Copyright2001 by the Genetics Society of America
Localized Remodeling of the
Escherichia coli
Chromosome: The Patchwork of
Segments Refractory and Tolerant to Inversion Near the Replication Terminus
Maria Isabel Guijo,* Josette Patte,
†Maria del Mar Campos,*
Jean-Michel Louarn
†and Jose´ Emilio Rebollo*
*Departamento de Bioquimica y Biologia Molecular y Gene´tica, Facultad de Ciencias, Universidad de Extremadura, 06080 Badajoz, Spain and†Laboratoire de Microbiologie et de Ge´ne´tique Mole´culaires du C.N.R.S., 31062 Toulouse Cedex, France
Manuscript received October 5, 2000 Accepted for publication November 30, 2000
ABSTRACT
The behavior of chromosomal inversions inEscherichia colidepends upon the region they affect. Regions flanking the replication terminus have been termed nondivisible zones (NDZ) because inversions ending in the region were either deleterious or not feasible. This regional phenomenon is further analyzed here. Thirty segments distributed between 23 and 29 min on the chromosome map have been submitted to an inversion test. Twenty-five segments either became deleterious when inverted or were noninvertible, but five segments tolerated inversion. The involvement of polar replication pause sites in this distribution was investigated. The results suggest that the Tus/pause site system may forbid some inversion events, but that other constraints to inversion, unrelated to this system, exist. Our current model for deleterious inversions is that the segments involved carry polar sequences acting in concert with other polar sequences located outside the segments. The observed patchwork of refractory and tolerant segments supports the existence of several NDZs in the 23- to 29-min region. Microscopic observations revealed that deleterious inversions are associated with high frequencies of abnormal nucleoid structure and distribution. Combined with other information, the data suggest that NDZs participate in the organization of the terminal domain of the nucleoid.
T
WO sites on the circular Escherichia coli chromo- tion within the cell and the existence of nondivisible zones (NDZs) whose disruption by inversion is delete-some play a major role during the cell cycle:oriC,where replisomes are assembled for bidirectional repli- rious.
Genetic studies of requirements for difactivity have cation (Messer and Weigler 1996, for review), and
revealed that dimer resolution is efficient only whendif
the diametrically oppositedifsite, where chromosome
is located within a limited region between replication dimers, resulting from an odd number of exchanges
arms, named DAZ for Difactivity zone, and when the between sister chromosomes, are resolved (Kuempelet
extensive regions flanking the site remain in their
natu-al.1991; Steiner and Kuempel 1998). The existence
ral orientation with respect to replication (Correet al.
of a long-range nucleoid organization on each of the
2000;Peralset al.2000). The orientation requirement chromosome arms joining these sites has been long
is presumably related to the mechanism that positions suspected (Louarnet al. 1982;Rebolloet al.1988) and
sisterdifsites near the growing septum where resolution is now supported by an increasing body of evidence.
is supposed to occur (Steiner et al. 1999). Our data Cytological studies have shown that origin and terminus
suggested that multiple oriented short sequences act in regions move within the cell with different
choreogra-a concerted mchoreogra-anner to locchoreogra-ate the DAZ choreogra-at the right plchoreogra-ace phy as nucleoids are replicated and partitioned,
sug-(in the septal plane) and at the right moment of the cell gesting the existence of two macrodomains in the
cycle (when the septum closes) and that such sequence nucleoid, the Ori and Ter domains (Nikiet al. 2000).
elements generate a functional long-range polarization Factors involved in the organization into
macro-of a significant fraction macro-of each chromosome arm on domains have not yet been directly approached.
How-either side of dif. The functional polarization of the ever, the region corresponding to the Ter domain is
terminus region in opposite directions fromdifmay be characterized by two phenomena that might contribute
correlated with the fact that a variety of small sequences to this organization: a regional control on the activity
are preferentially found on the chromosome strands of thedifsite that probably involves chromosome
migra-running 5⬘to 3⬘fromoriCtodif. First described by Salz-berget al.(1998), skewed oligomers of the RRNAGGGS family (R⫽purine, N⫽any base, S⫽G or C) display
Corresponding author:Jean-Michel Louarn, Laboratoire de Microbi- all topographical requirements expected for motifs de-ologie et de Ge´ne´tique Mole´culaires du CNRS, 118 route de
Nar-termining dif activity, though direct proof of their bonne, 31062 Toulouse Cedex, France.
E-mail: [email protected] involvement is still lacking (Capiauxet al.2001).
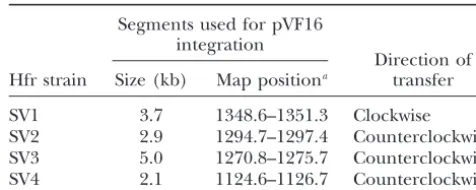
TABLE 1 Konrad(1977) was first to notice that the segment
between thelacoperon andattφ80 on theE. colichromo- New Hfr strains: integration sites oforiTplasmids some (a region we further analyze here) is refractory and directions of transfer
to inversion.Rebolloet al.(1988) extended this
obser-vation and analyzed the consequences of inversion of Segments used for pVF16 integration many segments covering the entireE. colichromosome.
Direction of They described three behaviors: segments may be either Hfr strain Size (kb) Map positiona transfer
(i) invertible and tolerant to inversion (type T, the
in-SV1 3.7 1348.6–1351.3 Clockwise
verted state is stable over generations); (ii) invertible
SV2 2.9 1294.7–1297.4 Counterclockwise
but refractory (type R, the inverted state is detrimental);
SV3 5.0 1270.8–1275.7 Counterclockwise
or (iii) nonpermissive (type N, either the inversion is not SV4 2.1 1124.6–1126.7 Counterclockwise feasible or so strongly disabling that it is never detected).
aOrdinates of Ecomap7.
