RESEARCH ARTICLE
A dual function of FGF signaling in
Xenopus
left-right axis
formation
Isabelle Schneider*, Jennifer Kreis, Axel Schweickert, Martin Blum and Philipp Vick‡
ABSTRACT
Organ left-right (LR) asymmetry is a conserved vertebrate feature, which is regulated by left-sided activation of Nodal signaling. Nodal asymmetry is established by a leftward fluid-flow generated at the ciliated LR organizer (LRO). Although the role of fibroblast growth factor (FGF) signaling pathways during mesoderm development is conserved, diverging results from different model organisms suggest a non-conserved function in LR asymmetry. Here, we demonstrate that FGF is required during gastrulation in a dual function at consecutive stages ofXenopus embryonic development. In the early gastrula, FGF is necessary for LRO precursor induction, acting in parallel with FGF-mediated mesoderm induction. During late gastrulation, the FGF/Ca2+-branch is required for specification of the
flow-sensing lateral LRO cells, a function related to FGF-mediated mesoderm morphogenesis. This second function in addition requires input from the calcium channel Polycystin-2. Thus, analogous to mesoderm development, FGF activity is required in a dual role for laterality specification; namely, for generating and sensing leftward flow. Moreover, our findings in Xenopus demonstrate that FGF functions in LR development share more conserved features across vertebrate species than previously anticipated.
KEY WORDS: FGF, Pkd2, Sprouty, Left-right asymmetry, Leftward flow, LRO
INTRODUCTION
Left-right (LR) asymmetry of inner organs is present in all deuterostome lineages and depends on the left-sided activation of the evolutionarily highly conserved Nodal signaling cascade. In fish, amphibian and mammalian embryos, the symmetry-breaking event, which activates this asymmetric gene cascade in the lateral plate mesoderm (LPM), is represented by cilia-based extracellular fluid flow at the left-right organizer (LRO) of the early neurula embryo (Dasgupta and Amack, 2016; Yoshiba and Hamada, 2014). The flow-generating LRO epithelium consists of mono-ciliated cells of mesodermal fate, which are transiently embedded in the endodermal endothelium of the forming archenteron at the posterior end of the notochord (Blum et al., 2007). In amphibians, it is of triangular shape and termed the gastrocoel roof plate (GRP; Blum et al., 2009b). TheXenopusGRP is subdivided into a medial flow-generating part, with posteriorly localized motile cilia, and lateral cells with non-polarized and immotile cilia, which presumably
sense flow. The former cells will later integrate into notochord and hypochord, whereas the latter will contribute to somites (Boskovski et al., 2013; Shook et al., 2004). Somitic GRP cells are characterized by co-expression ofnodaland the Nodal inhibitor genedand5. Flow is thought to downregulate dand5, de-repress Nodal and induce transfer of an asymmetric signal to the left LPM (Schweickert et al., 2010). The GRP derives from the superficial mesoderm (SM; Shook et al., 2004) of the early gastrula, which represents the outer-most layer of the Spemann organizer. The SM is marked by the expression of the transcription factor geneforkhead box J1(foxj1; Blum et al., 2014; Stubbs et al., 2008), which induces motile ciliogenesis, and the Nodal-related growth factor gene nodal3 (nodal homolog 3; previously known asxnr3; Smith et al., 1995; Beyer et al., 2012).
A number of signaling pathways have been implicated in LR development, although conserved functions have only been described for a few, such as Nodal (Namigai et al., 2014). Conflicting results have been obtained for FGF signaling. In chick and rabbit embryos, the ligand Fgf8 seems to act as a right determinant, repressing asymmetric gene expression in the right LPM (Boettger et al., 1999; Feistel and Blum, 2008; Fischer et al., 2002). In the mouse, in contrast, Fgf8 acts as a left determinant. Here, FGF signaling is required both fornodalgene expression in the sensory crown cells of the LRO at about the 2-somite stage, and for asymmetric gene expression in the LPM later on (Meyers and Martin, 1999; Oki et al., 2010). In addition, FGF signaling has been implicated as a regulator of vesicle release at the LRO; these vesicles transfer through flow to the left side, where they activate Nodal signaling (Tanaka et al., 2005). In teleost fish, FGF signals were shown to be required for symmetry breakage upstream of leftward flow. Loss of FGF signaling resulted in reduction or loss of LRO cilia in specimens harboring five to ten pairs of somites, and, consequently, LR defects later on (Hong and Dawid, 2009; Neugebauer et al., 2009; Yamauchi et al., 2009). Reduced ciliary lengths were reported in theXenopusGRP, but GRP morphogenesis, foxj1, flow and Nodal cascade genes have not been analyzed (Neugebauer et al., 2009). Thus, the role of FGF signaling seems to vary in the different model organisms analyzed to date.
Here, we studied the role of FGF signaling in LR axis formation in Xenopus. The frog is accessible to manipulation and analysis at all stages of LR development, from SM specification to GRP morphogenesis, leftward flow, Nodal cascade induction and organ morphogenesis (Blum et al., 2014, 2009a). Through pharmacological and molecular manipulation of Fgfr1 signaling, we identified a dual role of FGF signaling. During early gastrulation, FGF signaling was necessary for SM specification upstream offoxj1 but downstream ofnodal3. During late gastrulation, a second, cilia-independent function was involved in morphogenesis of the lateral flow sensor of the LRO, independently offoxj1. Our data suggest that this late phenotype was caused by interfering with FGF/Ca2+ signaling, which additionally required input from the calcium channel Polycystin-2. Our work reconciles conflicting findings in Received 13 November 2018; Accepted 18 April 2019
Institute of Zoology, University of Hohenheim, 70593 Stuttgart, Germany. *Present address: Molecular Cardiology, University Hospital Ulm, 89081 Ulm, Germany.
‡Author for correspondence ( [email protected])
P.V., 0000-0002-1430-4561
DEVEL
O
model organisms and demonstrate a conserved role of FGF signaling in LR axis formation, in much the same dual way as has been shown for mesoderm induction and morphogenesis.
RESULTS
FGF signaling during gastrulation is necessary for LR asymmetry
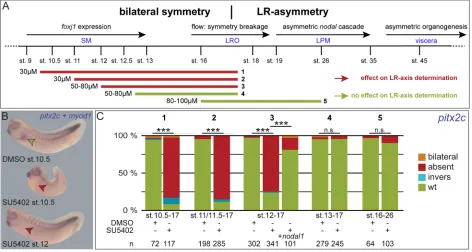
In order to determine the critical time frame of FGF signaling in XenopusLR development, we treated embryos of defined stages with SU5402, an inhibitor of the tyrosine kinase activity of Fgfr (Fig. 1A). SU5402 incubation at late blastula or early gastrula stages (until stage 10.5) impaired gastrulation movements leading to blastopore closure defects, reduced somitic expression ofmyogenic differentiation 1(myod1), and a shortened anterior-posterior axis, as previously described for inhibition of FGF signaling (Fig. 1B; Amaya et al., 1991). Concomitantly, FGF-dependent mesodermal marker genes were strongly (myogenic factor 5,myf5; Fig. S1A-C) or moderately (myf5;myod1;brachyury/T-box transcription factor T,tbxt; Fig. S1D-L) reduced at late gastrulation when treated from late blastula (stage 9) or early gastrula (stage 10) onwards, respectively. Mesodermal defects at tadpole stages could be minimized by treatment at mid gastrulation (stage 11/11.5), and circumvented when treated from late gastrula stages onwards (st.12; Fig. 1B; data not shown). In all cases, however, i.e. from early, mid or late gastrulation onwards, FGF inhibition resulted in absence of pitx2cexpression in the left LPM (Fig. 1; Schweickert et al., 2000). Treatments at later stages did not impact on laterality (Fig. 1C). Inhibition of FGF signaling did also not impair the competence of LPM tissue to respond to Nodal, as injection of a nodal1 DNA construct into the LPM of SU5402-treated embryos rescuedpitx2c expression (Fig. 1C). These experiments revealed a sensitive time
window for FGF signaling in LR axis development from late blastula until late gastrulation, i.e. before establishment of leftward flow and, thus, upstream ofnodal1activation in the left LPM.
