DOI: 10.1534/genetics.104.030064
Interactions Between Imprinting Effects in the Mouse
Bruce M. Cattanach,
1Colin V. Beechey and Josephine Peters
Mammalian Genetics Unit, Medical Research Council, Oxfordshire OX11 0RD, United Kingdom Manuscript received April 7, 2004
Accepted for publication June 7, 2004
ABSTRACT
Mice with uniparental partial or complete disomies for any one of 11 identified chromosomes show abnormal phenotypes. The abnormalities, or imprinting effects, can be attributable to an incorrect dosage of maternal or paternal copies of imprinted gene(s) located within the regions involved. Here we show that combinations of partial disomies may result in interactions between imprinting effects that seemingly independently affect fetal and/or placental growth in different ways or modify neonatal and postnatal imprinting effects. Candidate genes within the regions have been identified. The findings are generally in accord with the “conflict hypothesis” for the evolution of genomic imprinting but do not clearly demonstrate common growth axes within which imprinted genes may interact. Instead, it would seem that any gene that represses or limits embryonic/fetal growth to the advantage of the mother—by any developmental means—will have been subject to evolutionary selection for paternal allele repression. Likewise, any gene that favors embryonic/fetal development at consequent cost to the mother—by any developmental means—will have faced selection for maternal allele repression. The classicalIgf2-Igf2raxis may therefore be unique. The findings involve reinterpretation of older imprinting data and consequently revision of the mouse imprinting map.
T
HE discovery that certain chromosomal regions can neonatal abnormalities and inviabilities, prenatal fetal lead to developmental abnormalities when both and placental growth effects, and postnatal growth retar-copies are exclusively maternally or paternally derived dations sometimes with other subtle defects ( Catta-(Cattanach and Kirk 1985) has contributed signifi- nachandBeechey1997). A general feature of all these cantly to the recognition and understanding of the phe- effects has been their lack of variation, irrespective ofnomenon of genomic imprinting. The tools for this genetic background or translocation used. However,
approach have been the uniparental whole chromo- there have been clear exceptions. Here, we reevaluate some disomies and uniparental partial chromosome di- these exceptions and, together with new data, show that somies (conventionally called maternal and paternal the cause is the co-occurrence of imprinting effects at-duplications) that can readily be generated using Robert- tributable to both chromosomes involved in the translo-sonian and reciprocal chromosome translocations, re- cations used. This poses questions of interest in relation spectively. Application, step by step, to the whole ge- toMooreandHaig’s (1991) “conflict hypothesis” for nome has identified 11 regions (imprinting regions) the role of imprinting. According to the hypothesis, distributed over seven different chromosomes in which maternally expressed genes favor the well-being of the developmental abnormalities occur when two copies mother over the fetus, while paternally expressed genes from one parent and none from the other are present. favor the development of the fetus at the expense of Both parental copies are therefore clearly required for the mother, providing a reason why both paternal and normal development (CattanachandKirk1985;Cat- maternal alleles are required. In the situations to be tanachandBeechey1997). Over the years the size of described, two maternally expressed alleles at one im-the imprinting regions has been progressively reduced printed gene locus are set against two paternally ex-through the use of new translocations and, with few excep- pressed alleles at another. The question is then whether tions, all of the uniparentally expressed imprinted genes this will cause further problems in development or rees-have located within these reduced regions (Beechey et tablish a balance. The combination of maternal
duplica-al.2004). tion for one region with paternal duplication for
an-The developmental abnormalities identified have other therefore offers the possibility of ascertaining been diverse. Described as imprinting effects, these have whether the genes involved operate on the same or ranged from early embryonic to late fetal lethalities, different growth axes, as illustrated for the paternally
expressed Ig f 2-maternally expressed Ig f 2r interaction (Filsonet al.1993). Interpretation is difficult because 1Corresponding author:MRC Mammalian Genetics Unit, Harwell,
Did-it is often uncertain which genes are involved and how
cot, Oxfordshire OX11 0RD, United Kingdom.
E-mail: [email protected] they affect development. The findings suggest that
tor, Burlingame, CA). Images were captured using a
fluores-mic imprinting is a highly complex process involving
num-cent microscope with SmartCapture 2 software (Digital
Scien-bers of different growth axes that operate differently in
tific, UK) and merged with captured images of the same cells
different tissues and at different times in development. in which chromosomes were DAPI stained. In all cells the The “conflict hypothesis” provides a basis for many of DAPI staining was reversed and enhanced by the software to
produce reversed DAPI banding (or pseudo-G-bands) that
these imprinting effects, the multiplicity suggesting a
allowed precise location of the signal on the mouse ideogram.
continuity in the evolutionary adaptation of imprinting
Mapping and PCR expression studies withRasgrf1:Total
as a means of favoring fetal growth by paternal genes
RNA was isolated from the cerebellum from two sets of adult
and its negative control by maternal genes. The reinter- MatDp(dist9) mice generated with T(2;9)11H and T(4;9)45H pretations of the imprinting regions and effects require and also from wild-type siblings. RNA samples were prepared
using RNAzol B (Biotechnology Europe) and 1 g of total
changes to the mouse imprinting map (Beechey et al.
RNA per sample was reverse transcribed with M-MLV reverse
2004).
transcriptase (Life Technologies) using oligo (dT)15
(Pro-This article describes interactions involving proximal
mega, Madison, WI).Rasgrf1cDNA was amplified using
for-chromosome 2 (Chr 2) with proximal Chr 11, proximal ward primer CDCFO1 and reverse primer CDCRO2, as described Chr 7 with central Chr 7, proximal Chr 7 with proxi- byPlass et al.(1996). AsRasgrf1 is expressed predominantly
from the paternal allele in cerebellum, little or no expression
mal Chr 11, and distal Chr 2 with distal Chr 9.
would be expected in the MatDp(dist2) samples if the gene lies distal to the breakpoints. If the gene lies proximal to the breakpoints, expression would be detected.
MATERIALS AND METHODS
Mouse breeding:The imprinting effects studied were
inves-RESULTS tigated in mice with maternal and paternal duplication
(MatDp and PatDp; see standard nomenclature rules,Mouse Proximal Chr 2-Proximal Chr 11 interaction Genome Database 2004) for selected regions of
chromo-somes and in the case of chromosome 11 (Chr 11) also with Background: Mice with two maternal copies of Chr uniparental whole chromosome disomies (MatDi and PatDi).
11, MatDi(11), and no paternal copies show prenatal
The duplication classes were generated by the standard genetic
growth retardation and have small placentas. Although
method of intercrossing genetically marked and unmarked
trans-small at birth, they are generally fully viable, and
postna-location heterozygotes and identifying the MatDp and PatDp
classes with the aid of the markers (CattanachandBeechey tal growth is normal such that they remain small through
1997). A vital consideration in the present studies is that with to adulthood. The reciprocal genotype, PatDi(11), reciprocal translocations two chromosomes are of necessity shows enhanced fetal and placental growth. They are always involved and therefore, in creating animals with, for
large at birth and fully viable through to adulthood
example, MatDp for a region of one chromosome, these
ani-when they are still seen as oversized (Figure 2A; Catta-mals also have PatDp for the corresponding region for the
other (Figure 1). nachandKirk 1985;Cattanachet al.1996).