The striking observation was that deleterious inversions have one endpoint located in the 20–30% of theE. coli
chromosome flanking the replication terminus; these
conjugation assays, LN1902, is W945 ara leu lacY1 purE gal regions behaved differently from the rest of the chromo- pyrC76::Tn10(TcR)trp zdd230::Tn9(CmR)his gyrA(NalR)argG some and were termed nondivisible zones. The NDZs rpsL(SmR)xyl mtl ile metA/B. Plasmid pVF16, constructed by V. Francois to facilitate the creation of new Hfr’s, is composed also harbor multiple replication pause sites that, when
of a 2.7-kb fragment from pSC101 carrying a thermosensitive occupied by the Tus protein, are polar inhibitors of
replication origin (Hashimoto-GotoandSekiguchi1977) replication fork movement (Hill1996). In earlier
ex-associated with a 1-kb fragment from plasmid F carrying the periments Tus and pause sites appeared not to be in- origin of transferoriT, a multilinker for cloning, and a 1.85-volved in the NDZ phenomenon because inversions kb interposon providing resistance to chloramphenicol (
Fel-layet al. 1987). Plasmids pGR1 to pGR4 are derivatives of ending in the NDZs remained nonpermitted or
deleteri-pVF16 by cloning in the chromosomal segments listed in Table ous in tus-deleted strains (Francoiset al. 1990a,b). It
1. Plasmid pOX38::ApR(Guyeret al.1980) was kindly provided was proposed that the detriment caused by inverted
by M. Chandler. Bacteriophage TSK was used to replace segments of the NDZs could be an alteration of a higher- Tn10insertions by their Tes or Tek derivatives, as previously order nucleoid organization involving interactions be- described (Francois et al.1987;Rebolloet al. 1988). P1vir bacteriophage was used for generalized transduction of chro-tween ordered sequences (Rebollo et al. 1988). This
mosomal markers, as described inCaroandBerg(1971). was the first reported example of long-range
interac-Growth media and genetic procedures:Luria L rich medium tions in the chromosome.
and Vogel and Bonner E synthetic medium are described in In this work, we further analyze the nondivisible zone Miller(1992). Antibiotic concentrations used are as follows: NDZ1 (located between 20 and 30 min on the genetic ampicillin (Ap), 50g/ml; chloramphenicol (Cm), 25 g/ ml; kanamycin (Km), 25g/ml; nalidixic acid (Nal), 20g/ map) to determine more precisely the positions of
seg-ml; streptomycin (Sm), 20g/ml (E medium) or 10g/ml ments whose inversion is detrimental and to examine
(L medium); and tetracyclin (Tc), 10g/ml (E) or 50g/ml possible connections between NDZ and DAZ
phenom-(L). Allin vivoorin vitrogenetic experiments were performed ena. Inversions between predetermined positions were according to standard procedures (Sambrook et al. 1989; sought using a genetic system based on modified Tn10 Miller1992).
Tn10mutagenesis:New Tn10insertions in NDZ1 were iso-insertions (Rebolloet al. 1988). The results indicate the
lated from a pool of random Tn10insertions in strain CB0129, existence of several regions harboring elements whose
kindly provided by C. Labi and constructed as described in concerted arrangement is functionally important,
irre-Wayet al. (1984). The TcRresistance character was cotrans-spective of gene content. Splitting such clusters of se- duced with prototrophy to pyrC (24.2 min), trp(28.3 min), quences causes abnormalities in the shape and distribu- andpyrF(28.9 min) mutant strains via bacteriophage P1. Other Tn10insertions were gifts from B. Bachmann or C. Gutierrez. tion of nucleoids that are the probable reasons for the
Tn10-Tes or -Tek substitutions:Tn10derivatives carrying a disturbed growth of such bacteria. These observations
streptomycin/spectinomycin resistance (SmR/SpR)⍀ interpo-further support the model that inversions may be
delete-son in the proximal part of thetetAlocus (the Tes transposon) rious because they disturb the interplay of multiple se- or a kanamycin resistance (KmR) fragment in the distal part quence elements governing chromosome organization oftetA(the Tek transposon) were constructedin vivousing recombinant phageTSK (Francoiset al.1987;Rebolloet and its processing.
al. 1988). Strains harboring a Tes plus a Tek transposon were systematically constructed by adding, via P1 transduction, the Tek transposon to an already present Tes transposon. Since MATERIALS AND METHODS
the presence of a single copy of the SpR/SmR⍀interposon in the terminus region confers weak resistance to
streptomy-Bacterial, plasmid, and bacteriophage strains: Inversions
were engineered in derivatives of strain CB0129 (W1485 F⫺ cin, and virtually no resistance to spectinomycin, we avoided as much as possible direct selection for SpR/SmR bacteria, leu thy deoBorC supD;Birdet al. 1972) and are referred to
by their Tes and Tek insertions. One exception is INV(aroA- which may favor duplication of the resistance locus or other undesired rearrangements.
fadR), which was engineered in Hfr KL19. Hfr LN2085, also
a CB0129 derivative, was described earlier (Francois et al. Selection of chromosomal inversions:Figure 1A illustrates the genetic selection used in the inversion search. In a TcS 1990c); it promotes chromosome transfer from thetrpoperon
Figure1.—Genetic tools for selection and analysis of chro-mosomal inversions. (A) The Tes and Tek system. Tes and Tek transposons (large arrows), in-serted in opposite orientations in the chromosome,can recom-bine to give an easily selectable TcR determinant. The ex-change indicated by a double arrow may result, if reciprocal, in the inversion of the chromo-somal segment separating the Tes and Tek insertions, with re-constitution of a wild-type Tn10. Inversion may be detected in conjugation experiments by the inverted order of transfer fromoriTof markers X and Y external to the segment in-verted. (B) Mapping and orien-tation of Tn10 insertions and oforiT-carrier plasmid integra-tion sites. The upper line shows a simplified version of the E. colichromosome Ecomap7 be-tween 1050 and 1400 kb ( Ber-lynet al.1996). Flags indicate the locations of the three pause sites of the region, all inhib-iting leftward moving replica-tion forks. Tn10insertions are indicated by arrows oriented from IS10L to IS10R. Vertical dotted lines indicate their map-ping, estimated from both phy-sical and genetic data. They are designated in the “z” naming system assorted from a number recalling their position in kilo-bases on Ecomap7. The map positions of Tn10insertions in known genes are given be-tween brackets. Small rectan-gles with arrowheads superim-posed on the map indicate the positions of the integrated pVF16 (oriT) derivatives, with directions of transfer symbolized by an arrowhead (Hfr’s SV1 to SV4; see text). (C) Typical Hfr organization. Here Hfr SV1 is represented, resulting from integrative recombination of pGR1 into the chromosome, with vector pVF16 DNA flanked by direct repeats of the chromosomal segment cloned in pGR1.
