Olive-mill wastewaters: a promising substrate for microbial lipase production. Bioresour Technol 97: 1828-1833
Dahiya P, and Purkayastha S (2011) Isolation, Screening and Production of Extracellular Alkaline Lipase from a Newly Isolated Bacillus sp. PD-12. J Biol Sci 11: 381-387
Ferreira-Dias S, Georgina S, Francisco P, Francisco V (2013) The potential use of lipases in the production of fatty acid derivatives for the food and nutraceutical industries. Electronic J Biotechnol 16 Gopinath SCB, Hilda A, Priya TL, Annadurai G (2002)
8
using response surface methodology. World J Microbiol Biotech 18: 449-458
Immanuel G, Esakkiraj P, Jebadhas A, Iyapparaj P, Palavesam A (2008) Investigation of lipase production by milk isolate Serratia rubidaea. Food Technol Biotechnol 46: 60-65
Ire FS and Ike VC (2014) Screening and Optimization of Process Parameters for the Production of Lipase in Submerged Fermentation by Aspergillus carbonarius (Bainer) IMI 366159. Annu Res Rev Biol 4: 2587-2602
Lopes VRO, Farias MA, Belo IMP, Coelho MAZ (2016) Nitrogen Sources on Tpomw Valorization through Solid State Fermentation Performed by Yarrowia Lipolytica. Braz J Chem Eng 33: 261-270
Lowry OH, Rosenbrough NJ, Farr AL, Randall A (1951) Protein measurement with the folin phenol reagent. J Biol Chem193: 265-275
Maia MDMD, Morais MMCD, Morais MAD, Melo EHM, Filho JLDL (1999) Production of extracellular lipase by the phytopathogenic fungus Fusarium solani FS1. Revista de Microbiologia 30: 304-309
Niaz M, Iftikhar T, Akram F, Niaz M (2013) Protocol Optimization for Extracellular Lipase Production by Trichophyton Spp. (Mbl 23) under Solid State Fermentation. Pak J Bot 45: 1417-1421
Prabhakar T, Bhogavalli PK, Vallem PR, Srirangam V (2012) Studies on optimization of extracellular lipase from potential fungal strain(s) isolated from oil contaminated soil. J Microbiol Biotechnol Res 2: 418-425
Prasad MP (2013) Production of extracellular lipase by Serratia marcescens isolated from industrial effluent. Int J Curr Res Aca Rev 1: 26-32
Ray A (2012) Application of lipase in industry. Asian J Pharm Tech 2: 33-37
Sharma AK, Sharma V, Kuila A, Saxena J (2016b) Optimization of culture conditions for amylase production from locally isolated Aspergillus sp. J Biol Nat 6: 155-165
Sharma AK, Sharma V, Saxena J, Kuila A (2016a) Lipase production from a wild (LPF-5) and a mutant (HN1) strain of Aspergillus niger. Afr J Biotech 15: 2292-2300
Sharma AK, Sharma V, Saxena J, Yadav B, Alam A, Prakash A (2015a) Optimization of protease production from bacteria isolated from soil. Appl Res J 1: 388-394
Sharma AK, Sharma V, Saxena J, Yadav B, Alam A, Prakash A (2015b) Effect of culture conditions on protease production and activity of protease from soil borne fungi. Int J Sci Res Environ Sci 3: 0411-0419 Shivika S, Shamsher SK (2014) Organic solvent tolerant
lipases and applications. Sci World J 2014: 1-15
Sopuruchukwu IF, Ndidi EM, Okerentugba PO (2015) Optimization of Process Parameters for the Production of Lipase in Submerged Batch Fermentation by Fusarium sp. J Pharma Biol Sci 10: 70-78
Supakdamrongkul P, Bhumiratana A, Wiwat C (2010) Optimization of extracellular lipase production from the biocontrol fungus Nomuraea rileyi. Biocontrol Sci Technol 20: 595-604
Sztajer H, Maliszewska I, Wieczorek J (1988) Production of exogenous lipase by bacteria, fungi and actinomycetes. Enzyme Microb Technol 10: 492-497 Ulker S, Ozel A, Colak A, Karaoglu SA (2011) Isolation, production, and characterization of an extracellular lipase from Trichoderma harzianum isolated from soil. Turk J Biol 35: 543-550
Verma S and Prakash SK (2014) Isolation, identification and characterization of lipase producing microorganisms from environment. Asian J Pharm Clin Res 7: 219-222
Winkler UK and Stuckmann M (1979) Glycogen hyaluronate and some other polysaccharides greatly enhance the formation of exolipase by Serratia marcescens. J Bacteriol 138: 663-670