Translocation and marker gene stocks, with one exception, The MatDi(11) growth retardation is also found with are maintained on a mixed C3H/HeH-101/H genetic back- two maternal copies of the region proximal to the ground. The exception comprised the marker gene stock for
T(2;11)30H breakpoint in Chr 11,
MatDp(prox11)/Pat-distal Chr 2, which was maintained on a mixed C57BL/6J-JU/
Dp(prox2), thereby identifying the region of Chr 11
Ct-⫹c ⫹a genetic background. The translocations used are
identified according to standard nomenclature rules such that where the gene(s) responsible for the growth effects
the chromosomes involved are identified in brackets and the must lie. However, no progeny representing the PatDp laboratory of origin specified in the symbol following. Thus,
(prox11)/MatDp(prox2) class, which was expected to
T(2;11)30H indicates that this translocation involves Chrs 2
show the Chr 11-enhanced prenatal growth effect, were
and 11 and that this was the thirtieth translocation named
found in the reciprocal T30H cross. Further studies
by Harwell. Throughout the text, these symbols are reduced
wherever possible to T30H, etc. The Robertsonian transloca- indeed indicated that the class could not even be
de-tion used to generate uniparental Chr 11 disomies was Rb tected prenatally, suggesting that maternal Chr 2 proxi-(11.13)4Bnr, or Rb4Bnr. The marker genes used in the various mal to the T30H breakpoint is also subject to imprinting. crosses are specified in the figure legends. TheOed-Sml
muta-On this basis MatDp(prox2) appeared to cause an early
tion (Cattanachet al.2000) used in the distal Chr 2 studies
embryonic lethality (CattanachandKirk1985). With
was also maintained on the mixed C3H/HeH-101/H
back-ground. the use of a series of Chr 2 translocations with yet more
All animal studies were carried out under the guidance of proximal breakpoints (Figure 3A) and screening for the Medical Research Council in “Responsibility in the Use of known distal Chr 2 imprinting effects (to be described Animals for Medical Research” ( July 1993) and Home Office
later) at birth, the size of the proximal region was
pro-Project license nos. 30/1518 and 30/2065.
gressively reduced (Cattanach1986). The region
re-FISH mapping of Gatm on Chr 2: Mitotic chromosome
spreads from the bone marrow of a T(2;9)11H heterozygote sponsible for the MatDp(prox2) imprinting lethality was
were prepared by standard methods. These were used for finally defined as that lying above the most proximal FISH with bacterial artificial chromosomes RP23-101F14 and translocation (T68H) breakpoint and thus very close to RP-23-99H13 (UK MRC Mouse Genome Sequencing
Pro-the centromere (Figure 3A). No imprinted genes have
gram), each of which contains theGatmgene, by a modified
been found in this region.
method of Buckle and Rack (1993). Hybridization signals
Figure1.—System for generating uniparental partial disomies (MatDp and PatDp) using recip-rocal translocations. In the example shown the translocation is T(2;11)30H, involving Chrs 2 and 11, and the screen is for MatDp and PatDp for the regions proximal to the breakpoints in the two chromosomes. The region of Chr 11 proximal to the breakpoint in the male parent is marked withwa2.Complementary adjacent-2 disjunction at meiosis (homologous centromeres going to the same pole) in both parents can gener-ate chromosomally balanced offspring showing the recessivewa2phenotype. These will be geneti-cally PatDp for the proximal region of Chr 11 and, simultaneously, MatDp for the proximal region of Chr 2, MatDp(prox2)/PatDp(prox11). Unmarked PatDp(prox2)/MatDp(prox11) young will also be produced. In the reciprocal cross (not shown), withwa2on the maternal Chr 11, the wa2marker will identify the PatDp(prox2)/Mat-Dp(prox11) class. Any marker gene located proximally on either chromosome can be used. With adjacent-1 disjunction (homologous cen-tromeres going to opposite poles) in both par-ents, offspring with MatDp(dist2)/PatDp(dist11) and PatDp(dist2)/MatDp(dist11) can be similarly generated. These can be recognized in recipro-cal crosses with the aid of markers that are lo-cated distal to the breakpoints on one or the other chromosome.
T(5;11)57H and then with T(11;13)41Ad, in neither of detectable at birth by growth retardation and was found to be fully viable through to adulthood.
which is the second component chromosome (Chrs 5
or 13) imprinted, has reduced the imprinting region Reinvestigations with T30H:To establish more firmly that MatDp(prox2) results in an early lethal imprinting to a segment close to the centromere (Cattanachet
al.1998; Figure 2B). Significantly, this smaller region effect when generated with the T30H translocation, a repeat prenatal screen was initiated. Among 269 em-encompasses the loci of the two known Chr 11
im-printed genes,Gbr10andU2af1-rsl(Monket al.2003). bryos checked, none represented the MatDp(prox2) class. The zero incidence from the total T30H data The latter has not been found to affect growth (
Suna-haraet al.2000) but aGbr10knockout has been found differs significantly from the combined MatDp(prox2) frequency obtained with the T26H, T1Go, and T2Wa to cause a fetal and placental overgrowth when
mater-nally transmitted (Charalambouset al.2003). As a ma- translocations both prenatally and at birth (Table 1), verifying that two distinct MatDp(prox2) imprinting ternally expressed growth suppressor,Gbr10is the likely
candidate gene for the imprinting effects, two expressed phenotypes appear to be produced.
All the translocation crosses were carried out on the maternal copies suppressing growth with MatDp
(prox11) and an absence of an expressed copy resulting same genetic background. Genetic background effects cannot therefore be responsible for the differing im-in overgrowth with PatDp(prox11).
Chr 2: With Chr 2, new work with three different printing effects. None of the new translocations involve Chr 11, however. Therefore, a more feasible explana-translocations, T(2;8)26H, T(2;4)1Go, and T(2;8)2Wa,
the breakpoints of which all lie close to that of T30H tion of the findings is that the fetal/placental growth retardation with normal postnatal viability characterizes (Figure 3B), have provided reason to reinterpret the
conclusions on the proximal imprinting effect and the the MatDp(prox2) imprinting effect and that the early embryonic lethality generated only with T30H derives location of the region. In none of these new
transloca-tions is the second chromosome involved (Chrs 4 or 8) from an interaction between the MatDp(prox2) and PatDp(prox11) imprinting effects.
imprinted, and none generated the MatDp(prox2) early
embryonic imprinting lethality detected with T(2;11)30H PatDp(prox2) imprinting phenotype: In addition to the MatDp(prox2) findings, the work with the three (Table 1). Instead, a different MatDp(prox2) imprinting
TABLE 1
In uteroand birth recovery of MapDp(prox2) and Patdp(prox2) classes generated with different Chr 2 translocations
MatDp(prox2) PatDp(prox2)
Translocation In utero At birth In utero At birth
T(2;11)30H 0/473 (0%) 0/714 (0%) 1/63 (1.6%) 8/714 (1.1%) T(2;8)2Wa 13/742 (1.7%) 1/178 (0.6%) 3/210 (1.4%) 1/157 (0.6%) T(2;8)26H 3/476 (0.6%) 13/2012 (0.6%) 33/365 (0.8%) 17/1542 (1.1%) T(2;6)1Go 1/70 (1.4%) 3/716 (0.4%) 4/292 (1.4%) 8/340 (2.3%)
The duplication classes were recognized at 16.5–18.5 days of gestation and at birth using the pallid (pa) eye-color marker (Figure 3, A and B). The zero incidence with the T30H translocation was significantly different from the pooled incidencesin utero(P⫽0.010) and at birth (P⫽0.034) obtained with the T2Wa, T26H, and T1Go translocations.