A first FGF signal is required forfoxj1induction during early gastrulation
[image:2.612.71.543.415.667.2]Loss of asymmetricpitx2cexpression could be caused by lack of symmetry breaking due to impaired leftward flow. Gastrulation defects, however, prevented the analysis of leftward flow in embryos treated before stage 10.5 (Fig. 1B). We therefore decided to analyze the specification of the precursor tissue from which the LRO derives, namely the SM, using mRNA expression of the known marker genesnodal3,foxj1andwnt11b. This reasoning was further supported by our recent demonstration that knockdown of the Fgfr1 ligandnodal3in the SM inhibitedfoxj1induction (Vick et al., 2018; Yokota et al., 2003). Early SM-specific expression of nodal3 was unaltered when treated from late blastula onwards, confirming its dependence on canonical Wnt but independence of FGF signaling pathways, as demonstrated previously (Fig. 2A,B,G; Fletcher and Harland, 2008; Smith et al., 1995). In agreement with this notion, embryos treated with SU5402 at early gastrula (stage 10.5) showed a significant reduction of foxj1 and wnt11b in the SM, confirming a role of Nodal3/FGF in setting-up a functional GRP (Fig. 2C-G). Endodermal expression ofwnt11bwas unaffected by SU5402 treatment, underscoring the FGF specificity of this effect (2E-F′). Injection of a dominant-negative version of Fgfr1 (dnfgfr1; Amaya et al., 1991) mimicked SU5402 treatment and resulted in a significant loss of foxj1 expression as well (Fig. S2A,B,D). Together, these experiments demonstrated that FGF signaling was required for induction of gastrulation movements and in parallel for induction of foxj1 and wnt11b
Fig. 1. FGF signaling is required for LR axis development during gastrulation.(A) Timeline of events relevant for LR axis development (top), and SU5402 incubation periods (bottom; marked as 1-5). (B) Representative specimens encountered following co-staining forpitx2candmyod1mRNA expression. Top: DMSO-treated embryo; WT morphology and expression patterns (left-sided expression ofpitx2c, marked by green arrowhead, andmyod1in somites). Middle: specimen treated with SU5402 at early gastrulation (stage 10.5) displaying lack ofpitx2cexpression (red arrowhead). Bottom: specimen treated with SU5402 at late gastrula (stage 12). Note the strongly decreased expression ofmyod1upon early, but not late, SU5402 treatments. (C) Quantification ofpitx2cexpression patterns in specimens treated during indicated time periods (1-5). n.s., not significant; st., stage; wt, wild type; LR, left-right; LRO, left-right organizer; LPM, lateral plate mesoderm; SM, superficial mesoderm.n, number of analyzed embryos.
DEVEL
O
expression in the SM. Because earlier nodal3 expression was unaffected by SU5402 treatments, the initial SM identity seemed not to be perturbed, but an early FGF signal was required at the beginning of gastrulation for further SM specification.
A second FGF signal regulates Nodal cascade induction independent offoxj1and motile cilia
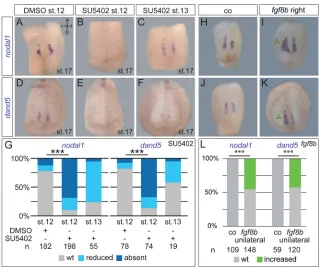
Interestingly, SU5402 incubation at late gastrula stages had no impact onfoxj1, in agreement with the robust expression offoxj1at these stages (Fig. 2H,I; cf. Beyer et al., 2012; Stubbs et al., 2008). Injection of low doses of dnfgfr1 did not cause a significant loss of foxj1 expression either, nor did it block gastrulation movements, although pitx2c expression in the left LPM was partially lost (Fig. S2C-E). At neurula stages, expression of the Foxj1 target gene tektin2 (tekt2) in the GRP was apparently unaffected by late-gastrula-stage SU5402 incubations. The tekt2 expression domain, however, appeared narrower than that of control embryos (Fig. 2J,K). We therefore analyzed cilia formation and functionality by scanning electron microscopy (SEM) and flow analysis, to test for effects downstream of ciliary gene activation. SEM analysis did not reveal significant differences in cilia length, GRP ciliation or posterior polarization of flow-generating motile cilia between treated and control specimens (Fig. S3A-E). Low dose dnfgfr1injections did not alter ciliary length either (Fig. S3F). Cilia motility and leftward flow were also unaffected, as demonstrated by flow analysis of DMSO- and SU56402-treated GRP explants from stage 17/18 embryos (Fig. S3G-I; Schweickert et al., 2007). Together, these data hint at two different modes of action of FGF signaling in Xenopus LR axis development: a cilia- and foxj1-dependent function at early gastrulation, and a later, cilia-independent mode of action downstream of flow and upstream of nodal1expression in the LPM.
Late FGF signaling is necessary and sufficient for LRO flow sensor formation
[image:3.612.46.351.53.373.2]Next, we investigated whether signaling downstream of flow was impaired in embryos treated at late gastrula stages, i.e. whether the left-asymmetric flow signal was perceived and transferred to the LPM. In wild-type (WT) embryos, polarized motile cilia at the center of the GRP generate flow, which is perceived by immotile and central cilia on lateral GRP cells (Boskovski et al., 2013; Schweickert et al., 2007; Shook et al., 2004). At the molecular level, the GRP in its entirety can be highlighted by expression of ciliary marker genes, such astekt2(Stubbs et al., 2008) ordynein axonemal heavy chain 9(dnah9; Schweickert et al., 2007), or, indirectly, by the lack of endodermal marker gene expression, such as alpha-2-macroglobulin-like 1(a2ml1, also known aspanza; Pineda-Salgado et al., 2005; Fig. S4A-C). Sensory lateral GRP cells are characterized by the co-expression of both nodal1 and its inhibitordand5(Fig. S4D; Schweickert et al., 2010; Vonica and Brivanlou, 2007), whereas flow-generating central GRP cells expresssonic hedgehog (shh; Fig. S4E). The combined analysis of shh(central GRP) and a2ml1 (endoderm) thus highlights the lateral, sensory part of the GRP by an absence of staining (Fig. S4F). SU5402 incubation at stage 12 resulted in lack of bothnodal1and dand5 expression (Fig. 3A,B,D,E,G), but effects were greatly reduced upon later incubations from stage 13 onwards (Fig. 3C,F, G). To confirm that lateral, sensory GRP cells were specifically absent upon late SU5402 treatment, we analyzed embryos from the same batches in parallel for expression of further marker genes. Whereas untreated embryos showed the signal-free area between the shh and a2ml1 expression domains, in addition to WT nodal1 expression (which together unequivocally mark the lateral GRP cells), about 50% of specimens treated at stage 12 lacked both nodal1-positive cells and theshh/a2ml1-negative region between Fig. 2. Inhibition of FGF signaling at early gastrulation acts upstream offoxj1.(A,B) Expression patterns of the SM marker genenodal3. Expression was unaltered following SU5402 treatment from stage 9 to 10.25 (N=3).
(C,D) Representativefoxj1expression patterns at stage 11.5 in embryos treated with DMSO (C) or SU5402 up to gastrula stage 10.5 (D) demonstrating downregulation of the SM marker genefoxj1in embryos treated with SU5402 (N=3).
(E,F) Representativewnt11bexpression patterns at stage 11 in embryos treated with DMSO (E) or SU5402 starting before stage 10.5 (F) demonstrating downregulation ofwnt11b
in the SM of embryos treated with SU5402 (N=2). (E′,F′) Mid-sagittal view of bisected specimens shown in E and F. (G) Quantification of results following early treatments. (H-K) Representative examples of expression patterns of
foxj1(H,I), andtekt2(J,K) in embryos treated with DMSO (H,J) or 80 µM SU5402 from mid-gastrula stages onward (I,K) demonstrating unaltered expression offoxj1(in the SM) and
tekt2(in the GRP) of embryos treated with SU5402. Embryos in J and K show ventral views of dorsal explants displaying archenteron roof and embedded GRP. Gray arrowheads highlight WT expression, blue arrowheads highlight reduced expression. a, anterior; l, left; p, posterior; r, right; st., stage; wt, wild type.N, number of experiments;n, number of embryos analyzed.
DEVEL
O
central GRP and endoderm. Again, treatments from stage 13 onwards impacted only slightly (in 5%) on the presence of the sensory GRP (Fig. S4G-L). The lack of somitic GRP cells was not due to interfering with mesodermal maintenance during neurulation, as only early but not late treatments resulted in reduction of expression of mesodermal marker genes brachyury (tbxt) andmyod1(Fig. S4M-R). Confirming the specificity of this effect on the sensory GRP, low-dosednfgfr1injections in a sided manner resulted in a significant reduction or loss ofnodal1 and dand5on the injected side (Fig. S4S-X). In line with this notion, activation of FGF signaling by injection of the ligandfgf8binto the lateral GRP was sufficient to increase the expression of bothnodal1 and dand5 in a highly significant proportion of specimens (Fig. 3H-L). Overexpression was achieved through injections of a DNA construct, which only gets transcribed after activation of the zygotic genome at late blastula stages, a procedure chosen to prevent earlier FGF-induced developmental defects. Together, these experiments pointed to a function of FGF signaling during late gastrulation to promote the lateral, somitic fate of the GRP.