inserted in opposite orientations, a globally reciprocal homol- that an F plasmid origin of transfer is located within the seg-ment analyzed (Figure 1). This was obtained using plasmid ogous exchange occurring between the 0.9-kb segments
sepa-rating the KmRand SmRinsertion positions of the Tes and pVF16, arepA(Ts) derivative of pSC101 that carries an F plas-mid origin of transfer,oriT, and a gene for resistance to chlor-Tek sequences may yield a selectable TcRrecombinant with
inversion of the chromosomal segment separating the transpo- amphenicol. Four segments of the 23- to 30-min region were cloned into this vector (Table 1 and Figure 1), and the re-sons. Inversions being preferentially found among TcRSmR
KnRtriply resistant recombinants (Francoiset al.1987, 1990a; sulting plasmids were integrated into the chromosome by ho-mologous recombination, selecting for CmRclones at 42⬚(the
Rebolloet al. 1988), selection for TcRrecombinants was in
general done in the presence of kanamycin, then screening expected structure is shown in Figure 1C). This provided insertion oforiTat convenient positions. Thetrans-acting trans-for the SmRcharacter and finally a search for inversions in
this class of indigenous recombinants were performed. Triply fer functions were eventually provided by the conjugation-proficient F factor derivative pOX38::ApR(Guyeret al. 1980). resistant clones were picked up, resuspended in 0.5 ml of
Luria broth (LB) medium, and incubated without aeration These Hfr’s form an isogenic strain family and the small size of theoriTplasmid minimizes local perturbation, but they are for 1 hr at 37⬚before the inversion test.
transfers its chromosome clockwise and the three others (SV2, trp (28.3 min), or pyrF (28.9 min) markers. Fourteen SV3, SV4) transfer counterclockwise. For the conjugation test, out of 70 isolates were retained for further analysis after the triply resistant Hfr bacteria were mixed with F⫺LN1902
mapping by measurements of cotransduction frequen-bacteria grown in LB (one male for five females), incubated
cies with thepyrC,trp, orpyrFmarkers and by Southern for 1 hr, and then plated after convenient dilution on minimal
medium selecting for PurE⫹NalRor His⫹NalRexconjugants. analysis ofPstI,HindIII, andEcoRI segments responding The selected markers are located on either side of the region to a Tn10probe. Some suitably positioned Tn10 inser-analyzed (purEat 12 min andhisat 45 min). tions previously isolated were added to this list. Tn10
Microscopy:Nucleoids of living bacteria were visualized by
orientation within the chromosome was determined by 4⬘,6-diamidino-2-phenylindole (DAPI) staining. DAPI was
chromosome mobilization using a Tn10-carrying conju-added at a final concentration of 2g ml⫺1to cells
exponen-tially growing in LB medium at OD560 nmofⵑ0.3. The cultures gative plasmid (Rebolloet al. 1988) and was confirmed were chilled on ice 20 min later, centrifuged, and concen- by Southern analysis. This set of Tn10insertions is pre-trated 10-fold in DAPI-containing (2g ml⫺1) LB medium.
sented in Figure 1B. The procedure for generating in-A 1- to 2-l aliquot was then spread on a microscope slide and
versions, presented in materials and methods and observed under combined phase-contrast and fluorescence
Figure 1A, required that the Tn10insertions were even-emission microscopy (Leica DRMB microscope; final
magni-fication,⫻2450). tually converted into Tes and Tek derivatives. An
addi-tional requirement was that an oriT plasmid must be present between the Tes and Tek insertions in opposite RESULTS
orientations. The set oforiTinsertions used, presented in Table 1 and Figure 1B, has restricted our analysis to
Establishment of inversions in the 23- to 29-min
re-gion:New Tn10insertions in this region were selected the 30 segments shown in Figure 2.