(prox2) class. Fetal growth was not affected but there readily be noted in the presence of the abnormal pheno-type attributable to a MatDp(dist2) imprinting effect (to was a marked overgrowth of the placenta (Table 2;
Fig-ure 3B), thereby breaking the normal relationship be- be described). As a consequence, the proximal region could be enlarged to comprise the majority of the chro-tween fetal and placental size. The placental effect has
since also been seen in limited studies with T30H. Thus, mosome, extending from the centromere down to the distally located T26H breakpoint (Figure 3B). However, among 63 fetuses screened, 2 PatDp(prox2)/MatDp
(prox11) showed the expected growth retardation at- newer prenatal studies using T11H have shown that
MatDp and PatDp for the region distal to the breakpoint tributable to MatDp(prox11) imprinting (mean 1.23 g
cf. 1.72 g for their normal sibs; 71%) and the PatDp express only the distal Chr 2 imprinting phenotypes and not the small embryo/placenta and large placenta (prox2) overgrowth of their placentas relative to those
of their normal sibs (0.158 gcf. 0.094 g; 162%). Thus, effects. The region, although still large, is therefore likely to lie proximal to T11H (Figure 3B). It is therefore the combination of Chr 2 and 11 imprinting effects
enhances the fetal-placental growth discordance, with of significance that the recently discovered imprinted gene, Gatm, which is maternally expressed in mouse the PatDp(prox2) overgrowth in the placenta
overrid-ing the MatDp(prox11) placental growth reduction. Fe- placenta and postulated to be a growth suppressor ( San-dellet al. 2003), maps close to the T11H breakpoint and tal size was clearly not influenced by the larger placental
size. therefore potentially within this newly defined proximal
Chr 2 imprinting region. This location has been
con-Changes in the proximal Chr 2 imprinting region and
candidate gene:The new interpretation of the data in- firmed by FISH analysis with BACs in which all mitoses showed hybridization with Gatm on both the normal validates the early refinements of the MatDp(prox2)
imprinting region. The small size at birth would not Chr 2 and the large rearranged Chr 29(data not shown).
TABLE 2
Effects of MatDp(prox2) and PatDp(prox2) generated with different translocations on fetal and placental weights
Ratios of weights of Ratios of weights of embryos to those placentas to those Genotype Translocation of normal sibs of normal sibs
MatDp T2Wa 0.762⫾0.026 0.626⫾0.059
T26H 0.882⫾0.057 0.702⫾0.122
T1Go 0.711⫾0.079 0.704⫾0.217
All 0.782⫾0.023 0.647⫾0.048
PatDp T2Wa 0.997⫾0.124 1.449⫾0.501
T26H 0.992⫾0.143 1.665⫾0.664
T1Go 1.076⫾0.076 1.614⫾0.297
All 1.046⫾0.059 1.586⫾0.234
Gatmis therefore the candidate gene for the placental study, however, is the expectation that the growth retar-growth effects, two expressed copies suppressing placen- dation/lethality seen with the T9H and T50H transloca-tal growth with MatDp (prox2) and an absence of an tions (SearleandBeechey1990) could be a combined expressed copy resulting in the placental overgrowth phenotype derived from proximal and central Chr 7 with PatDp(prox2). An interaction betweenGatm(Chr imprinting effects.
2) andGrb10(Chr 11) effects may account for the Chr In addition to the MatDp findings, PatDp for the
2-Chr 11 interaction seen with T30H. region proximal to the T9H breakpoint generated a
mouse model of Angelman syndrome (Cattanach et
Conclusions:
al.1997; Figure 4A), the phenotype including abnormal 1. The imprinting effect found with MatDp for the brain wave patterns similar to those of AS patients. For proximal region of Chr 2 no longer comprises an technical reasons (B. M.Cattanach, unpublished re-early embryonic lethality but rather a fetal and pla- sults), it has proven impossible to investigate PatDp for
cental growth retardation. the central region using the IsCt insertional
transloca-2. A new imprinting effect, comprising a placental over- tion.
growth, has been found associated with PatDp for New data: Refinement of the proximal imprinting region:
the region. Studies using two further translocations, T(7;13)43Ad
3. The region now comprises a large part of Chr 2 and T(5;7)30Ad, which have breakpoints proximal to
extending down to the T11H breakpoint. the PWS/AS homologous region, have shown that
4. The lethality observed with MatDp(prox2)/PatDp MatDp for the regions proximal to both breakpoints
(prox11) using the T30H translocation can be de- (Figure 4B) does indeed exhibit a different imprinting duced to result from an interaction between the effect from that seen with T(7;15)9H and T(7;18)50H.
proximal Chr 2 and proximal Chr 11 imprinting In none of these translocations is the second
chromo-effects. some involved subject to imprinting. Only Chr 7
there-5. The imprinted gene,Gatm, which is maternally ex- fore needs to be considered. The new imprinting pheno-pressed in the placenta, lies within this extended type comprised a prenatal growth retardation effect, as
region and becomes the candidate gene for the pla- seen with T9H and T50H, but without the neonatal
cental imprinting growth effects. lethal component. Thus, of 37 MatDp(prox7) mice
gen-6. Gbr10 is the candidate locus for the proximal Chr erated with T40Ad and detected at birth, all but 2
sur-11 imprinting effects. vived to adulthood. This seeming discord supports the
7. The interaction between the Chr 2 and Chr 11 im- contention that the T9H and T50H neonatal lethality printing effects may therefore involve theGatmand derives from the cumulative effects of the MatDp
Gbr10imprinted gene loci. (prox7) Chr 7 fetal growth retardation, seen with T43Ad
and T30Ad, and the MatDp(cen7) PWS early postnatal
Proximal Chr 7-Central Chr 7 interaction lethality.
Candidate genes: Clusters of imprinted genes have
Background:The imprinting of Chr 7 (Figure 4A) is
been identified for both regions. Peg3, which maps complex insofar as there are at least three regions that
within the proximal cluster close to the centromere are subject to imprinting: (a) a proximal region (Searle
(Monket al.2003), is the likely candidate for the growth-andBeechey1990), (b) a central region (Cattanach
promoting effect as the gene is expressed in the embryo et al.1992), and (c) the well-studied distal region
encom-and placenta encom-and the paternally inherited knockout passing the H19-Igf2 imprinted gene cluster (Searle
shows a fetal growth retardation (Li et al.1999). The andBeechey 1990;Beechey et al.2004).
closely associated Zim1 and Usp29 are other possible Mice with two maternal copies and no paternal copies
candidate genes (Kimet al.1999;Liet al.1999) but are of the regions proximal to the T(7;18)50H and
not known to affect fetal growth. On the basis of work T(7;15)9H breakpoints on Chr 7, MatDp(prox7), show
on the knockout, Ndn is the likely candidate for PWS a fetal and placental growth retardation with neonatal
in the mouse and elements of the disorder in humans lethality (SearleandBeechey1990; Figure 4A). With
(Renet al.2003). the Is(7;X)Ct insertional translocation, which allows a
Absence of a PatDp(prox7) imprinting effect: The central region of the chromosome to be studied
sepa-T43Ad and T30Ad studies also showed that PatDp for rately (Cattanach et al. 1992), a distinctly different
the regions proximal to their breakpoints was not associ-postnatal lethality occurs with MatDp for the small
re-ated with an imprinting effect and notably no compo-gion that lies between the T9H breakpoint and the more
nent of the recognized AS-type imprinting effect ( Cat-proximal IsCt breakpoint (Figure 4A). As this region is
tanach et al. 1997). This, therefore, further refines homologous to the Prader-Willi/Angelman syndrome
the position of the central AS imprinting region on the (PWS/AS) imprinting region of human Chr 15, the
paternal chromosome to the small region between the postnatal effect has been taken as a mouse model of
mologous region lies in this segment of mouse Chr 7 these mice at 3 weeks of age appeared to be reduced, but normal size was attained by adulthood (Figure 5B). (Figure 4B).
As previously discussed,Peg3may be regarded as
candi-Conclusions: date gene for the proximal Chr 7 growth retardation
withGrb10 remaining the candidate gene for the Chr 1. The proximal Chr 7 imprinting effect found with
11 growth effect. An interaction between thePeg3and MatDp now consists of a fetal and placental growth
Grb10imprinting effects might therefore be indicated. retardation with full viability.
MatDp(prox11)/PatDp(prox7): Without a PatDp 2. The region is restricted to the segment proximal to
(prox7) imprinting effect for the region proximal to the the T43Ad breakpoint.