As bothnodal1anddand5expression were lost, we wondered if FGF inhibition prevented activation of these genes, or whether the late FGF signal was required for formation of sensory cells. These cells in due course detach from the gastrocoel roof and integrate into the presomitic mesoderm (Shook et al., 2004). Their somitic fate is marked already in the GRP by expression ofmyod1(Fig. 4A,B,E; Schweickert et al., 2010). This contribution ofmyod1-positive cells to the GRP was essentially lost in specimens treated with SU5402 from stage 12 onwards (Fig. 4C,E), to a lesser degree when incubation started at stage 13 (Fig. 4D,E). This loss was quantified by assessing the widths of GRP subpopulations in transversal sections at the level of the posterior aspect of the GRP. The relative widths of the central GRP were not different between control and SU5402-treated specimens. The mean widths of the lateral, somitic part of the GRP, however, were reduced to less than 20% of the values of control specimens upon late FGF inhibition (Fig. 4E). Treatments from stage 13 onwards showed some reductions in
widths as well (Fig. 4D′,E), although LR development was not affected. Low doses ofdnfgfr1injections likewise resulted in a lack ofmyod1-positive lateral GRP cells on the injected side (Fig. 4F), demonstrating specificity of effects.
Next, we wondered whether proliferation and/or apoptosis were involved in FGF-mediated GRP patterning, as the FGF pathway impacts on both processes (Schlessinger, 2000). The proliferation rate in the GRP was assessed in explants at two stages (13/14 and 16), using a phospho-histone3-specific antibody. No differences were recorded between untreated samples and specimens incubated with SU5402 from stage 11.5 onwards (Fig. S5A-C; Saka and Smith, 2001). Apoptosis was unaffected by FGF inhibition as well, as demonstrated by terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) staining of control and SU5402-treated embryos at stages 13/14 or stage 16. Here, we found apoptosis in the neuroectoderm, as previously reported (Yeo and Gautier, 2004), but never at the GRP (Fig. S5D,E). In agreement with the notion that apoptosis was not involved in FGF-mediated specification of lateral GRP cells, injection of the anti-apoptotic B-cell CLL/lymphoma (bcl-XL) DNA construct, which inhibits chemically induced apoptosis (Johnston et al., 2005), into the lineage of the lateral GRP was unable to rescue SU5402-induced loss of nodal1/dand5-positive GRP cells (Fig. S5F-K). Thus, neither apoptosis nor altered proliferation within the GRP are causative for the observed lack of somitic GRP cells upon late-gastrula-stage loss of FGF signaling.
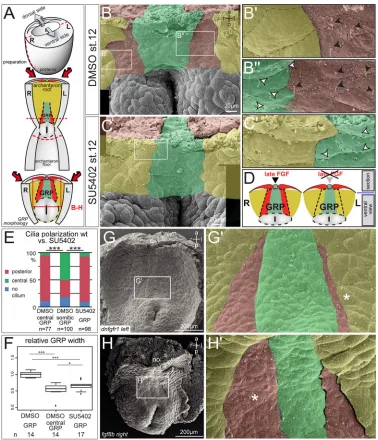
[image:4.612.47.368.55.324.2]In a last set of experiments, we analyzed the morphological distortions of GRP morphogenesis and patterning following FGF inhibition by SEM. Using GRP explants from DMSO- or SU5402-treated embryos (Fig. 5A), three traits were analyzed: cilia polarization, tissue morphology and vesicle abundance. Ciliated GRP cells are easily distinguishable from the much larger non-ciliated lateral endodermal cells (LECs; Fig. 5B). As previously reported (Schweickert et al., 2010), cilia at the central part of the GRP are posteriorly polarized, whereas lateral cells harbor central cilia (Fig. 5B′,B″). Cilia polarization was facilitated by viewing SEM preparations of GRP explants at an angle (cf. Fig. S6A).
Fig. 3. FGF signaling regulates somitic GRP specification from mid to late gastrulation.
(A-F) Analysis ofnodal1(A-C) anddand5(D-F) mRNA expression in stage 17 embryos treated with DMSO (A,D) or 50 µM SU5402 (B,C,E,F) from stage 12 (A,B,D,E) or 13 (C,F) onwards. Lateral GRP-specific expression patterns shown in ventral view of dorsal explants. (G) Quantification of results (N=3). (H-K) Analysis of
nodal1(H,I) anddand5(J,K) in stage 17 control specimens (H,J) or embryos unilaterally injected with
fgf8bDNA (5-8 pg; I,K), specifically targeting the right expression domain ofnodal1/dand5. Injected embryos showed a lateral increase of expression domains on the targeted side. (L) Quantification of results (N=3). a, anterior; co, control; l, left; n.s., not significant; p, posterior; r, right; st., stage; wt, wild type.N, number of experiments;nare numbers of embryos analyzed.
DEVEL
O
SU5402-treated GRPs lacked lateral GRP cells harboring non-polarized cilia (Fig. 5C,D; Fig. S6B), such that GRP cells with polarized cilia were directly bordered by LECs (Fig. 5C′,D). Quantification of cilia polarization revealed the same percentage of posteriorized cilia in central GRPs of DMSO-treated and whole GRPs of SU5402-treated specimens. The fraction of about 50% non-polarized cilia in lateral GRP cells of WT specimens (Fig. 5E) was lost following SU5402 treatment. With respect to overall tissue morphology, there is a clear separation between central and lateral parts of WT GRPs, enabling a direct assessment of tissue widths (Fig. 5B; Fig. S6C): cells align at the border between central and lateral GRP areas, giving rise to an almost straight line separating the two cell populations, as previously observed by Shook et al. (2004). SU5402-treated GRPs were found to be narrower, encompassing just the widths of WT central GRPs (Fig. 5F; Fig. S6B). Finally, higher magnification of SEM photographs revealed the presence of small vesicles across the apical surface of all GRP cells (Fig. S6C). Upon close inspection, we noticed that vesicles were enriched on lateral GRP cells, which offered itself for direct quantification. GRPs from SU5402-treated specimens showed lower vesicle abundance compared with untreated specimens (Fig. S6D), in line with a loss of lateral GRP cells. Of note, vesicle abundance did not differ between left and right somitic cells in WT GRP cells (Fig. S6E). SEM analysis ofdnfgfr1-injected embryos revealed a strong reduction of lateral GRP cells on the injected side as well, confirming the specificity of SU5402 treatment (Fig. 5G). Finally,
unilateral injection offgf8bresulted in a marked increase of lateral GRP widths (Fig. 5H). Together, these analyses demonstrated that Fgfr-mediated signaling is required for the morphogenesis of lateral, flow-sensing GRP cells during late gastrulation.
Sprouty-induced inhibition of FGF/Ca2+signaling blocks the GRP flow sensor
A dual role of FGF signaling has been previously described in the context of mesoderm induction and subsequent morphogenesis (Nutt et al., 2001; Sivak et al., 2005). Specifically, MAPK-dependent FGF signaling is necessary during early/mid gastrulation for mesoderm induction and maintenance. At mid-gastrulation, the PKC-dependent FGF/Ca2+ branch is required for morphogenetic gastrulation movements, which, however, does not affect mesoderm induction. The FGF antagonistsproutyis expressed during early gastrulation; overexpression does not impact on MAPK-mediated mesoderm induction, but blocks FGF/Ca2+-activation during late gastrulation, i.e. specifically regulates FGF-mediated morphogenesis (Nutt et al., 2001; Sivak et al., 2005). The dual role of FGF signaling in LR axis formation made us wonder whether the FGF/Ca2+branch was likewise involved in the FGF-mediated specification of the sensory GRP.
[image:5.612.140.474.54.330.2]In order to test this possibility, we injectedspry1mRNA to target the SM and lateral GRP cells specifically. Sprouty overexpression did not affectfoxj1expression in the SM, nor subsequent activation of its target tekt2 in the GRP (Fig. 6A-F). Mesoderm induction and mesodermal marker gene expression (tbxt,myod1) at gastrula and Fig. 4. FGF inhibition during late gastrulation blocksmyod1expression in the lateral GRP.(A) Schematic of explant preparation formyod1analyses. Neurula-stage embryos were bisected transversally and the posterior part bisected into dorsal and ventral halves (top). Wild-typemyod1-expression in the somitic GRP is highlighted in blue. Dorsal halves were analyzed formyod1expression (B-F′;N=4). (B-D) ISH analysis ofmyod1expression in embryos treated with DMSO (B) or 50 µM SU5402 (C,D) from stage 12 (B,C) or stage 13 (D) onwards. Ventral views of representative dorsal explants. Position of GRP is indicated by the curved dashed line. (B′-D′) Histological analysis ofmyod1in transversal sections, as indicated by dashed white lines in B-D. Blue bars mark widths ofmyod1-positive lateral GRP, which was strongly (C′; stage 12) or slightly (D′; stage 13) reduced by SU5402 treatment. Black bars indicate unaltered central part of GRP. (E) Quantification of relative widths of central (top) and lateral somitic (bottom) GRP in DMSO- and SU5402-treated embryos, as described in the Materials and methods section. (F) Analysis ofmyod1in stage 17 dorsal explants (ventral view) injected unilaterally withdnfgfr1mRNA (8-12 pg) targeted to the left lateral GRP. (F′) Histological analysis in transversal section (at the level indicated in F). Contribution of somiticmyod1-positve cells to the GRP (highlighted by dotted line on uninjected side) was lost on the injected side (black arrowheads). a, anterior; ar, archenteron; ec, ectoderm; en, endoderm; l, left; no, notochord; n.s., not significant; p, posterior; psm, presomitic mesoderm; r, right; s, somite; st., stage; wt, wild type.N, number of experiments;nare numbers of embryos analyzed.