Bidirectional transfer indicates a deleterious
inver-from a pool of random Tn10 insertions by phage
P1-mediated cotransduction with either pyrC (24.2 min), sion:For nearly half the segments assayed, a variable but
TABLE 2 evolved toward a pattern of transfer identical to that of the noninverted ancestor at rates that, although
vari-Evolution with time of clones promoting early bidirectional
able, did not fit in the no-selection model. If the two
transfer: relative transfer ofpurEⴙandhisⴙmarkers
types grew with similar generation times, the initial data of Table 2 (see also Rebollo et al. 1988) would be Time of growth before crossing
compatible with a frequency of return to wild-type order
Segment 0 hr 1 hr 2 hr 4 hr 7 hr
as high as 1–2% per cell generation, yielding popula-zce-1138-pyrF(B13) 1.6 0.85 0.53 0.22 0.03 tions containing roughly equal numbers of wild-type fadR-trpB(A2) 0.75 0.17 0.03 0.02 0.04 and inversion-type bacteria at the time of testing the recombinant colonies (ⵑ30 generations after the initial Cultures of two Hfr SV2 derivatives carrying Tes and Tek
inversion event). Even with such high recombination at the indicated positions were plated on Tc-Kn L plates. The
smallest colonies found after overnight incubation at 42⬚were frequencies, additional growth for a few generations picked up and resuspended in 2 ml L medium plus 15% would not change significantly the proportion of each glycerol. An aliquot was used to test for streptomycin resis- type in this model. The clone evolution reported in tance, and the rest of the culture was immediately frozen at
Table 2 is clearly much too rapid and cannot be
ex-⫺70⬚. Next day, for each strain, one of the triply resistant
plained without assuming a growth handicap of the clones thus isolated was thawed and incubated at 42⬚to restart
growth. At an OD540 nmof 0.5 (time 0), an aliquot of the culture bacteria carrying the inversion. The fastest evolution was was crossed with LN1902 recipient bacteria as described in observed for INV(fadR-trp) (A2 in Figure 2). Computer
material and methods, with eventual selection for purE⫹ simulations indicate that this evolution rate may be ob-NalRandhis⫹Nalrrecombinants. Meanwhile, the cultures were
served when the wild-type revertant grows 2.5 times maintained in exponential phase of growth by successive
dilu-faster than the original inversion-type bacterium and tions in prewarmed L medium, and conjugation tests were
repeated at each indicated time. The parental Hfr strain trans- occurs at a frequency of 10⫺4per cell generation of the fers thehismarkerⵑ100 times more frequently than thepurE inversion type. In this example, most of the recombinant marker in the same conditions (not shown). The data at time clones tested were classified as noninverted (Table 2). 0 are thus consistent with approximately equal numbers of
In the other example analyzed (Table 3, segment B13 inverted and noninverted bacteria in the clones tested. After
in Figure 2), the accumulation of wild-type bacteria, 2 hr of additional growth in LB medium at 37⬚in the case of
the clone issued from an A2 inversion bacterium, or after 7 hr though less rapid than for A2, also could not fit in in the case of the clone issued from a B13 inversion bacterium, the no-selection model. For other inversions, such as almost no inverted bacteria could be detected in either cul- INV(narI–zci-1385) (B3), the results reported in Table ture.
2 suggest that clone evolution is even slower, since some clones still showed the inversion phenotype. Growth defects attached to inversions may thus have variable important fraction of the triply resistant recombinants
displayed bidirectional transfer in the conjugation test: intensities, but, since the frequency of mixed clones depends both on the difference in generation times both purE⫹ and his⫹ markers were transferred to the
female with similar frequencies (Table 2). Subcloned between inversion-type and wild-type bacteria and on the frequency of return to wild-type order, we cannot immediately, such recombinants gave rise to two types
of colonies in roughly equal proportions: one type pro- estimate precisely the selective disadvantage due to a given inversion. In the following, the existence among moted bidirectional transfer like the original clone,
while the other transferred the chromosome in the same TcRrecombinants of an elevated ratio of clones displaying bidirectional transfer was taken as indicating that the inver-direction as the original Hfr ancestor. This suggested
that the triply resistant clones contained, at the time of sion of the tested segment was deleterious (R segments in Table 3).
the test, a mixed population composed of roughly equal
numbers of inversion-type and wild-type bacteria. South- The mosaic of permissive tolerant, permissive refrac-tory, and nonpermissive segments:TcR indigenous re-ern analyses performed on wild-type bacteria isolated
from mixed clones confirmed the view that these bacte- combinants between Tes and Tek insertions have always been detected, but their frequencies varied over two ria arose from a second exchange between the Tn10
sequences that restored a normal gene order and elimi- orders of magnitude. The TcR SmR KnRtriply resistant clones, among which inversion events are preferentially nated the initial inversion (data not shown). The mixed
populations might have arisen in either of two ways: observed, also occurred with variable frequencies among the TcRrecombinants (data not shown). A low either the inversion was highly unstable and reverted
at a high frequency to the wild-type genome order, or frequency of triply resistant clones, added to the fact that in general we avoided direct selection for the SmR the inversion was deleterious and rare wild-type
re-vertants overgrew the inversion strain more or less rap- character, explains why the number of candidate clones tested is low for some segments. In some cases, inter-idly. The second possibility was supported by analyses of
the rates of evolution of the recombinant populations. changing the Tes and Tek positions provided identical results (not shown). This is consistent with the idea that When mixed clones were incubated for longer periods
TABLE 3
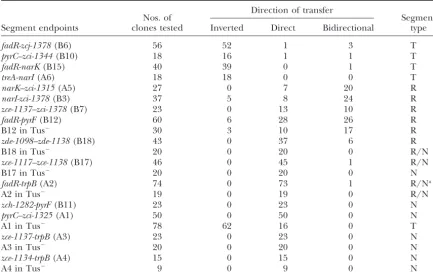
Conjugational transfer by triply resistant recombinant clones: examples of distribution and classification into different types
Direction of transfer
Nos. of Segment
Segment endpoints clones tested Inverted Direct Bidirectional type
fadR-zcj-1378(B6) 56 52 1 3 T
pyrC–zci-1344(B10) 18 16 1 1 T
fadR-narK(B15) 40 39 0 1 T
treA-narI(A6) 18 18 0 0 T
narK–zci-1315(A5) 27 0 7 20 R
narI-zci-1378(B3) 37 5 8 24 R
zce-1137–zci-1378(B7) 23 0 13 10 R
fadR-pyrF(B12) 60 6 28 26 R
B12 in Tus⫺ 30 3 10 17 R
zde-1098–zde-1138(B18) 43 0 37 6 R
B18 in Tus⫺ 20 0 20 0 R/N
zce-1117–zce-1138(B17) 46 0 45 1 R/N
B17 in Tus⫺ 20 0 20 0 N
fadR-trpB(A2) 74 0 73 1 R/Na
A2 in Tus⫺ 19 0 19 0 R/N
zch-1282-pyrF(B11) 23 0 23 0 N
pyrC–zci-1325(A1) 50 0 50 0 N
A1 in Tus⫺ 78 62 16 0 T
zce-1137-trpB(A3) 23 0 23 0 N
A3 in Tus⫺ 20 0 20 0 N
zce-1134-trpB(A4) 15 0 15 0 N
A4 in Tus⫺ 9 0 9 0 N
Segment endpoints are written Tes first; designations between parentheses refer to Figure 2. Triply resistant TcRSmRKnRrecombinants were assayed for transfer of PurE⫹and His⫹characters to the female strain LN1902 as described inmaterials and methods. With Hfr SV3 as an example, “direct” recombinant clones transferred, like their parent strains (TcS), thehis⫹markerⵑ50 times more frequently than thepurE⫹marker in the long-lasting (1 hr) conjugation assay used here. “Inverted” clones transferredpurE⫹at least 10 times more frequently thanhis⫹. “Bidirectional” clones provided an equivalent transfer ofhis⫹andpurE⫹markers (see Table 3).
aSegmentfadR-trp(A2) has been reclassified R on the basis of the analysis reported in Table 3.
to Tn10sequences, should not interfere with the expres- (bidirectional transfer). They constitute the major cate-gory. They are found all over the region analyzed, do sion of nearby genes.