T43Ad breakpoint (Figure 4B), the MatDp(prox11)/ 3. The lethal imprinting effect found with MatDp for
PatDp(prox7) combination presented only the expected the larger T9H-centromere region can be attributed
small MatDp(prox11) fetal/placental imprinting effect to the combination of the proximal and central
im-(Cattanach and Kirk 1985; Figure 3B). The growth printing effects.
retardation of this class was evident at birth, as usual, and 4. Peg3is the candidate gene for the imprinting effect
maintained through to adulthood (data not shown). of proximal growth retardation.Ndnmay be the
can-didate gene for PWS in the mouse. Conclusions:
5. PatDp for the region proximal to the T30Ad
break-point does not give an imprinting effect. It can be 1. On the basis of the placental effect, the proximal Chr 7 imprinting region can now be reduced to a concluded that, as expected, the mouse AS effect is
attributable to the more distal region between the small region very close to the centromere.
2. The PatDp(prox11) imprinting effect potentially at-T30Ad and T9H breakpoints, which encompasses the
PWS/AS homologous region. tributable to Grb10 appears to operate primarily
upon early fetal development, as previously sug-gested (Cattanachet al.1992).
3. The MatDp(prox7) imprinting effect appears to
op-Proximal Chr 7-op-Proximal Chr 11 interaction
erate primarily upon the placenta, as might be
ex-Background: As described above, the proximal Chr pected ifPeg3is the responsible gene.
11 imprinting effects comprise a fetal and placental 4. Both the fetal and placental effects appeared
transi-growth retardation with MatDp for the region proximal tory.
to the T41Ad breakpoint and a fetal and placental over- 5. The interactions between the proximal Chr 7 and growth with PatDp for the region (Figure 2B). A proxi- proximal Chr 11 imprinting effects may be attribut-mal Chr 7 imprinting effect is found with MatDp for able to theGrb10andPeg3imprinted gene loci. the region proximal to the T43Ad breakpoint (Figure
4B), comprising a fetal and placental growth retardation (see previous section).
Distal Chr 2-Distal Chr 9 interaction New data: T(7;11)40Ad: Evidence of an interaction
between the proximal Chr 7 and 11 imprinting effects Background:The distal Chr 2 imprinting region first detected in the T30H studies (Cattanach and Kirk was made with a new translocation, T(7;11)40Ad. The
initial objective of this study was to distinguish whether 1985) yields two distinct imprinting effects. MatDp (dist2) newborns are seen as flat-sided, hump-backed, the proximal Chr 7 imprinting effect was attributable
to regions proximal or distal to the translocation and hypoactive. They fail to suckle and die within a few hours of birth. The reciprocal PatDp class typically breakpoint, which lies very close to the centromere
(Fig-ure 4B). The T40Ad breakpoint in Chr 11 is located exhibit short, square bodies, edema, paddle feet, kinked tails, hyperactive behavior, and, as demonstrated by all fairly distally (Figure 2B) such that studies upon the
region proximal to the breakpoint clearly include the but one of a series of Chr 2 translocations (Cattanach 1986), invariably dies within a few days of birth. Through proximal Chr 11 imprinting region.
MatDp(prox7)-PatDp(prox11) interaction: MatDp the use of these Chr 2 translocations the imprinting region was reduced to the region lying between the (prox7)/PatDp(prox11) fetuses generated with the
T40Ad translocation were found to be significantly T2Wa and T28H breakpoints (Figure 3A), limiting its size toⵑ7 Mb (E. P.Evans, personal communication). larger than their normal sibs (Figure 5A) at 14.5 and
15.5 days, thus exhibiting the expected PatDp(prox11) This encompasses the Gnascluster (Williamson et al. 1996), and a new nonfunctional mutation detected in imprinting effect on fetal growth (CattanachandKirk
1985). By contrast, placental sizes were significantly re- Gnas(Cattanachet al.2000;Skinneret al.2002; Wil-liamson et al. 2004) has been found to produce an duced (Figure 5A), apparently showing the MatDp
(prox7) imprinting effect in this tissue. Both effects edematous phenotype, that, when maternally inherited, closely resembles that of the PatDp class, although with-were transitory as at a later age both fetal size and
Figure5.—Fetal and placental growth anomalies attributable to an interaction between MatDp (prox7) and PatDp(prox11) im-printing effects; ratios relative to normal sibs. (A) Fetal weights were significantly larger than those of normal sibs, averaged over all litters, at 14.5 and 15.5 days (P⫽0.010 and⬍0.000001, respectively) but not at 16.5 days (P ⫽ 0.974). Placental weights were significantly smaller than those of normal sibs, averaged over all litters, at 14.5 and 15.5 days (P⫽0.0098 and 0.0012, re-spectively) but not at 16.5 days (P⫽0.56). (B) Postnatal weights were not significantly different from those of normal sibs at birth, 1 week, and 6 weeks (P⫽ 0.313, 0.60, and 0.93, respec-tively) but were significantly dif-ferent at 3 weeks (P⫽0.0019). Overall, there was no significant difference between weight ratios at all ages (P⫽0.012).
that the absence of maternally expressedGnasis largely dence has been found that would suggest that genetic background differences are responsible (Cattanach responsible for the PatDp phenotype.
The one exceptional PatDp class lacks the behavioral 1991a,b) and as the size of the duplicated region is larger rather than smaller than that generated with most effect and is semiviable rather than lethal (Cattanach
evi-Figure6.—Imprinting map of Chr 9. New map showing the location of the imprinting region distal to the T45H breakpoint and the Chr 2–Chr 9 interaction (in italics) obtained us-ing the T11H translocation. The Chr 2 marker gene,bp, was used in the T11H studies; the Chr 9 genes,dand ln, in the T138Ca studies; the Chr 4 gene,b, in the T45H studies; and the Chr 1 gene,ln, in the T27H studies. The latter, for a reason as yet un-known, did not distinguish the distal Chr 9 imprinting effect. The T11H breakpoint has been reassigned to a position close to that of T45H. Pre-viously, it had been located more proximally on the chromosome.
sponsible. The second chromosome involved in the of the T138Ca and T45H MatDp growth retardations
(Figure 7). This raises the possibility that the loss of the T11H translocation, Chr 9, was initially not recognized
as being imprinted and potentially involved. behavioral effect and increased viability of the PatDp
(dist2) class generated with the T(2;9)11H translocation
New data: Evidence that distal Chr 9 is imprinted:
The translocation evidence on Chr 9 imprinting (Figure is attributable to an interaction between the distal Chr 2 (Gnascomplex) and distal Chr 9 (Rasgrf1) imprinting 6) has been ambiguous. A postnatal growth retardation
was initially found in MatDp(dist9) mice generated with effects. There is another Chr 9 imprinted gene, A19, which maps close toRasgrf(de la Puenteet al.2002). T(9;17)138Ca but this was not evident with another Chr
9 translocation, T(1;9)27H (CattanachandBeechey However, as it is a noncoding gene, it is unlikely to be involved in any interaction with the Chr 2 imprinting 1994), suggesting that Chr 17 [PatDp(dist17)] was
re-sponsible for the growth effect. This conclusion proved effect.