DEVEL
O
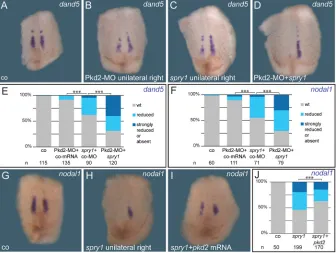
neurula stages were unaffected, as previously described (Fig. S7A-F; Sivak et al., 2005). However, the width of thetekt2expression domain was reduced in about 45% of injected specimens followingspry1 injections (compare Fig. 6D and E). Concomitantly, expression of nodal1anddand5 were lost on the injected side in about 50% of unilaterally injected specimens (Fig. 6G-L). The loss of the lateral GRP cells upon unilateralspry1injection was confirmed by SEM analysis, which revealed about 75% of embryos with loss (Fig. S7G,I) or strong reduction of lateral GRP cell populations (Fig. S7H,I).
Together, these results unveil an inhibitory effect of Sprouty proteins on the morphogenesis of the LRO flow sensor, i.e. somitic GRP cell formation during late gastrulation, suggesting a requirement of the FGF/Ca2+pathway in this process.
FGF/Ca2+and
[image:6.612.117.495.55.495.2]pkd2synergize in LRO sensor formation Like FGF,pkd2is required in a dual way during symmetry breakage inXenopus. In a first stage, it cooperates with the FGF ligandnodal3 to inducefoxj1in the SM. Later, it is required for somitic GRP cells Fig. 5. FGF inhibition during late gastrulation impairs somitic GRP morphogenesis.(A) Schematic of explant preparation for morphological analysis. Neurula-stage embryos were bisected transversally to isolate posterior-dorsal halves, which were bisected transversally to enable visualization of GRPs by SEM. Lateral somitic GRP and somites are indicated in red, central GRP and notochord in green, and endoderm in yellow. (B-D) SEM of stage 17 dorsal explants (ventral view) treated with DMSO (B) or 60 µM SU5402 (C) from stage 12 onwards, highlighting GRP subpopulations and flanking LECs. Higher magnification of DMSO- (B′,B″) and SU5402-treated (C′) specimens reveal the presence (B′,B″) or absence (C′) of lateral, somitic GRP areas, which are characterized by non-polarized central cilia (indicated by black arrowheads). Cells are outlined by dashed lines and colored in red (lateral somitic GRP), green (central GRP) and yellow (LECs). White arrowheads indicate polarized cilia of central GRP cells, which are localized to the posterior part of the cell. Note the lack of non-polarized, somitic GRP after late SU5402 treatments, as illustrated in D. (E) Quantification of cilia polarization (N=2). (F) Quantification of GRP widths (mean widths of DMSO-treated GRPs set to 1.0;N=2). (G,H) Representative SEM photographs of stage 17 dorsal explants (ventral view) injected unilaterally on the left withdnfgfr1mRNA (G; 8-12 pg;N=3;n=25) or withfgf8bDNA on the right (H; 5 pg). (G′,H′) Higher magnifications of the boxed areas in G,H reveal reduction (G′;dnfgfr1-injections on left indicated by asterisk) or expansion (H′;fgf8b-injection on right indicated by asterisk) of lateral somitic GRP areas. a, anterior; l, left; no, notochord; n.s., not significant; p, posterior; r, right; s, somite; st., stage; wt, wild type.N, number of experiments;n, number of embryos analyzed.
DEVEL
O
and initiation ofnodal1 and dand5(Vick et al., 2018). As pkd2 encodes the calcium channel Polycystin-2 (Busch et al., 2017; Koulen et al., 2002), we reasoned that it may interact synergistically with the FGF/Ca2+ pathway. To investigate this possibility, we injected a characterizedpkd2antisense morpholino oligomer (MO; Tran et al., 2010) to target the nodal1/dand5-expressing somitic GRP cells. The MO prevented expression of both genes specifically on the injected side, as described (Fig. S8A-D; cf. Vick et al., 2018). To test for genetic interaction, we lowered bothpkd2-MO dose and concentration of spry1 mRNA such that nodal1 and dand5 expression were only mildly affected on the injected side (Fig. 7A-F). Co-injections of sub-phenotypic doses, however,
[image:7.612.50.384.55.319.2]resulted in a highly significant reduction of both genes (Fig. 7D-F). These experiments indicated thatpkd2loss- andsprouty gain-of-function impact on the same process, namely somitic GRP-induction. As Sprouty presumably blocks the FGF/Ca2+ pathway upstream of endoplasmic reticulum-mediated intracellular Ca2+ -release (Akbulut et al., 2010; Nutt et al., 2001),pkd2might be able to rescue the spry1-induced loss of sensory GRP cells. Indeed, co-injection ofspry1andpkd2partially restorednodal1expression in lateral GRP cells (Fig. 7G-J). These final experiments suggested a synergy of pkd2 and the FGF/Ca2+ branch of FGF signaling in inducing the somitic GRP, i.e. for specification of the LRO flow sensor.
Fig. 6. Sprouty1-mediated FGF inhibition blocks specification of somitic GRP.(A-L) Marker gene analyses of control andspry1-injected embryos at stage 11 (A-C), and dorsal explants at stage 17 (D-L;N=3). (C,F,I,L) Quantification of results. (A-C) Wild-typefoxj1expression in control specimens (A,C) and embryos bilaterally injected withspry1mRNA (B,C;N=3). (D-F) Narrowedtekt2
expression inspry1-injected specimens (E; light blue arrowheads), compared with control embryos (D; gray arrowheads;N=3). (G-L)nodal1(G-I) and
dand5(J-L) were reduced or absent on thespry -injected sides following unilateral mRNA injection (dark blue arrowheads;N=3).spry1mRNA was injected at concentrations ranging from 320 to 800 pg. (A,B) Vegetal views, dorsal side up. (D,E,G,H,J,K) Ventral views of dorsal explants. bi, bilateral; n.s., not significant; wt, wild type.
N, number of experiments;n, numbers of embryos analyzed.
Fig. 7. spry1andpkd2cooperate in somitic GRP specification.(A-F) Epistatic analysis of FGF/Ca2+and Polycystin-2 function by
simultaneous overexpression ofspry1and knockdown ofpkd2, using sub-phenotypic doses. ISH analysis ofdand5(A-E;N=3) ornodal1
(F;N=3) in stage 17 embryos showed wild-type expression in control specimens (A,E,F). No or minor reductions were observed following unilateral injection of low doses ofspry1mRNA (50-160 pg) in combination with a control-MO (4 ng; C,E,F), or by injection of a Pkd2-MO (4 ng) in combination with a control mRNA (50-160 pg; B,E,F). Combinations ofspry1mRNA (50-160 pg) and Pkd2-MO (4 ng) resulted in a high proportion (>50%) of embryos with reduced or absent gene expression levels (D-F). GFP mRNA was used as control mRNA, control (co) bar represents uninjected specimens. (G-J) Rescue ofnodal1
expression (G) inspry1-injected specimens (H; 320-800 pg) by co-injection ofpkd2mRNA (I). (J) Quantification of results (N=4). (G-I). Ventral views of dorsal explants. wt, wild type.N, number of experiments;n, numbers of embryos analyzed.