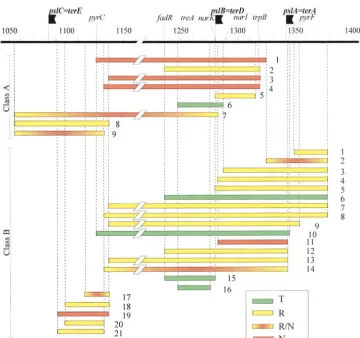
Results of inversion assays are summarized in Figure not necessarily overlap, and their sizes are variable (from 30 to 245 kb). They belong mostly to class B.
2, where segments are classified according to two
crite-ria: (i) the behavior in the inversion assay (T, R, or N), N segments:For five segments inversion was never de-tected. Some do not overlap, and their sizes vary from with reference to the examples presented in Table 3,
and (ii) the presence within the segment assayed ofpslB 39 to 200 kb. Three belong to class A, and those belong-ing to class B are among the smallest segments analyzed. and/orpslCbut not of pslApause sites (class A) or all
other situations (class B). The reason for considering R/N segments:Five segments, characterized by very few mixed clones and a majority of noninverted clones, were class A and B inversions separately is presented in the
next section. assigned to this ambiguous class. In the case of segment A2, the evolution analysis reported in Table 3 allowed
T segments:Five of the segments were tolerant to
in-version. They represent a novel feature of this region, an assignment to the R class.
Segment behavior in Tus⫺conditions:The present set since they were not observed in our previous analyses
(Rebolloet al. 1988;Francoiset al.1990a). Their sizes of Tn10insertions was exploited to complete our previ-ous finding that constraints to inversion exist indepen-range from 27 (fadR-narK) to 247 kb (zce1138-zcj1385).
They overlap or encompass many refractory or nonin- dently of the Tus/pause site system activity (Francois
et al.1990a). We chose to focus on segments classified vertible segments, and in some cases they share one
endpoint with refractory segments. Only one belongs A in Figure 3, since inversion ofpslBand/orpslCwithout inversion of pslA may hinder completion of chromo-to Class A (A6).
A3, and A4 was unchanged (they remained N or R/N), but that of segment A1 was dramatically altered: it shifted from N state (no inverted clone among 50 triply resistant colonies tested) to T state (80% inverted clones among 80 triply resistant clones). The behavior of B segments, either harboring pause sites (B12, R) or not (B17, R, and B18, N), was not affected by the⌬(tus)::ApR mutation.
Filament formation and abnormal nucleoids in bacte-ria harboring deleterious inversions: Filaments were routinely detected in mixed clones issuing from bacteria that had undergone a deleterious inversion. To charac-terize this phenotype, we examined the aspect of nucleoids in filaments by fluorescence microscopy after staining the DNA by DAPI. The analysis was not per-formed directly on the Hfr strains used above because we noted that bacteria carrying the repA(Ts) plasmid integrated in their chromosome tend to form filaments even when cultivated at a temperature nonpermissive for plasmid replication. Rather, it was carried out on inversion strains derived from Hfr strains that do not normally display filaments (Hfr’s KL19 and LN2085). The examinations also included F⫺bacteria, picked up from the smallest colonies found after selection for TcR SmRKnRcharacters. We expected such colonies to issue from a bacterium carrying a disabling inversion. Bacte-ria with abnormal morphology were easily detected for every strain in which the Tes and Tek insertions flanked an R segment. In all these strains, the abnormal cells (in general a few percent of the clone population) looked very similar, with filaments and abnormal distri-bution of nucleoids that remained grouped in elon-gated masses located at one or few positions in the
Figure3.—Predicted changes in chromosome replication
filaments (Figure 4, A–C, F). Note that the three R after pause site inversion. The simplified map shows only the
segments examined are not overlapping. For the Hfr positions of the polar pause sites (pslA,pslB, andpslC), oriented
as indicated in Figure 1. Vertical thin arrows indicate inversion strains, the bacteria carrying the inversion represented endpoints. Inverted segments are indicated by dotted lines. ⵑ30–70% of the mixed clones analyzed, as deduced Horizontal large arrows represent replication forks.(A)
Wild-from conjugation data. This frequency was much type organization: Replication forks travel through the region
greater than that of filaments. Abnormal bacteria could from left to right. The replication fork trap (RFT) is located
also be observed from time to time in strains having to the right ofpslA⫽TerA, and termination occurs normally
outside the region explored in the vicinity ofpsrA ⫽ TerC undergone inversion of a T segment but at a much (1607 kb;Louarn et al. 1994). (B) Class B inversion: The lower frequency than for R segments (an example is inverted segment includespslAbut notpslBorpslC. The arrest
provided in Figure 4E for segment A6). of the rightward moving fork at invertedpslAmust force
termi-nation to occur at this site but the overall replication time is
not dramatically increased. Ade novoRFT is generated. (C) DISCUSSION Class A inversion: The inverted segment includes pslB but
notpslA. The inverted pause site stops the rightward moving Experimental study of long-range interactions in the replication fork, and the leftward moving fork is blocked at bacterial chromosome is hampered by methodological pslA. Replication of the region between these terminators must
obstacles. Our approach has been to use precisely lo-be delayed.