Evidence of a Rasgrf1-PatDp(dist2) interaction:The erroneous, however, because the growth effect was
subse-quently found with a third Chr 9 translocation, T(4;9)45H above T11H PCR expression studies confirmed that Rasgrf1is not expressed in T11H MatDp(dist9)/PatDp (Figure 7), suggesting that the Chr 9 genotype, MatDp
(dist9), was in fact responsible. Consistent with this con- (dist2) mice. However, to ascertain if it is the absence ofRasgrf1that is responsible for the modified behavior clusion is the finding that the growth curves of the
T138Ca and T45H MatDp(dist9) classes accord well and viability, MatDp(dist4)/PatDp(dist2) mice
gener-ated with T26H were investiggener-ated in crosses in which a (Figure 7) with that of a paternally inherited knockout
of the imprinted paternally expressed Chr 9 gene, null mutation of Rasgrf1 (Clapcott et al. 2003) was segregating. Those that did not inherit the mutation Rasgrf1(Itieret al.1998), and also with that of a
pater-nally inherited mutation at the locus (Clapcott et al. from the father showed evidence of the typical PatDp (dist2) hyperactive behavior from birth and none sur-2003). Rasgrf1is paternally expressed in the neonatal
and adult brain (Plasset al.1996). Therefore, to con- vived for more than 4 days (n ⫽ 14). However, 14 of 27 that carried the paternalRasgrf1mutation, and which firm that the gene lies distally on the chromosome
(Pearsall et al. 1998) and could be responsible for therefore did not express the gene, showed little sign of the behavioral effect and survived for at least 10 days. the MatDp(dist9) growth retardations, PCR expression
studies were conducted on cerebellum samples from The difference is statistically significant (P⫽0.00057). Furthermore, uniquely for T26H-generated PatDp MatDp(dist9) mice generated with T11H and T45H
(Figure 6). No evidence of paternal allele expression (dist2) animals, some survived to adulthood. An addi-tional finding was that, at birth, the incidence of the was found, whereas expression was found in the controls
Figure 7.—Postnatal growth curves of MatDp(dist9) mice gen-erated with T45H and T138Ca and those with male and female pater-nally inheritedRasgrf1knockouts; ratios relative to normal sibs. Knockout data are from Itier et al.(1998).
(dist2) prenatal viability is also enhanced in the absence between Chr 2 (Gnas) and Chr 9 (Rasgrf1) imprinting effects.
ofRasgrf1. The segregation of theRasgrf1mutation was
5. Specifically, the modification can be attributed to normal among 10 normal sibs studied andRasgrf1
segre-the absence ofRasgrf1expression in these animals. gates normally in other crosses (Clapcottet al.2003).
Finally, to determine if an absence ofRasgrf1also modi-fied the phenotype of the maternally inheritedOed-Sml
mutation at the Gnas locus (Cattanach et al. 2000), DISCUSSION
the same test system was applied. Mice showing the
Changes in the mouse imprinting map:The data pre-maternally inheritedOedphenotype do not show a
be-sented modify the imprinting map of the mouse as last havioral effect like PatDp(dist2), but nevertheless few
formally published (Cattanach and Beechey 1994)
survive more than a few days. The absence of Rasgrf1
and as currently reported on the Harwell website expression was not found to modify the viability or any
(Beechey et al. 2004). Thus, (1) the proximal Chr 2 other component of the phenotype (data not shown).
region is extended down to the T11H breakpoint
(Fig-Conclusions: ure 3B), (2) the proximal Chr 7 region is reduced to
the centromeric region (Figure 4B), (3) the proximal 1. MatDp for the regions distal to the T138Ca and T45H boundary of the central region on the paternal Chr 7 breakpoints on Chr 9 results in postnatal growth has been brought closer (Figure 4B) to the recognized retardations similar to those found with paternal in- PWS/AS homologous region, and (4) a new imprinting heritance of a knockout and mutation of the pater- region has been identified in Chr 9 distal to the T45H
nally expressed imprinted gene,Rasgrf1. breakpoint (Figure 7).
2. The Rasgrf1 locus lies distal to the translocation Changes in the imprinting effects: The imprinting breakpoints on Chr 9 and on the basis of the knock- effects have also changed such that now (1) both the out and mutation phenotypes is likely to be the gene proximal Chr 2 and proximal Chr 7 MatDp imprinting responsible for the MatDp(dist9) growth retardation. effects comprise fetal and placental growth retardations 3. The similarities between the PatDp(dist2) phenotype with effectively normal postnatal viabilities (Figures 3B and that of the maternally inheritedGnasmutation and 4B), (2) PatDp for proximal Chr 2 has now been (Oed-Sml) imply that theGnaslocus is responsible for found to have an imprinting effect comprising a placen-the edematous component of placen-the former phenotype. tal overgrowth (Figure 3B), and (3) MatDp for distal 4. The absence of a behavioral effect and the enhanced Chr 9 has now been seen to have an imprinting effect postnatal viability of T11H PatDp(dist2)/MatDp comprising a postnatal growth retardation (Figure 6).
Chr 7 study clearly separates the two imprinting regions, a placental growth retardation, with perhaps a second-ary fetal growth retardation (Table 2), while PatDp both observed with MatDp, for the first time (Figure
4B). Their interaction to provide a compound third (prox11) brings about the fetal overgrowth followed by a like effect in the placenta (Cattanachet al. 1996). imprinting effect could most simply be explained by
the additive effects of the proximal prenatal growth Together, the MatDp(prox2)/PatDp(prox11) genotype might therefore have been expected to give a large fetus retardation and the central postnatal lethality, giving a
combined fetal/placental growth retardation with neo- and a small placenta. Instead, an as-yet-undetected early embryonic lethality was indicated (Table 3). This might natal lethality. However, the other interactions, which
combine MatDp for one chromosome with PatDp for be interpreted in terms of a failure of the small, possibly less functional placenta to sustain a large fetus, although another, potentially provide new insight into the
regula-tion of fetal and placental growth and also the growth this was not seen with a Chr 7-Chr 11 interaction (Figure 4B, to be discussed). Whatever the exact mechanism, it axes upon which imprinting operates.
Implications of the interactions:Each of the individ- might appear that the Chr 2 and Chr 11 effects operate independently and primarily in different sites (fetus and ual imprinting effects, seen separately with MatDp and
PatDp for the various regions, generally accord well with placenta, respectively). Therefore, while one cannot rule out a common growth axis, this cannot be the basis theMooreandHaig’s (1991) “conflict hypothesis” by
which maternally expressed imprinted genes favor the for the interaction observed.
The candidate Chr 2 and Chr 11 genes can be accom-well-being of the mother over the growth of the fetus
and paternally expressed genes favor fetal growth. Thus, modated within the same framework. Gatmlies within the revised Chr 2 imprinting region (Figure 3B) and is MatDp genotypes with two copies of a maternally
ex-pressed gene might present extra disadvantage to fetal expressed only in the placenta and only from the mater-nal allele (Sandellet al.2003). It encodes a metabolic growth, whereas PatDp genotypes might potentially
en-hance fetal growth. Each of the interactions described enzyme involved in creatine synthesis and has been pos-tulated to be a growth suppressor.Grb10is the candi-combine MatDp for one region with PatDp for another.
Therefore, if the genes involved in any imprinting effect date gene of choice for the Chr 11 effect as maternal transmission of a knockout enhances fetal and placental combination operate upon the same growth axis, then
some balance, such as seen with the combination ofIgf2 growth in the same way, as seen with PatDp(prox11) (Miyoshiet al.1998;Charalambouset al.2003). Con-andIgf2rknockouts to rescue the nullIgf2rphenotype
(Filson et al. 1993), might be expected. However, if sidered to be another maternally expressed growth re-pressor,Grb10encodes a signaling protein that interacts there is a combination of MatDp and PatDp imprinting
genes working in different axes, either an additive effect with tyrosine kinase receptors that include growth factor receptors. With PatDp(prox2)/MatDp(prox11), the such as that found with the combination ofGrb10and
Igf2 knockouts (Charalambous et al. 2003) or some large placenta could be attributed to the absence of the maternally expressedGatmsuppressor, while the small exaggeration of the “conflict” might be generated.
Inter-pretation of most of the interactions described here, fetus could be attributed to the presence of two mater-nal copies of the Grb10 suppressor. In the reciprocal however, is complicated by a number of uncertainties.