DEVEL
O
[image:7.612.50.386.484.737.2]DISCUSSION
Our analysis of FGF signaling during left-right axis formation in Xenopusdemonstrated a dual role, namely for specification of the LRO precursor, the SM, during early gastrulation, and a later, independent role for sensory cell formation in the LRO itself. These findings reconcile conflicting data reported previously from zebrafish and mouse. LRO ciliogenesis and earlier foxj1 expression were reduced or lost in zebrafish between 90% epiboly and the 10-somite stage following SU5402 treatment or knockdown of fgfr1or fgf8 (Caron et al., 2012; Hong and Dawid, 2009; Neugebauer et al., 2009). In contrast, data reported from a hypomorphic Fgf8 mouse line suggested that lateral, sensory LRO cells might be missing, based on the absence of Nodal expression at the LRO, which is an evolutionarily conserved hallmark of ciliated LROs (Blum et al., 2007, 2009b; Meyers and Martin, 1999). Strikingly, manipulation of FGF signaling during late gastrulation (late-bud to 3-somites) using FGF inhibitors also resulted in loss ofNodalexpression at the mouse LRO (Oki et al., 2010). Whether or not this function was mediated through the FGF/Ca2+pathway, as suggested here, was not addressed in either of these studies, nor a possible morphogenetic role in sensory cell formation, a concept that emerged only later (McGrath et al., 2003; Tabin and Vogan, 2003). For future analyses, it may be worthwhile to re-evaluate LRO specification (Foxj1 expression) and ciliation in mouse embryos with reduced FGF signaling (in the hypomorphic Fgf8line or following early SU5402 treatment), in order to analyze whether the first phase of FGF-dependent laterality formation, i.e. LRO precursor formation, is conserved in mammalian embryos as well. Lateral, flow-sensing cells in zebrafish are less well defined (Ferreira et al., 2017). MO-mediated knockdown offgf8resulted in loss ofdand5at the LRO as well (Hong and Dawid, 2009). However, this loss-of-function scenario does not allow us to distinguish between an early inhibition of FGF-dependent LRO specification or a later FGF requirement for flow sensor morphogenesis.
Together, the data on FGF function in cilia-dependent LR axis formation in the vertebrates strongly suggest an evolutionarily conserved dual role during early and late gastrulation. Interestingly, this dual function parallels the known role of FGF signaling during Xenopusmesoderm induction ( phase 1 during early gastrulation; Amaya et al., 1993; Schulte-Merker and Smith, 1995) and morphogenesis ( phase 2 during late gastrulation; Dorey and Amaya, 2010; Pownall and Isaacs, 2010). Both processes are intimately linked through the timing of events. Morphogenetic gastrulation behavior in phase 2 requires calcium, and is mediated through the FGF/Ca2+ pathway as well (Sivak et al., 2005; Wallingford et al., 2001; Wang and Steinbeisser, 2009). In the lateral GRP ofXenopus, Polycystin-2 might cooperate with FGF/ Ca2+downstream of inositol trisphosphate receptor activation at the level of intracellular calcium release, a positive interaction that has been demonstrated inXenopusoocytes (Berridge et al., 2000; Dorey and Amaya, 2010; Li et al., 2005). This notion is supported by thapsigargin-mediated interference with calcium fluxes, which also resulted in lack of nodal expression in mouse crown cells and Xenopuslateral LRO cells (Takao et al., 2013; Thastrup et al., 1990; Vick et al., 2018).
Although not considered mesodermal at the stage of emergence and function, cells at theXenopusSM (i.e. superficial cells of the organizer) and LRO (embedded in the archenteron) are fated to become mesodermal: they integrate into the notochord (flow-generating central LRO cells) and somites (flow-sensing lateral LRO cells) once the LRO submerges under the endodermal cells, which migrate medially to close the gap in the posterior dorsal archenteron during neurulation (Shook et al., 2004). Thus, SM
specification may be considered as one aspect of mesoderm induction, and LRO formation as a morphogenetic process during early mesoderm differentiation. In that sense, LRO cells take a unique detour to assume their final mesodermal fate, starting out as superficial cells overlying the organizer and dorsal mesoderm at the beginning of gastrulation, and lining the dorsal posterior archenteron later on (Blum et al., 2014). Interestingly, pkd2 is involved at both stages inXenopus(Vick et al., 2018; data presented here). It remains to be seen to what extent this parallel holds, i.e. which other known molecules previously involved in either mesodermal process participate in early LR axis formation as well. Brachyury mutant mouse lines and zebrafishno tailmorphants support this reasoning, as do our unpublished results on the role of brachyuryduring LR development inXenopus(Sabrina Kurz and M.B., unpublished; Amack and Yost, 2004; King et al., 1998). In addition, mechanical strain has been recently shown to constitute a decisive physical force in LRO morphogenesis and function, namely cilia polarization and motility (Blum and Ott, 2018; Chien et al., 2018). Remarkably, strain is only effective when foxj1 is present; therefore, it would be interesting to analyze whether or not FGF signaling is a prerequisite of strain-mediated LRO morphogenesis and function.
There is but one presumed FGF function in LR axis formation that remains unresolved in an evolutionary context: FGF-mediated vesicle release at the mouse LRO (Tanaka et al., 2005). This is more than just a cursory inconsistency, as it impacts on our conceptional understanding and perception of cilia-mediated symmetry breaking in general. To date, two models of symmetry breaking, though not necessarily mutually exclusive, co-exist: (1) flow-sensing itself by immotile lateral sensory LRO cells, which through Polycystin-2 transmits a calcium signal into the cells (two-cilia model; McGrath et al., 2003; Tabin and Vogan, 2003), and (2) the morphogen model, in which a secreted factor (Nonaka et al., 1998; Okada et al., 1999), released from LRO cells, transfers through flow to the left side, where it initiates asymmetric signaling. In one variant of the morphogen model, the flow-transported factor is localized in vesicles (i.e. exosomes or exocytic vesicles), which are released in an FGF-dependent manner from the LRO. This variant has regained attention recently, following the proposal that sensory cilia do not qualify as calcium-responsive mechanosensors at the mouse LRO (Delling et al., 2016). This proposal contrasts with experimental and genetic evidence in the mouse, which demonstrated (1) that artificial flow was able to rescue lack of cilia motility as well as to revert laterality upon inversion of flow directionality (Nonaka et al., 2002); and (2) that Polycystin-2 is required on lateral LRO cells for flow sensing (McGrath et al., 2003; Yoshiba et al., 2012). The case, therefore, seems undecided on this point. In particular, it remains to be seen at what stage vesicles or a secreted morphogen arise at the LRO, and how they transfer and fuse with cells or cilia at the left margin of the LRO. In our experiments, application of SU5402 at stages following LRO formation, i.e. at time points when such vesicles could emerge for the first time, did not impact on LR axis formation. Our data thus suggest that the reported role of FGF in releasing vesicles at the mouse LRO is likely not conserved in the frog. In addition, the vesicles observed during our SEM analyses did not accumulate on the left side, i.e. at the left lateral GRP cells. Their distribution therefore seemed unaffected by flow (cf. Fig. S6).
In conclusion, our work re-defines the role of FGF signaling in vertebrate LR axis formation. In parallel with mesoderm induction, FGF specifies superficial organizer cells as LRO precursor during early gastrulation. This holds true despite apparent morphological differences between fish, amphibian and, perhaps, also mouse
DEVEL
O
embryos. As mesodermal tissues undergo morphogenesis beginning at mid gastrulation, FGF signaling, specifically the FGF/Ca2+branch, is instrumental for the generation of flow sensor cells at the lateral margins of the LRO. LR axis formation, thus, seems to be more highly conserved as previously anticipated. It remains to be seen whether this notion holds for flow-sensing mechanisms as well.
MATERIALS AND METHODS Xenopus laeviscare and maintenance
Frogs were purchased from Nasco (Fort Atkinson, WI, USA). Handling, care and experimental manipulations of animals was approved by the Regional
Government Stuttgart, Germany (Vorhaben A379/12 ZO ‘Molekulare
Embryologie’), according to German regulations and laws (§6, article 1,
sentence 2, nr. 4 of the Animal Welfare Act). Animals were kept at the appropriate conditions ( pH 7.7, 20°C) at a 12 h light cycle in the animal facility of the Institute of Zoology of the University of Hohenheim (Stuttgart, Germany). Female frogs (4-15 years old) were stimulated with 25-75 units of human chorionic gonadotropin (hCG; Sigma), depending on weight and age, which was injected subcutaneously 1 week prior to oviposition. On the day before ovulation, female frogs were injected with 300-700 units of hCG (10-12 h before). Eggs were collected into a Petri dish by careful squeezing of the
females andin vitrofertilized. Sperm of male frogs was gained by dissection of
the testes and was stored at 4°C in 1× MBSH (Modified Barth’s saline with
HEPES) solution. Embryos were staged according to Nieuwkoop and Faber (1994). Only clutches of embryos from healthy females were used for the experiments reported here, provided the early embryonic stages showed normal survival rates as well. Individual embryos from one batch were randomly picked and used either as control or tested specimens. If control groups displayed unusual developmental defects later in development, such clutches were excluded as well, based on empirical judgement.
mRNA synthesis and microinjections
Prior to in vitro synthesis of mRNA using the Ambion sp6 message
machine kit, the plasmid was linearized with NotI. Embryos were injected at the 4- to 8-cell stage, using a Harvard Apparatus. Drop size was calibrated to 8 nl/injection. Amounts of injected mRNA or MOs are indicated in the main text. For specific lineage targeting of constructs/MOs, tracers used for injection control included fluorescein (70,000 MW) and rhodamine B dextran (10,000 MW; both Thermo Fisher). [For more detail on lineage-specific injections to target central or lateral GRP and ventro-lateral tissues (i.e. LPM) refer to Blum et al., 2009a; Vick et al., 2018; Vick et al., 2009.]