cated genetic inversions to look for orientation-depen-dentcisinteractions between unknown elements remote from each other on the chromosome. It has recently carrying Tes and Tek insertions flanking segments A1
to A4 and B12, B17, and B18 as controls. Except for R proved its usefulness for understanding the regional control ondifactivity and for definition of the mecha-segments A2 and B12, they were classified N in Tus⫹
conditions (Table 3). TcRSmRKnRrecombinants of the nism generating thedif activity zone (DAZ; Peralset
al.2000), as well as for localizing the DNA regions re-Tus⫺derivatives were analyzed for presence of
Figure4.—Micrographs of bacteria undergoing inversions of T or R segments. Cultures of the strains indicated below were plated on L medium with tetracycline, streptomycin, and kanamycin. The smallest colonies arising after 24 hr of incubation at 37⬚were picked up, resuspended in 1 ml of LB, and incubated for 2–3 hr. Bacteria were then stained by DAPI and examined as indicated inmaterials and methods. For the two Hfr strains used, subsequent conjugation tests correlated the presence of abnormal bacteria with bidirectional transfer. (A) HfrKL19 aroA::Tes fadR::Tek (an R class A segment located between map positions 962 and 1236 kb;Rebolloet al.1988). (B) Hfr LN2085pyrF::TesfadR::Tek (R segment B12;Francoiset al.1990a). (C) CB0129pyrF::TesfadR::Tek (same R segment B12). (D) CB0129narK::TesfadR::Tes (T segment B15). (E) CB0129treA::Tes narI::Tek (T segment A6). (F) CB0129zci-1349::Teszcj-1378::Tek (R segment B1).
prespore during sporulation inBacillus subtilis( J.Err- A4, B12, B17, and B18, for instance, are R or N type in Tus⫹or Tus⫺ conditions, indicating that Tus was not ington, personal communication). Extended analyses
of inversions have also been performed in Salmonella involved in the behavior of these segments. The peculiar behavior of segment A1 can be related to similar
obser-typhimurium(SchmidandRoth1983;Segallet al.1988;
Mieselet al.1994), where examples of true noninverti- vations made independently inS. typhimurium:Miesel
et al. (1994) have observed segments that, although non-ble segments have been reported. The genetic system
used here allowed the generation of many conservative invertible by indigenous recombination, could give rise to well-tolerated inversions when the recombining se-inversions. We have analyzed the reaction of the cell to
inversion of 30 segments within the 23- to 29-min region quences consisted of exogenous DNA introduced by phage-mediated transduction. Eventually,J. Rothand to further explore a chromosome region (NDZ1)
pre-viously identified as refractory to inversion (Rebolloet co-workers (personal communication) discovered that the Tus protein was the factor inhibiting the indigenous
al. 1988;Francoiset al.1990a,b). The data confirm the
existence of constraints to inversion in this region since recombination required for inversion of these segments. In these closely related organisms, it appears that some most segments appeared either of N or R types and
illustrate a degree of complexity ofciseffects responsible chromosome segments are nonpermissive to inversion because the recombination event responsible for their for constraints to inversion in that (i) inversion of
cer-tain segments of this region is well tolerated and (ii) at formation aborts systematically for some as yet undeter-mined activity of Tus.
least one inversion is detectable in Tus⫺conditions only.
The fact that segment A1, the second longest of the At least inE. coli, the R or N character of many seg-ments is not determined by Tus activity. We focus in A series, shifted from the N state to the T state when
thetuslocus was deleted indicates that the Tus/pause the following only on segments for which inversion has been detected (T and R segments). The existence of T site system is one phenomenon determining segment
behavior in the inversion tests. However, smaller seg- segments encompassing R ones, plus the fact that many R segments do not overlap, make it unlikely that the ments A2 and A3, included in A1, remained N type in
Tus⫹as well as in Tus⫺conditions. Whether the inhibi- deleterious effects are due to reorientation of a single locus or to a change in direction of replication. Our tory effect of Tus on inversion of segment A1 directly
implicates the pause site it harbors is an open question. general interpretation is that NDZ1 is not a continuum but a succession of several smaller NDZs each harboring Since the smallest segments of the A series, A5 and A6,
may be inverted in Tus⫹conditions, it appears that (i) sequences whose relative orientation is physiologically important. Long T segments might contain entire a pause site present within the segment to be inverted
may not be an obstacle to the inversion event and (ii) NDZs, and the R segments might split and inactivate one of these NDZs, whereas small T segments might an inverted pause site is in itself not very detrimental,
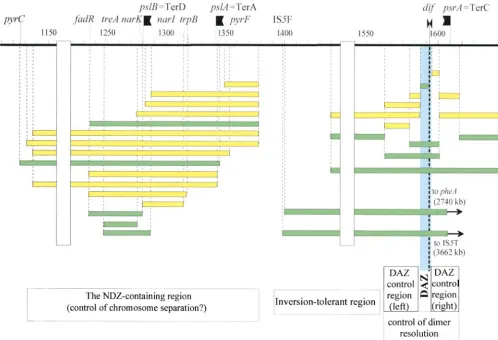
Figure5.—Distribution of tolerated and deleterious inversions in the terminus region. This map combines data redrawn from Figure 2 for T and R segments (between positions pyrC and zcj-1378) with previously published data to show the respective positions of the NDZ region, of the DAZ and the surrounding region involved in its formation, and of the seemingly inversion-tolerant intervening region. The large IS5F-IS5T inversion (Inv29-78) is described inLouarnet al.(1985), the large inversion from position 1397 kb topheAis described in Rebolloet al.(1988), and other inversions to the right of position 1430 kb are described inPerals et al.(2000). Green bars, tolerated inversions; yellow bars, deleterious inversions. Deleterious effects of inversions are probably different in the NDZ region (defects in chromosome separation?) and in thedif-controlling region (defects in dimer chromosome resolution).