These include correct identification of the genes in- MatDp(prox2)/PatDp(prox11) genotype, two maternal copies of theGatmsuppressor operating in the placenta volved and whether it is the presence of two expressed
copies or the absence of an expressed copy of an im- with the absence ofGbr10expression to suppress growth in the fetus might explain the lethality. There is no evi-printed gene that causes the observed imprinting
ef-fects. Further difficulties concern the sites of effective dence that the two genes may operate in the same growth axis.
gene action (for example, fetusvs.placenta), the
rela-tionship between fetal and placental growth, the rela- Chr 7–Chr 11 interaction: With the Chr 7–Chr 11 combination, evidence of yet another fetal and placental tionship between placental size and function, and, in
one case, even transient effects. growth interaction is suggested (Table 3). Separately,
both MatDp(prox7) and PatDp(prox11) affect fetal and
Chr 2–Chr 11 interactions: Such complications are
found with the proximal Chr 2–proximal Chr 11 interac- placental growth, the former reducing growth and the latter increasing it. Postnatally, both genotypic classes tion (Table 3). Separately, PatDp(prox2) does not affect
fetal growth but clearly operates in the placenta to cause retain their growth differences through to adulthood. When together, the Chr 11 effect initially (14.5 days) a placental overgrowth (Table 2), while MatDp(prox11)
causes a fetal and placental growth retardation that ap- overrode the Chr 2 effect in the fetus, and the reverse occurred in the placenta to give a large fetus with small pears to be initiated first in the fetus (Cattanachet al.
1996) and perhaps therefore primarily operates here. placenta (Figure 5A). At later prenatal ages, a different interaction was observed: the fetal growth rate appeared Together, in the MatDp(prox11)/PatDp(prox2)
combi-nation, the fetus showed the Chr 11 growth retardation to decline while the placental growth rate increased, and newborn mice of this genotype were normal sized. and the placenta showed the Chr 2 overgrowth (Table 3).
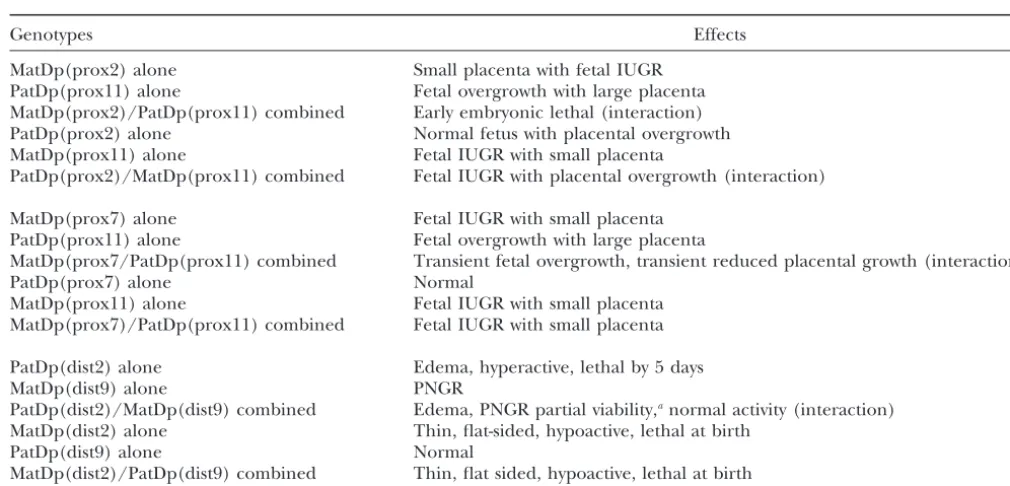
TABLE 3
Summary of imprinting effect interactions
Genotypes Effects
MatDp(prox2) alone Small placenta with fetal IUGR PatDp(prox11) alone Fetal overgrowth with large placenta MatDp(prox2)/PatDp(prox11) combined Early embryonic lethal (interaction) PatDp(prox2) alone Normal fetus with placental overgrowth MatDp(prox11) alone Fetal IUGR with small placenta
PatDp(prox2)/MatDp(prox11) combined Fetal IUGR with placental overgrowth (interaction)
MatDp(prox7) alone Fetal IUGR with small placenta PatDp(prox11) alone Fetal overgrowth with large placenta
MatDp(prox7/PatDp(prox11) combined Transient fetal overgrowth, transient reduced placental growth (interaction)
PatDp(prox7) alone Normal
MatDp(prox11) alone Fetal IUGR with small placenta MatDp(prox7)/PatDp(prox11) combined Fetal IUGR with small placenta
PatDp(dist2) alone Edema, hyperactive, lethal by 5 days
MatDp(dist9) alone PNGR
PatDp(dist2)/MatDp(dist9) combined Edema, PNGR partial viability,anormal activity (interaction)
MatDp(dist2) alone Thin, flat-sided, hypoactive, lethal at birth
PatDp(dist9) alone Normal
MatDp(dist2)/PatDp(dist9) combined Thin, flat sided, hypoactive, lethal at birth
IUGR, intrauterine growth retardation; PNGR, postnatal growth retardation.
aBecause of the various phenotypic effects and reduced viability, it is difficult to deduce whether or not postnatal growth is
impaired.
thereafter (Figure 5B). This may present a different lowing within a few days when generated with all translo-cations other than T11H (Figure 2B). MatDp(dist9) situation from the Chr 2–Chr 11 interaction in which the
limited prenatal and more extensive historical postnatal alone causes a postnatal growth retardation starting soon after birth (Figure 7). When combined in PatDp data (Cattanach and Kirk 1985; B. M.Cattanach,
unpublished results) showed no evidence of a temporal (dist2)/MatDp(dist9) animals, achieved with T11H, the key observation is that the hyperactivity of the edema-effect. The overall picture for the Chr 7–Chr 11
interac-tion therefore suggests that the two imprinting effects tous PatDp(dist2) class is lost and the viability substan-tially improved (Table 3). The connection between the operate in both the fetus and the placenta but perhaps
at different but overlapping stages of development, and two effects is not immediately clear but consideration of the genes involved provides one basis for hypothesis. this does not immediately suggest a common growth
axis. Without any detectable PatDp(prox7) imprinting For Chr 2, the complexGnaslocus has a clear relation-ship with the PatDp(dist2) phenotype because (1) it lies effect, the PatDp(prox7)/MatDp(prox11) combination
produced only the MatDp(prox11) fetal and placental within the 7-Mb region of Chr 2 involved (Williamson et al.1996), and (2) theOed-Smlmutation, which gives an growth retardation.
The Chr 7 and Chr 11 candidate genes can again be almost identical edematous phenotype when maternally inherited, has been shown to result from a base-pair accommodated within the interpretation. The best
can-didate for proximal Chr 7 is Peg3, a gene of unknown substitution within exon 6 ofGnasitself (Skinneret al. 2002). For Chr 9, Rasgrf1is surely responsible for the function although expressed in a variety of embryonic
tissues and placenta (Kuroiwaet al.1996;Relaixet al. postnatal growth retardation because (1) the gene lies within in the appropriate region of the chromosome, 1998). APeg3mutation has been shown to cause a
simi-lar fetal and placental growth retardation (Liet al.1999) (2) both a knockout (Itieret al.1998) and a null muta-tion (Clapcott et al. 2003) of the gene, on paternal to that seen with MatDp(prox7). There is no reported
interaction with the Chr 11 candidate gene,Grb10. transmission, cause postnatal growth retardations simi-lar to that of PatDp(dist2)/MatDp(dist9) mice (Figure
Chr 2–Chr 9 interactions:Finally, with the Chr 2–Chr
9 combination achieved using the T11H translocation 7), (3) Rasgrf1expression is absent in brain of PatDp (dist2)/MatDp(dist9), and (4) the extreme behavior some basis for the observed interaction is indicated.