Treatment with the Fgfr inhibitor SU5402
SU5402 (Calbiochem) was dissolved at 20 mM in DMSO and stock
solutions were stored in aliquots at−20°C. Incubations were conducted in
24-well plates and protected from light at room temperature. Concentrations used ranged from 30 µM to 100 µM (diluted in 0.1× MBSH) as indicated. Incubation start points and duration are indicated by Nieuwkoop and Faber stages in the main text. Incubations were terminated by removal of the inhibitor and several rounds of washes using 0.1× MBSH buffer solution.
Proliferation analysis
DMSO control or 60 µM SU5402-treated embryos from two independent experiments were fixed in 4% paraformaldehyde at stage 13/14 or 16 for 1 h at room temperature, followed by the preparation of dorsal explants. Immunofluorescence with anti-pH3 antibody [ phospho-histone H3 (Ser10), 06-570, Millipore; diluted 1:140 and incubated at 4°C] was used to visualize proliferating cells. Cell borders were highlighted by cortical actin staining using Alexa Fluor 488 Phalloidin (Thermo Fisher; 1:40; 30 min incubation at room temperature) and nuclei were stained with Hoechst 33342 (Thermo Fisher; 1:10,000; 1 h at room temperature).
Analysis of apoptosis by TUNEL staining and Bcl-XL injections TUNEL analyses were performed using standard protocols (Hensey and Gautier, 1998). Dorsal explants of DMSO control specimens or 60 µM SU5402-treated embryos from two independent experiments were fixed at
stage 13/14 or 16 and analyzed for apoptotic cells in the neuroectoderm and GRP. Briefly, fixed embryos were incubated at room temperature overnight with digoxygenin-dUTP (1 µM) and terminal deoxynucleotidyl transferase (150 U/ml). Following incubation with an anti-digoxygenin-AP (alkaline phosphatase) antibody (11093274910, Roche; 1:5000) overnight at 4°C, apoptotic cells were visualized using BM Purple AP substrate (Roche) at room temperature.
The apoptosis rescue experiment using Bcl-XL was performed by targeting 8 pg of Bcl-XL/CS2+ plasmid DNA (Johnston et al., 2005) into the left and right somitic GRP lineages (dorsolateral part of the 4-cell embryo). Embryos were treated with either DMSO or 60 µM SU5402 from
stage 12-17, fixed and processed for in situ hybridization (ISH) using
dand5-andnodal1-specific antisense probes.
ISH
For in situ mRNA detection, whole-mount ISH was performed. Embryos were fixed in MEMFA [3.7% formaldehyde, 100 mM
3-(N-morpholino)propanesulfonic acid, 2 mM EGTA, 1 mM MgSO4] for 2-3 h at
room temperature and processed following standard protocols (Sive et al.,
2000). RNAin situprobes were transcribed using SP6 or T7 polymerases.
Whole-mount ISH protocol was modified from Belo et al. (1997).
SEM
For SEM analysis, injected or treated specimens were fixed with 4%
paraformaldehyde/2.5% glutaraldehyde in Sorenson’s phosphate buffer
overnight at 4°C. Dorsal explants were prepared for SEM and analyzed as previously described (Schweickert et al., 2007; Sulik et al., 1994). SEM photographs were analyzed for GRP ciliation, cilia polarization and apical cell surface area using ImageJ and evaluated as described (Beyer et al., 2012; Sbalzarini and Koumoutsakos, 2005).
Analysis of leftward fluid-flow
For analysis of leftward flow, dorsal posterior GRP explants were dissected from control or SU5402-treated embryos at stage 17. Explants were placed in a Petri dish containing fluorescent microbeads (diameter 0.5 µm; diluted 1:2500 in 1× MBSH) and incubated for a few seconds. Explants were transferred to a microscope slide which was prepared with vacuum grease to create a small chamber that contained a fluorescent microbead solution; a cover slip was carefully pressed on to seal the chamber. Time-lapse movies of leftward flow were recorded using an AxioCam HSm video camera (Zeiss) at two frames per second using an Axioplan2 imaging microscope
(Zeiss). Using the ImageJ ‘Particle-Tracker’ plug-in, leftward flow was
analyzed and particle movement was assessed (Beyer et al., 2012; Sbalzarini and Koumoutsakos, 2005; Vick et al., 2009). For evaluation of flow quality, i.e. directionality of transported fluorescent particles, a custom-made analysis tool written in the statistical software R (R Core Team, 2013) by T. Thumberger and described by Vick et al. (2009) was used. Briefly, a mask covering the GRP was used to exclusively measure beads transported across the GPR. Only particles were evaluated that were detected in ten consecutive frames. To measure directionality of single beads and leftward flow, and to
exclude Brownian movement, a Rayleigh’s test of uniformity was
performed, first on single beads, then on the mean angles of single
particles moving in a directed manner. The parameter rho (ρ) describes the
directionality of flow (i.e. the sum of all remaining particles), withP>0.6
defined as WT andP<0.6 considered as reduced or aberrant flow (cf. Vick
et al., 2009). Diagrams of flow quality show mean direction of all particles
and aP-value as the index of directionality (Fig. S3G-I).
GRP width analyses
For quantification of GRP widths, SEM photographs of GRP explants or
vibratome sections ofmyod1-stained embryos were used. For evaluation of
superficial myod1 expression in the GRP, transversal GRP sections
containing the widest part ofmyod1-positive cells were selected. Widths
were measured in pixels using ImageJ (Sbalzarini and Koumoutsakos, 2005). For each analysis, the mean value of DMSO-treated controls was set to 1.0 and the relative sizes of manipulated embryos was calculated to
generate box plots. For measurements ofmyod1-positive, lateral somitic
GRP widths, the left and right halves were added up.
DEVEL
O
Statistical analysis
Statistical calculations of marker gene expression patterns and cilia
distribution were performed using Pearson’sχ2test (Bonferroni corrected,
if required). For statistical calculation of ciliation, cilia length, GRP widths, flow velocity and directionality Wilcoxon-Match-Pair test was used, for
vesicle abundance per µm2Kruskal–Wallis test, and for left versus right
vesicle abundance and pH3 analysis Mann–Whitney test. For calculations,
the statistical software R was used (R Core Team, 2013; https://www.r-project.org).
In all figures, statistical results are represented as *P<0.05, **P<0.01, ***P<0.001.
Acknowledgements
We thank T. Thumberger for expert help with flow and GRP analysis, M. Tingler for help withpkd2epistasis analysis, T. Beyer for technical advice, and E. Amaya, J. Heasman, D. Kessler, C. Kintner, R. Rupp, J. Smith, H. Steinbeisser and A. Vonica for constructs.
Competing interests
The authors declare no competing or financial interests.
Author contributions
Conceptualization: A.S., M.B., P.V.; Validation: I.S., J.K., P.V.; Formal analysis: I.S.; Investigation: I.S., J.K., P.V.; Writing - original draft: P.V.; Writing - review & editing: I.S., J.K., M.B., P.V.; Visualization: I.S., J.K., P.V.; Supervision: A.S., M.B., P.V.; Funding acquisition: M.B.
Funding
M.B. was funded by the Deutsche Forschungsgemeinschaft (BL285/9-2). J.K. was a recipient of a Ph.D. fellowship from the Landesgraduiertenförderung Baden-Württemberg.
Supplementary information
Supplementary information available online at
http://dev.biologists.org/lookup/doi/10.1242/dev.173575.supplemental
References
Akbulut, S., Reddi, A. L., Aggarwal, P., Ambardekar, C., Canciani, B., Kim, M. K. H., Hix, L., Vilimas, T., Mason, J., Basson, M. A. et al.(2010). Sprouty proteins inhibit receptor-mediated activation of phosphatidylinositol-specific phospholipase C.Mol. Biol. Cell21, 3487-3496. doi:10.1091/mbc.e10-02-0123
Amack, J. D. and Yost, H. J.(2004). The T box transcription factor no tail in ciliated cells controls zebrafish left-right asymmetry.Curr. Biol.14, 685-690. doi:10.1016/ j.cub.2004.04.002
Amaya, E., Musci, T. J. and Kirschner, M. W.(1991). Expression of a dominant negative mutant of the FGF receptor disrupts mesoderm formation in Xenopus embryos.Cell66, 257-270. doi:10.1016/0092-8674(91)90616-7
Amaya, E., Stein, P. A., Musci, T. J. and Kirschner, M. W.(1993). FGF signalling in the early specification of mesoderm in Xenopus.Development118, 477-487.