NDZs in the 10-min region left ofdifleads to the pro- 1988), as if the rearranged chromosomes have difficulty in partitioning during the cell cycle. This suggests the posal that a given NDZ may be initiated and/or
termi-nated at specialized sequence elements. An NDZ might interesting novelty that the function of NDZs is to partic-ipate in spatial separation of sister chromosomes by include such initiator/terminator elements, plus
polar-ization factors. The nature of elements generating an facilitating or directing decatenase action. It may be that the NDZs revealed here all engage in a similar NDZ must for the moment remain uncertain, since the
present data do not allow precise determination of num- phenomenon and participate independently but addi-tively in nucleoid dynamics.
ber and positions of NDZs in the 23- to 30-min region.
The severity of growth defects due to deleterious in- A point of interest is the relation between the two phenomena typical of the terminus, the organization versions varies with the segments, but it is difficult to
estimate in the absence of precise information on fre- within NDZs and the generation of the DAZ. These phenomena are probably independent of one another. quencies of return to normal gene order. We suspect
that the growth defect is the consequence of a high risk The regions where NDZs are observed are peripheral to the terminus and do not cover the region governing of fatal problems leading to abnormal nucleoid and
cell morphology. The abnormal bacteria are probably dif activity: the map in Figure 5 shows that the NDZ region is separated from the DAZ-controlling region by destined to die, and the apparent generation time of
deleterious inversion strains is consequently increased. a zone (roughly between 1400 and 1565 kb on the map) that seems to tolerate inversions, even of very large seg-The altered nucleoid morphology closely resembles that
Neidhardt, R. Curtiss III, J. L. Ingraham, E. C. C. Lin, B. at IS5F; Louarn et al. 1985). Polarization of NDZs is
Magasaniket al.American Society for Microbiology, Washington, probably not achieved by the polar elements influencing DC.
Bird, R., J. Louarn, J. MartuscelliandL. Caro, 1972 Origin and
difactivity (Correet al.2000;Peralset al.2000), since
sequence of chromosome replication inEscherichia coli. J. Mol. these elements are not necessarily deleterious when
in-Biol.70:549–566.
verted: the long segment between positions 1440 and Capiaux, H., F. Cornet, J. Corre, M. Guijo, K. Perals et al., 2001 Polarization of theEscherichia coli chromosome: a view 1565 kb (Figure 5) is a T segment, although this segment
from the terminus. Biochimie (in press). harbors polar elements able to substitute for those
nor-Caro, L., andC. Berg, 1971 P1 transduction. Methods Enzymol. mally controllingdif(Peralset al.2000). However, the 21:444–458.
Corre, J., J. PatteandJ. M. Louarn, 2000 Prophage lambda in-alteration of cell morphology due to NDZ splitting, if
duces terminal recombination inEscherichia coli by inhibiting strongly reminiscent of that observed in Par⫺mutants,
chromosome dimer resolution: an orientation-dependentcis ef-is not very different from that due to def-istortion of the fect lending support to bipolarization of the terminus. Genetics
154:39–48. DAZ (Peralset al.2000).
De Massy, B., S. Bejar, J. Louarn, J.-M. LouarnandJ.-P. Bouche, Other possible leads to the nature of NDZs have
ap-1987 Inhibition of replication forks exiting the terminus region peared recently. First, the sequence of the region where of theE. colichromosome occurs at two loci separated by 5 min.
Proc. Natl. Acad. Sci. USA84:1758–1763. NDZs are found reveals a high potential for DNA
curva-Fellay, R., J. FreyandH. K. Krisch, 1987 Interposon mutagenesis ture and for low helix stability compared with the rest
of soil and water bacteria: a family of DNA fragments designed of the chromosome (Pedersenet al.2000). These prop- forin vitro insertional mutagenesis of Gram-negative bacteria.
Gene52:147–154. erties should be examined, in conjunction with skewed
Francois, V., J. Louarn, J. PatteandJ. M. Louarn, 1987 A system sequences, for their relationship to NDZ organization.
forin vivoselection of genomic rearrangements with predeter-Second, N. P.Higgins(personal communication) has mined endpoints inEscherichia coliusing modified Tn10
transpo-sons. Gene56:99–108. recently discovered that the ability of two res sites
in-Francois, V., J. Louarn, J. Patte, J. E. RebolloandJ. M. Louarn, serted in the terminus region of theS. typhimurium
chro-1990a Constraints in chromosomal inversions inEscherichia coli
mosome to recombine with each other depends upon are not explained by replication pausing at inverted terminator-like sequences. Mol. Microbiol.3:995–1002.
their relative locations, and he has located several
barri-Francois, V., J. Louarn, J. E. RebolloandJ. M. Louarn, 1990b ers hindering communication between the sites. Taking
Replication termination, nondivisible zones and structure of the into account the requirements for recombination be- Escherichia colichromosome, pp. 351–359 inThe Bacterial
Chromo-some, edited by K.Drlicaand M.Riley. American Society for tween res sites, he concluded that such barriers must
Microbiology, Washington, DC. behave as elements preventing supercoiling diffusion
Francois, V., A. ConterandJ. M. Louarn, 1990c Properties of new (Higginset al.1996), a property that would make them Escherichia coliHfr strains constructed by integration of
pSC101-derived conjugative plasmids. J. Bacteriol.172:1436–1440. good candidates for limits between NDZs.
Guyer, M. S., R. R. Reed, J. A. SteizandK. B. Low, 1980 Identifica-Finally, the Ter macrodomain, described recently by
tion of a sex factor affinity site inEscherichia colias gamma delta. Niki et al.(2000) and characterized by co-location in Cold Spring Harbor Symp. Quant. Biol.45:135–140.