PatDp(dist2) alone causes the anomalous phenotype at and inviability of PatDp(dist2) animals generated with another translocation (T26H) that does not involve Chr birth of short square body, edema, “paddle” feet, kinked
The basis for the interaction cannot be precisely speci- is an interaction betweenGnasandRasgrf1, it is still unclear fied but a likely possibility is that the PatDp(dist2) behav- if this operates through a common growth axis.
ioral and inviability effects stem from a neuronal abnor- Conclusions: The findings illustrate considerable mality in the brain. Consistent with this hypothesis is complexity in the interactions of imprinted genes in the fact that the maternally expressed Gnas and the development. It seems likely that some of these genes paternally expressedGnasxleach produce, in addition operate largely or exclusively in the placenta (proximal to their respective full-length proteins, shortened forms Chr 2), while others may operate primarily in the fetus of their proteins in neural tissue (Crawfordet al.1993; (subproximal Chr 6;Beecheyet al.2004), and in other Pasolliet al.2000;PasolliandHuttner2001). The cases (proximal Chrs 7 and 11) they may operate in absence of maternal neuronalGnasor the presence of both. Temporal factors may also be involved. Placental two paternal copies of neuronalGnasxlmight therefore and fetal growths do not appear to be interdependent, account for the PatDp(dist2) behavioral and inviability but rather are independently regulated, and placental phenotype. Several observations suggest that it is the size in itself may (Chr 2–Chr 11 interaction) or may not absence of theGnasneural protein, however, that might (Chr 7–Chr 11 interaction) be important in sustaining be involved, but the evidence is not strong. Thus: fetal development. The interactions described raise a multitude of questions concerning development and 1. Partially trisomic mice with one maternal copy in
the role of imprinting that may be investigated by the addition to two paternal copies of Chr 2 have not
use of combinations of candidate gene knockouts or been found to show the behavioral effect (Beechey
combinations of uniparental duplications and such andPeters1994). This observation should exclude
knockouts. a role for the Gnasxl neural protein, but few such
Much of the data presented are consistent with the animals have been produced and other
abnormali-conflict hypothesis (Moore and Haig 1991). Growth ties attributable to the chromosomal unbalance may
enhancements, whether in the fetus or placenta, are have disguised the PatDp(dist2) behavioral effect.
seen with PatDps, and growth suppression is evident 2. Mice with a maternally inheritedGnasknockout (Yu
with MatDps. This applies irrespectively of whether the et al.1998), which should lack both full-lengthGnas
candidate genes are growth repressors, for MatDps, or protein andGnasneural protein, exhibit a behavioral
promoters, for PatDps. It may therefore be that the effect. This might implicateGnas, but it is not clear
imprinted genes do serve a maternal-fetal conflict. The if the behavioral abnormality equates to that seen
anomaly that the main growth-promoting gene,Igf1, is with PatDp(dist2).
not imprinted is also possibly resolved by the Chr 2–Chr 3. When maternally derived, theOed-Sml mutation
re-sults in a nonfunctional full-length Gnas protein 9 findings as these suggest that Igf1may be regulated (Williamsonet al.2004) but, because the mutation by imprinted genes to behave as if it were imprinted.
lies downstream of the genome sequence for the However,HurstandMcVean(1998) have reviewed a
neural protein, the latter should not be affected. number of observations that do not fit with the conflict
Indeed, such mice do not show the PatDp(dist2) hypothesis of which the preweaning growth-retarded
behavioral effect. PatDp(cen7) (Cattanach et al. 1992) and maternal
Ube3a knockout (Jiang et al. 1998) “Angelman” mice On balance it would seem most likely that the absence
provide significant examples. Nevertheless, imprinting of theGnasneural protein is responsible for the
abnor-clearly plays a vital role in the regulation of early mam-mal behavior in PatDp(dist2) mice, just as the absence
malian development, and its observed diversity in terms of a functional maternal copy of full-lengthGnasprotein
of numerous growth axes, tissues implicated, and timing accounts for the edema component of the phenotype
of action suggests a continuity of the evolutionary mater-(Williamsonet al.2004).
nal-fetal conflict. The original role of imprinting may In the case of Rasgrf1,there are two potential links
have been to aid placentation (Hall 1990), initially with Gnas/Gnasxl neuronal proteins. First, brain is a
promoting the growth of the trophoblast through the tissue in whichRasgrf1is imprinted (Plasset al.1996).
actions of paternal genes (Bartonet al.1984;Surani Second, mice with paternally inheritedOed-Sml or null
et al. 1984) with maternal genes operating to control Rasgrf1 mutations both show diminished IGF1 levels
the growth of this foreign tissue. However, to account (Itieret al.1998; J.Peters, unpublished results), and
for the observed multiplicity in the control of fetal and the growth factor has an important role in the
develop-early postnatal growth by imprinting, it may be con-ment and repair of the central nervous system,
stimulat-cluded that subsequently, throughout evolution, any ing neurone proliferation and myelination (Zumkeller
mechanism that might favor fetal growth has been ex-1997;Mu¨ lleret al.1999). It is of further note that the
ploited by paternal genes, again counterbalanced by Rasgrf1interaction with PatDp(dist2) enhances prenatal
growth control by maternal genes. This concept is not as well as postnatal viability. This suggests that IGF1 is
inconsistent with the conclusion of Reik et al.(2003) again involved and has a role in prenatal as well as
Rescue of theT-associated maternal effect in mice carrying null
fetal demand for, and placental supply of, maternal
mutations inIgf-2and reciprocally imprinted genes. Development
nutrients. 118:731–736.
Hall, J., 1990 Genomic imprinting: review and relevance to human We thank David Papworth for statistical analyses of the data and
diseases. Am. J. Hum. Genet.46:857–873. Anne-Marie Woodword, Lynn Jones, Jackie McNaughton, Mark
Har-Hurst, L. D., andG. T. McVean, 1998 Do we understand the evolu-rison, and Debra Brooker for their dedicated and conscientious work tion of genomic imprinting? Curr. Opin. Genet. Dev.8:701–708. with the detailed mouse breeding. Itier, J.-M., G. L. Tremp, J.-F. Le´onard, M.-C. Multon, G. Retet
al., 1998 Imprinted gene in postnatal growth role. Nature393:
125–126.
Jiang, Y. H., D. Armstrong, U. Albrecht, C. M. Atkins, J. L. Noebelset al., 1998 Mutation of the Angelman ubiquitin ligase
LITERATURE CITED in mice causes increased cytoplasmic p53 and deficits of
contex-tual learning and long-term potentiation. Neuron21:647–649.
Barton, S. C., M. A. H. SuraniandM. L. Norris, 1984 Role of
Kim, J., X. Lu andL. Stubs, 1999 Zim1, a maternally expressed paternal and maternal genomes in mouse development. Nature
mouse Kruppel-type zinc-finger gene located in proximal
chro-311:374–376.
mosome 7. Hum. Mol. Genet.8:847–854.
Beechey, C. V., and J. Peters, 1994 Dosage effects of the distal
Kuroiwa, Y., T. Kaneko-Ishino, F. Kagitani, T. Kohda, L. L. Li
chromosome 2 imprinting region. Mouse Genome92:353–354.
et al., 1996 Peg3 imprinted gene on proximal chromosome 7
Beechey, C. V., B. M. Cattanach, A. BlakeandJ. Peters, 2004
encodes for a zinc finger protein. Nat. Genet.12:186–190. Mouse imprinting map. MRC Mammalian Genetics Unit,
Ox-Li, L.-L., E. B. Keverne, S. A. Aparicio, F. Ishino, S. C. Bartonet al., fordshire, UK (http://www.mgu.har.mrc.ac.uk/imprinting/
1999 Regulation of maternal behaviour and offspring growth by imprinting.html).
paternally expressedPeg3.Science284:330–333.
Buckle, V. J., and K. A.Rack, 1993 Fluorescent in situ hybrisation,
Miyoshi, N., Y. Kuroiwa, T. Kohda, H. Shitara, H. Yonekawaet
pp. 59–80 inHuman Genetic Disease Analysis: A Practical Approach,
al., 1998 Identification of the Meg1/Grb10 imprinted gene on Ed. 2, edited by K. E.Davies. IRL Press, Oxford.
mouse proximal chromosome 11, a candidate for the
Silver-Rus-Cattanach, B. M., 1986 Parental origin effects in mice. J. Embryol.
sell syndrome gene. Proc. Natl. Acad. Sci. USA3:1102–1107. Exp. Morphol.97(Suppl.): 137–150.
