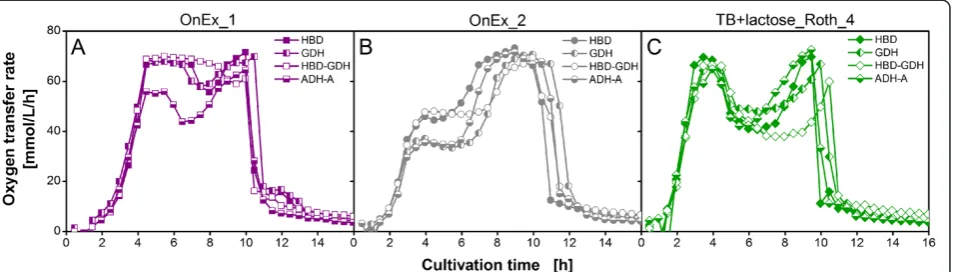
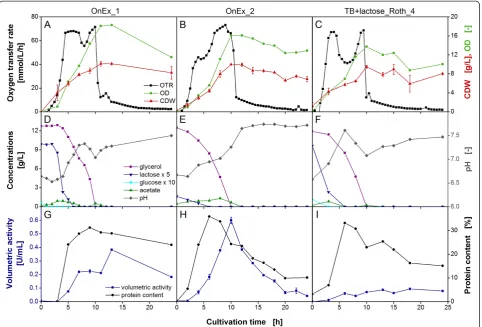
Phenotyping the quality of complex medium components by simple online-monitored shake flask experiments
Full text
Figure
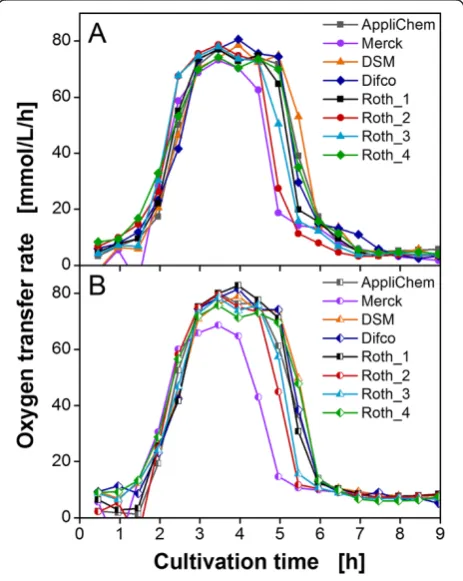
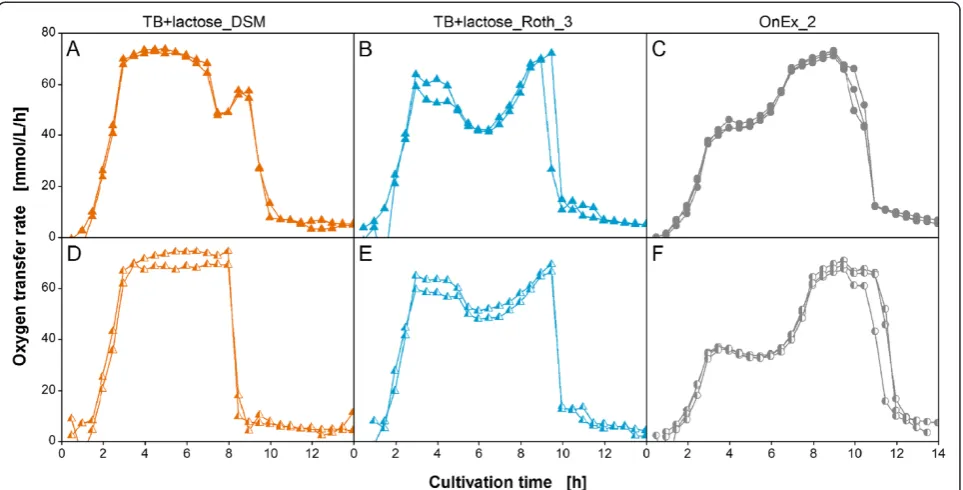
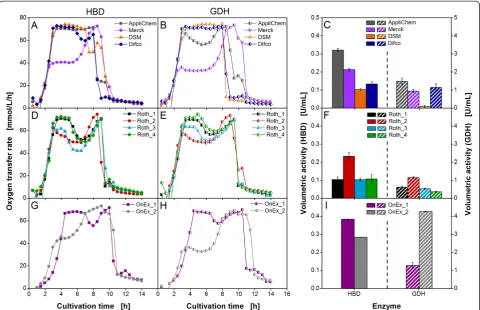
Related documents
In this present study, antidepressant activity and antinociceptive effects of escitalopram (ESC, 40 mg/kg) have been studied in forced swim test, tail suspension test, hot plate
Child sexual abuse' is often difficult to prove. The Supreme Court observed in Pennsylvania v. 3 The problems of ineffective testimony and few eye- witnesses are
Acme Glue + dynamite + standard gravity (parabolic arc); second time–parabolic arc; catapults; dynamite; Acme Outboard Motor + jim-dandy Wagon + washtub + roller skates = Gravity
Although the issues of cultural and linguistic competence and performance hardly constitute a new idea in literary writing, my discussion will be dedicated to some aspects of
Preliminarily, Molien and [Sesma] are inapposite, because the Canavins neither pleaded an independent cause of action for personal injury based upon emotional
Background: Councils of Chiropractic Education (CCE) indirectly influence patient care and safety through their role of ensuring the standards of training delivered by
It was decided that with the presence of such significant red flag signs that she should undergo advanced imaging, in this case an MRI, that revealed an underlying malignancy, which
Twenty-five percent of our respondents listed unilateral hearing loss as an indication for BAHA im- plantation, and only 17% routinely offered this treatment to children with