Belo, J. A., Bouwmeester, T., Leyns, L., Kertesz, N., Gallo, M., Follettie, M. and De Robertis, E. M.(1997). Cerberus-like is a secreted factor with neuralizing activity expressed in the anterior primitive endoderm of the mouse gastrula.Mech. Dev.68, 45-57. doi:10.1016/S0925-4773(97)00125-1
Berridge, M. J., Lipp, P. and Bootman, M. D. (2000). The versatility and universality of calcium signalling.Nat. Rev. Mol. Cell Biol.1, 11-21. doi:10.1038/ 35036035
Beyer, T., Danilchik, M., Thumberger, T., Vick, P., Tisler, M., Schneider, I., Bogusch, S., Andre, P., Ulmer, B., Walentek, P. et al. (2012). Serotonin signaling is required for Wnt-dependent GRP specification and leftward flow in Xenopus.Curr. Biol.22, 33-39. doi:10.1016/j.cub.2011.11.027
Blum, M. and Ott, T.(2018). The power of strain: organizing left-right cilia.Dev. Cell 45, 277-279. doi:10.1016/j.devcel.2018.04.015
Blum, M., Andre, P., Muders, K., Schweickert, A., Fischer, A., Bitzer, E., Bogusch, S., Beyer, T., van Straaten, H. W. M. and Viebahn, C.(2007). Ciliation and gene expression distinguish between node and posterior notochord in the mammalian embryo.Differentiation75, 133-146. doi:10.1111/j.1432-0436.2006. 00124.x
Blum, M., Beyer, T., Weber, T., Vick, P., Andre, P., Bitzer, E. and Schweickert, A.
(2009a). Xenopus, an ideal model system to study vertebrate left-right asymmetry.
Dev. Dyn.238, 1215-1225. doi:10.1002/dvdy.21855
Blum, M., Weber, T., Beyer, T. and Vick, P.(2009b). Evolution of leftward flow.
Semin. Cell Dev. Biol.20, 464-471. doi:10.1016/j.semcdb.2008.11.005
Blum, M., Schweickert, A., Vick, P., Wright, C. V. E. and Danilchik, M. V.(2014). Symmetry breakage in the vertebrate embryo: when does it happen and how does it work?Dev. Biol.393, 109-123. doi:10.1016/j.ydbio.2014.06.014
Boettger, T., Wittler, L. and Kessel, M.(1999). FGF8 functions in the specification of the right body side of the chick.Curr. Biol.9, 277-280. doi:10.1016/S0960-9822(99)80119-5
Boskovski, M. T., Yuan, S., Pedersen, N. B., Goth, C. K., Makova, S., Clausen, H., Brueckner, M. and Khokha, M. K.(2013). The heterotaxy gene GALNT11 glycosylates Notch to orchestrate cilia type and laterality.Nature504, 456-459. doi:10.1038/nature12723
Busch, T., Köttgen, M. and Hofherr, A.(2017). TRPP2 ion channels: critical regulators of organ morphogenesis in health and disease.Cell Calcium66, 25-32. doi:10.1016/j.ceca.2017.05.005
Caron, A., Xu, X. and Lin, X.(2012). Wnt/β-catenin signaling directly regulates Foxj1 expression and ciliogenesis in zebrafish Kupffer’s vesicle.Development 139, 514-524. doi:10.1242/dev.071746
Chien, Y.-H., Srinivasan, S., Keller, R. and Kintner, C.(2018). Mechanical strain determines cilia length, motility, and planar position in the left-right organizer.Dev. Cell45, 316-330.e4. doi:10.1016/j.devcel.2018.04.007
Dasgupta, A. and Amack, J. D.(2016). Cilia in vertebrate left-right patterning.
Philos. Trans. R. Soc. Lond., B, Biol. Sci.371, 20150410. doi:10.1098/rstb.2015. 0410
Delling, M., Indzhykulian, A. A., Liu, X., Li, Y., Xie, T., Corey, D. P. and Clapham, D. E.(2016). Primary cilia are not calcium-responsive mechanosensors.Nature 531, 656-660. doi:10.1038/nature17426
Dorey, K. and Amaya, E. (2010). FGF signalling: diverse roles during early vertebrate embryogenesis. Development 137, 3731-3742. doi:10.1242/dev. 037689
Feistel, K. and Blum, M.(2008). Gap junctions relay FGF8-mediated right-sided repression of Nodal in rabbit.Dev. Dyn.237, 3516-3527. doi:10.1002/dvdy.21535
Ferreira, R. R., Vilfan, A., Jülicher, F., Supatto, W. and Vermot, J.(2017). Physical limits of flow sensing in the left-right organizer.eLife6, e25078. doi:10. 7554/eLife.25078
Fischer, A., Viebahn, C. and Blum, M.(2002). FGF8 acts as a right determinant during establishment of the left-right axis in the rabbit.Curr. Biol.12, 1807-1816. doi:10.1016/S0960-9822(02)01222-8
Fletcher, R. B. and Harland, R. M.(2008). The role of FGF signaling in the establishment and maintenance of mesodermal gene expression in Xenopus.
Dev. Dyn.237, 1243-1254. doi:10.1002/dvdy.21517
Hensey, C. and Gautier, J. (1998). Programmed cell death during Xenopus development: a spatio-temporal analysis.Dev. Biol.203, 36-48. doi:10.1006/dbio. 1998.9028
Hong, S.-K. and Dawid, I. B. (2009). FGF-dependent left-right asymmetry patterning in zebrafish is mediated by Ier2 and Fibp1.Proc. Natl. Acad. Sci. USA106, 2230-2235. doi:10.1073/pnas.0812880106
Johnston, J., Chan, R., Calderon-Segura, M., McFarlane, S. and Browder, L. W.
(2005). The roles of Bcl-xL in modulating apoptosis during development of Xenopus laevis.BMC Dev. Biol.5, 20. doi:10.1186/1471-213X-5-20
King, T., Beddington, R. S. P. and Brown, N. A.(1998). The role of the brachyury gene in heart development and left-right specification in the mouse.Mech. Dev. 79, 29-37. doi:10.1016/S0925-4773(98)00166-X
Koulen, P., Cai, Y., Geng, L., Maeda, Y., Nishimura, S., Witzgall, R., Ehrlich, B. E. and Somlo, S.(2002). Polycystin-2 is an intracellular calcium release channel.
Nat. Cell Biol.4, 191-197. doi:10.1038/ncb754
Li, Y., Wright, J. M., Qian, F., Germino, G. G. and Guggino, W. B.(2005). Polycystin 2 interacts with type I inositol 1,4,5-trisphosphate receptor to modulate intracellular Ca2+signaling.J. Biol. Chem.280, 41298-41306. doi:10.1074/jbc.
M510082200
McGrath, J., Somlo, S., Makova, S., Tian, X. and Brueckner, M.(2003). Two populations of node monocilia initiate left-right asymmetry in the mouse.Cell114, 61-73. doi:10.1016/S0092-8674(03)00511-7
Meyers, E. N. and Martin, G. R.(1999). Differences in left-right axis pathways in mouse and chick: functions of FGF8 and SHH.Science285, 403-406. doi:10. 1126/science.285.5426.403
Namigai, E. K. O., Kenny, N. J. and Shimeld, S. M.(2014). Right across the tree of life: the evolution of left-right asymmetry in the Bilateria.Genesis52, 458-470. doi:10.1002/dvg.22748
Neugebauer, J. M., Amack, J. D., Peterson, A. G., Bisgrove, B. W. and Yost, H. J.
(2009). FGF signalling during embryo development regulates cilia length in diverse epithelia.Nature458, 651-654. doi:10.1038/nature07753
Nieuwkoop, P. D. and Faber, J.(1994).Normal Table of Xenopus laevis, New York: Garland.
Nonaka, S., Tanaka, Y., Okada, Y., Takeda, S., Harada, A., Kanai, Y., Kido, M. and Hirokawa, N.(1998). Randomization of left-right asymmetry due to loss of nodal cilia generating leftward flow of extraembryonic fluid in mice lacking KIF3B motor protein.Cell95, 829-837. doi:10.1016/S0092-8674(00)81705-5
Nonaka, S., Shiratori, H., Saijoh, Y. and Hamada, H.(2002). Determination of left-right patterning of the mouse embryo by artificial nodal flow.Nature418, 96-99. doi:10.1038/nature00849
Nutt, S. L., Dingwell, K. S., Holt, C. E. and Amaya, E.(2001). Xenopus Sprouty2 inhibits FGF-mediated gastrulation movements but does not affect mesoderm induction and patterning.Genes Dev.15, 1152-1166. doi:10.1101/gad.191301
DEVEL
O
Okada, Y., Nonaka, S., Tanaka, Y., Saijoh, Y., Hamada, H. and Hirokawa, N.