Monk, D., R. Smith, P. Arnaud, M. A. Preece, P. Stanieret al., 2003
Cattanach, B. M., 1991a Chromosome imprinting and its
signifi-Imprinted methylation profiles for proximal mouse chromo-cance for mammalian development, pp. 41–71 inGenome Analysis,
somes 11 and 7 as revealed by methylation-sensitive representa-edited byK. E. DaviesandS. M. Tilgham. Cold Spring Harbor
tion difference analysis. Mamm. Genome14:805–816. Laboratory Press, Cold Spring Harbor, NY.
Moore, T., andD. Haig, 1991 Genomic imprinting in mammalian
Cattanach, B. M., 1991b Lack of genetic background modification development: a parental tug-of-war. Trends Genet.7:45–49. upon distal Chr 2 imprinting. Mouse Genome89:273.
Mouse Genome Database(MGD), 2004 Mouse genome informatics.
Cattanach, B. M., andC. V. Beechey, 1994 Further evidence that The Jackson Laboratory, Bar Harbor, ME (http://www.informatics. differential recoveries of maternal and paternal disomies are not jax.org).
attributable to imprinting. Mouse Genome92:504. Mu¨ ller, E. E., V. LocatelliandD. Cocchi, 1999 Neuroendocrine
Cattanach, B. M., andC. V. Beechey, 1997 Genomic imprinting control of growth hormone secretion. Physiol. Rev.79:511–607. in the mouse: possible final analysis, pp. 118–145 in Genomic Pasolli, H. A., andW. B. Huttner, 2001 Expression of the extra-Imprinting, edited by W.Reikand A.Surani. IRL Press/Oxford large G protein␣-subunit XL␣s in neuroepithelial cells and young University Press, Oxford/London/New York. neurons during development of the rat nervous system. Neurosci.
Cattanach, B. M., andM. Kirk, 1985 Chromosome 11 parental Lett.301:119–122.
source effect upon fetal growth in mice. Genet. Res.45:220–221. Pasolli, H. A., M. Klenke, R. H. Kehlenbach, Y. WangandW. B.
Cattanach, B. M., J. A. Barr, E. P. Evans, M. Burtenshaw, C. V. Huttner, 2000 Characterization of the extra-large G protein Beecheyet al., 1992 A candidate mouse model for Prader-Willi ␣-subunit XL␣s. J. Biol. Chem.275:33622–33632.
syndrome which shows an absence of Snrpn expression. Nat. Pearsall, R. S., K. Imai, H. Shibata, Y. Hayashizaki, V. M. Chapman Genet.2:270–274. et al., 1998 TheRasgrf1-repeat sequence (D9Ncvs53) maps
be-Cattanach, B. M., C. V. Beechey, C. Rasberry, J. JonesandD. tween Mod1 and Rbp1 on mouse chromosome 9 and may define
Papworth, 1996 Time of initiation and site of action of the a putative imprinted region. Mamm. Genome9:261–262. mouse chromosome 11 imprinting effects. Genet. Res.68:35–44. Plass, C., H. Shibata, I. Kalcheva, L. Mullins, N. Kotelevtseva
Cattanach, B. M., J. A. Barr, C. V. Beechey, J. Martin, J. Noebels et al., 1996 Identification of Grf1 on mouse chromosome 9 by
et al., 1997 A candidate model for Angelman syndrome in the RLGS-M. Nat. Genet.14:106–109.
mouse. Mamm. Genome8:472–478. Reik, W., M. Constaˆncia, A. Fowden, N. Anderson, W. Deanet al.,
Cattanach, B. M., H. Shibata, Y. Hayashizaki, K. M. S. Townsend, 2003 Regulation of supply and demand for maternal nutrients S. Ballet al., 1998 Association of a redefined proximal mouse in mammals by imprinted genes. J. Physiol.547:35–44. chromosome 11 imprinting region and U2afbp-rs/U2af1-rs1ex- Relaix, F., X-J. Wei, X. WuandD. A. Sassoon, 1998 Peg3/Pew1is pression. Cytogenet. Cell Genet.80:41–47. an imprinted gene involved in the TNF-NFkB signal transduction
Cattanach, B. M., J.Peters, S.Balland C.Rasberry, 2000 Two pathway. Nat. Genet.18:287–291.
imprinted gene mutations: three phenotypes. Hum. Mol. Genet. Ren, J., S. Lee, S. Pagliardini, M. Ge´rard, C. L. Stewartet al.,
9:2263–2273. 2003 Absence of Ndn, encoding the Prader-Willi
syndrome-Charalambous, M., F. M. Smith, W. R. Bennett, T. E. Crew, F. deleted genenecdin, results in congenital deficiency of central
Mackenzieet al., 2003 Disruption of the imprintedGrb10gene respiratory drive in neonatal mice. J. Neurosci.23:1569–1573. leads to disproportionate overgrowth by an Igf2-independent Sandell, L. L., X.-J. Guan, R. IngramandS. M. Tilghman, 2003 mechanism. Proc. Natl. Acad. Sci. USA100:8292–8297. Gatm, a creatine synthesis enzyme, is imprinted in mouse
pla-Clapcott, S. J., J. Peters, P. C. Orban, R. BrambillaandC. F. centa. Proc. Natl. Acad. Sci. USA100:4622–4627.
Graham, 2003 Two ENU mutations inRasgrf1and early mouse Searle, A. G., andC. V. Beechey, 1990 Genome imprinting phe-growth retardation. Mamm. Genome14:495–505. nomena on mouse chromosome 7. Genet. Res.56:237–244.
Crawford, J. A., K. J. Mutchler, B. E. Sullivan, T. M. Lanigan, Skinner, J. A., B. M. CattanachandJ. Peters, 2002 The imprinted
M. S. Clarket al., 1993 Neural expression of a novel alterna- oedematous-small mutation on mouse chromosome 2 identified tively spliced and polyadenylated GS␣transcript. J. Biol. Chem. new roles for Gnas and Gnasxl in development. Genomics80:
268:9879–9885. 373–375.
de la Puente, A., J. Hall, Y-Z. Wu, L. Gustavo, J. Peterset al., Sunahara, S., K. Nakamura, K. Nakao, Y. Gondo, Y. Nagataet al., 2002 Structural characterization ofRasgrf1and a novel linked 2000 The oocyte-specific methylated region of the U2afbp-rs/ imprinted locus. Gene291:287–297. U2af1-rs1 gene is dispensable for its imprinted methylation.
Bio-chem. Biophys. Res. Commun.268:590–595.
Surani, M. A. H., S. C. BartonandM. L. Norris, 1984 Develop- Yu, S., D. Yu, E. Lee, M. Eckhaus, R. Leeet al., 1998 Variable and ment of reconstituted mouse eggs suggests imprinting of the tissue-specific hormone resistance in heterotrimeric Gsprotein
genome during gametogenesis. Nature308:548–550. ␣-subunit (Gs ␣) knockout mice is due to tissue-specific im-Williamson, C. M., J. Schofield, E. R. Dutton, A. Seymour, C.V. printing of the Gs␣gene. Proc. Natl. Acad. Sci. USA95:8715–
Beechey et al., 1996 Glomerular-specific imprinting of the 8720.
mouse GS␣gene: How does this relate to hormone resistance in Zumkeller, W., 1997 The effect of insulin-like growth factors on Albright hereditary osteodystrophy? Genomics36:280–287. brain myelination and their potential therapeutic application in
Williamson, C. M., S. Ball, W. Nottingham, J. Skinner, A. Plagge myelination disorders. Eur. J. Paediatr. Neurol.4:91–101.
et al., 2004 Acis-acting control region is required exclusively