Hashimoto-Goto, T., andM. Sekiguchi, 1977 Mutations to tem-the cell of all tested segments of a large region (20–30%
perature sensitivity in R plasmid pSC101. J. Bacteriol.131:405– of the chromosome) during the cell cycle, coincides
412.
well with the region where NDZs are found. One may Higgins, N. P., X. Yang, Q. FuandJ. R. Roth, 1996 Surveying a supercoil domain by using the gamma delta resolution system in speculate that the organization within NDZs involves
Salmonella typhimurium. J. Bacteriol.178:2825–2835. interactions between ordered sequences and some
scaf-Hill, T. M., 1996 Features of the chromosomal terminus region, pp. folding apparatus that not only participate in spatial 1602–1614 inEscherichia coli and Salmonella Cellular and Molecular
Biology, Ed. 2, edited by F. C.Neidhardt, R. Curtiss III, J. L. separation of sister nucleoids but also help compact this
Ingraham, E. C. C. Lin, B. Magasaniket al.American Society region of the chromosome. The behavior of the Ter
for Microbiology, Washington, DC.
domain in nucleoid dynamics might be due in part to Horiuchi, T., andY. Fujimura, 1995 Recombination rescue of the stalled replication fork: a model based on analysis of anEscherichia
the NDZ-dependent compaction and in part to the
DAZ-coli strain with a chromosome region difficult to replicate. J. associated mobilization of terminus DNA.
Bacteriol.177:783–791.
Kato, J.-I., Y. Nishimura, M. Yamada, H. SuzukiandY. Hirota, We are indebted to David Lane for numerous improvements to the
1988 Gene organization in the region containing a new gene manuscript. We thank Rafael Camacho and John Roth for discussions
involved in chromosome partition inEscherichia coli. J. Bacteriol. and sharing of unpublished information. We are grateful to Koryn
170:3967–3977. Perals for help with microscopy, and to Vincent Francois for pVF16
Konrad, E. B., 1977 Method for the isolation ofEscherichia coli mu-construction. This work has been supported by contract no.
SC1*-tants with enhanced recombination between chromosomal dupli-CT91-0713 of the Science Program of the European Community, cations. J. Bacteriol.130:167–172.
by contract CE92-0003 of the Direccion General de Investigacion Kuempel,P. L., J. M.Henson, L.Dircks, M.Tecklenburgand D. F. Cientifica y Te´cnica (Spain), and by the France-Spain “Mercure” Coop- Lim, 1991 dif, a recA-independent recombination site in the eration Program. terminus region ofEscherichia coli. New Biol.3:799–811.
Louarn, J., J. PatteandJ.-M. Louarn, 1982 Suppression ofE. coli dnaA46mutation by integration of plasmid R100.1 derivatives; constraints imposed by the replication terminus. J. Bacteriol.151:
657–667. LITERATURE CITED
Louarn, J.-M., J. P. Bouche, F. Legendre, J. LouarnandJ. Patte, Berlyn, M. K. B., K. B. Low, K. E. RuddandM. Singer, 1996 Link- 1985 Characterization and properties of very large inversions age map ofEscherichia coliK12, pp. 1715–1902 inEscherichia coli of theE. colichromosome along the origin-to-terminus axis. Mol.
Gen. Genet.201:467–476.
Louarn, J., F. Cornet, V. Francois, J. PatteandJ.-M. Louarn, 1994 possible role of two large, nondivisible zones in theEscherichia colichromosome. Proc. Natl. Acad. Sci. USA85:9391–9395. Hyperrecombination in the terminus region of the Escherichia
colichromosome: possible relation to nucleoid organization. J. Salzberg, S. T., A. J. Salzberg, A. R. KerlavageandJ. F. Tomb, 1998 Skewed oligomers and origins of replication. Gene217:
Bacteriol.176:7524–7531.
Messer, W., andC. Weigler, 1996 Initiation of chromosome repli- 57–67.
Sambrook, J., E. F. FritschandT. Maniatis, 1989 Molecular
Clon-cation, pp. 1579–1601 inEscherichia coli and Salmonella Cellular
and Molecular Biology, Ed. 2, edited by F. C.Neidhardt, R. Curtiss ing, Ed. 2. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
III, J. L. Ingraham, E. C. C. Lin, B. Magasaniket al.American
Society for Microbiology, Washington, DC. Schmid, M. B., andJ. R. Roth, 1983 Selection and endpoint distribu-tion of bacterial inversion mutadistribu-tions. Genetics105:539–557. Miesel, L., A. SegallandJ. R. Roth, 1994 Construction of
chromo-somal rearrangements inSalmonellaby transduction: inversions Segall, A., M. J. MahanandJ. R. Roth, 1988 Rearrangements of the bacterial chromosome: forbidden inversions. Science 241:
of non-permissive segments are not lethal. Genetics137:919–932.
Miller, J. H., 1992 A Short Course in Bacterial Genetics. Cold Spring 1314–1318.
Steiner, W. W., andP. L. Kuempel, 1998 Cell division is required Harbor Laboratory Press, Cold Spring Harbor, NY.
Niki, H., Y. YamaichiandS. Hiraga, 2000 Dynamic organization for resolution of dimer chromosomes at thediflocus of
Escheri-of chromosomal DNA inEscherichia coli.Genes Dev.14:212–223. chia coli. Mol. Microbiol.27:257–268.
Pedersen, A. G., L. J. Jensen, S. Brunak, H.-H. Staerfeldtand Steiner, W., G.Liu,W. D.Donachieand P.Kuempel, 1999 The D. W. Ussery, 2000 A DNA structural Atlas forEscherichia coli. cytoplasmic domain of FtsK is required for resolution of chromo-J. Mol. Biol.299:907–930. some dimers. Mol. Microbiol.31:579–583.
Perals, K., F. Cornet, Y. Merlet, I. DelonandJ.-M. Louarn, 2000 Way, J. C., M. A. Davis, D. Morisato, D. E. RobertsandN. Kleck-Functional polarization of theEscherichia colichromosome termi- ner, 1984 New Tn10 derivatives for transposon mutagenesis nus. Thedifsite acts in chromosome dimer resolution only when and for construction of lacZ operon fusions by transposition. located between long stretches of opposite polarity. Mol. Micro- Gene32:369–379.
biol.36:1–12.