(1999). Abnormal nodal flow precedes situs inversus in iv and inv mice.Mol. Cell 4, 459-468. doi:10.1016/S1097-2765(00)80197-5
Oki, S., Kitajima, K. and Meno, C.(2010). Dissecting the role of Fgf signaling during gastrulation and left-right axis formation in mouse embryos using chemical inhibitors.Dev. Dyn.239, 1768-1778. doi:10.1002/dvdy.22282
Pineda-Salgado, L., Craig, E. J., Blank, R. B. and Kessler, D. S. (2005). Expression of Panza, an alpha2-macroglobulin, in a restricted dorsal domain of the primitive gut in Xenopus laevis.Gene Expr. Patterns6, 3-10. doi:10.1016/j. modgep.2005.09.001
Pownall, M. E. and Isaacs, H. V.(2010).FGF Signalling in Vertebrate Development. San Rafael, CA: Morgan and Claypool Life Sciences. doi:10.4199/ C00011ED1V01Y201004DEB002
R Core Team(2013).R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing.
Saka, Y. and Smith, J. C.(2001). Spatial and temporal patterns of cell division during early Xenopus embryogenesis.Dev. Biol.229, 307-318. doi:10.1006/dbio. 2000.0101
Sbalzarini, I. F. and Koumoutsakos, P. (2005). Feature point tracking and trajectory analysis for video imaging in cell biology.J. Struct. Biol.151, 182-195. doi:10.1016/j.jsb.2005.06.002
Schlessinger, J.(2000). Cell signaling by receptor tyrosine kinases.Cell103, 211-225. doi:10.1016/S0092-8674(00)00114-8
Schulte-Merker, S. and Smith, J. C.(1995). Mesoderm formation in response to Brachyury requires FGF signalling. Curr. Biol. 5, 62-67. doi:10.1016/S0960-9822(95)00017-0
Schweickert, A., Campione, M., Steinbeisser, H. and Blum, M.(2000). Pitx2 isoforms: involvement of Pitx2c but not Pitx2a or Pitx2b in vertebrate left-right asymmetry.Mech. Dev.90, 41-51. doi:10.1016/S0925-4773(99)00227-0
Schweickert, A., Weber, T., Beyer, T., Vick, P., Bogusch, S., Feistel, K. and Blum, M.(2007). Cilia-driven leftward flow determines laterality in Xenopus.Curr. Biol.17, 60-66. doi:10.1016/j.cub.2006.10.067
Schweickert, A., Vick, P., Getwan, M., Weber, T., Schneider, I., Eberhardt, M., Beyer, T., Pachur, A. and Blum, M.(2010). The nodal inhibitor coco is a critical target of leftward flow in Xenopus.Curr. Biol.20, 738-743. doi:10.1016/j.cub. 2010.02.061
Shook, D. R., Majer, C. and Keller, R.(2004). Pattern and morphogenesis of presumptive superficial mesoderm in two closely related species, Xenopus laevis and Xenopus tropicalis.Dev. Biol.270, 163-185. doi:10.1016/j.ydbio.2004.02.021
Sivak, J. M., Petersen, L. F. and Amaya, E.(2005). FGF signal interpretation is directed by Sprouty and Spred proteins during mesoderm formation.Dev. Cell8, 689-701. doi:10.1016/j.devcel.2005.02.011
Sive, H. L., Grainger, R. M. and Harland, R. M.(2000).Early Development of Xenopus Laevis. CSHL Press.
Smith, W. C., McKendry, R., Ribisi, S. and Harland, R. M.(1995). A nodal-related gene defines a physical and functional domain within the Spemann organizer.Cell 82, 37-46. doi:10.1016/0092-8674(95)90050-0
Stubbs, J. L., Oishi, I., Izpisúa-Belmonte, J. C. and Kintner, C.(2008). The forkhead protein Foxj1 specifies node-like cilia in Xenopus and zebrafish embryos.Nat. Genet.40, 1454-1460. doi:10.1038/ng.267
Sulik, K., Dehart, D. B., Inagaki, T., Carson, J. L., Vrablic, T., Gesteland, K. and Schoenwolf, G. C.(1994). Morphogenesis of the murine node and notochordal plate.Dev. Dyn.201, 260-278. doi:10.1002/aja.1002010309
Tabin, C. J. and Vogan, K. J.(2003). A two-cilia model for vertebrate left-right axis specification.Genes Dev.17, 1-6. doi:10.1101/gad.1053803
Takao, D., Nemoto, T., Abe, T., Kiyonari, H., Kajiura-Kobayashi, H., Shiratori, H. and Nonaka, S.(2013). Asymmetric distribution of dynamic calcium signals in the node of mouse embryo during left–right axis formation.Dev. Biol.376, 23-30. doi:10.1016/j.ydbio.2013.01.018
Tanaka, Y., Okada, Y. and Hirokawa, N.(2005). FGF-induced vesicular release of Sonic hedgehog and retinoic acid in leftward nodal flow is critical for left-right determination.Nature435, 172-177. doi:10.1038/nature03494
Thastrup, O., Cullen, P. J., Drøbak, B. K., Hanley, M. R. and Dawson, A. P.
(1990). Thapsigargin, a tumor promoter, discharges intracellular Ca2+ stores by specific inhibition of the endoplasmic reticulum Ca2(+)-ATPase.Proc. Natl Acad. Sci. USA87, 2466-2470. doi:10.1073/pnas.87.7.2466
Tran, U., Zakin, L., Schweickert, A., Agrawal, R., Döger, R., Blum, M., De Robertis, E. M. and Wessely, O.(2010). The RNA-binding protein bicaudal C regulates polycystin 2 in the kidney by antagonizing miR-17 activity.Development 137, 1107-1116. doi:10.1242/dev.046045
Vick, P., Schweickert, A., Weber, T., Eberhardt, M., Mencl, S., Shcherbakov, D., Beyer, T. and Blum, M.(2009). Flow on the right side of the gastrocoel roof plate is dispensable for symmetry breakage in the frog Xenopus laevis.Dev. Biol.331, 281-291. doi:10.1016/j.ydbio.2009.05.547
Vick, P., Kreis, J., Schneider, I., Tingler, M., Getwan, M., Thumberger, T., Beyer, T., Schweickert, A. and Blum, M.(2018). An early function of polycystin-2 for left-right organizer induction in Xenopus.iScience2, 76-85. doi:10.1016/j.isci.2018. 03.011
Vonica, A. and Brivanlou, A. H.(2007). The left–right axis is regulated by the interplay of Coco, Xnr1 and derrière in Xenopus embryos.Dev. Biol.303, 281-294. doi:10.1016/j.ydbio.2006.09.039
Wallingford, J. B., Ewald, A. J., Harland, R. M. and Fraser, S. E.(2001). Calcium signaling during convergent extension in Xenopus.Curr. Biol.11, 652-661. doi:10. 1016/S0960-9822(01)00201-9
Wang, Y. and Steinbeisser, H.(2009). Molecular basis of morphogenesis during vertebrate gastrulation.Cell. Mol. Life Sci.66, 2263-2273. doi:10.1007/s00018-009-0018-2
Yamauchi, H., Miyakawa, N., Miyake, A. and Itoh, N.(2009). Fgf4 is required for left-right patterning of visceral organs in zebrafish.Dev. Biol.332, 177-185. doi:10. 1016/j.ydbio.2009.05.568
Yeo, W. and Gautier, J.(2004). Early neural cell death: dying to become neurons.
Dev. Biol.274, 233-244. doi:10.1016/j.ydbio.2004.07.026
Yokota, C., Kofron, M., Zuck, M., Houston, D. W., Isaacs, H., Asashima, M., Wylie, C. C. and Heasman, J.(2003). A novel role for a nodal-related protein; Xnr3 regulates convergent extension movements via the FGF receptor.
Development130, 2199-2212. doi:10.1242/dev.00434
Yoshiba, S. and Hamada, H.(2014). Roles of cilia, fluid flow, and Ca2+ signaling in breaking of left-right symmetry.Trends Genet.30, 10-17. doi:10.1016/j.tig.2013. 09.001
Yoshiba, S., Shiratori, H., Kuo, I. Y., Kawasumi, A., Shinohara, K., Nonaka, S., Asai, Y., Sasaki, G., Belo, J. A., Sasaki, H. et al.(2012). Cilia at the node of mouse embryos sense fluid flow for left-right determination via Pkd2.Science338, 226-231. doi:10.1126/science.1222538