Effect of tea flavanols on protein processing and quality
control in the endoplasmic reticulum
Doctoral (Ph.D.) Thesis
Éva Judit Magyar
Semmelweis University
Molecular Medicine Doctoral School
Supervisor: Dr. Miklós Csala associate professor, Ph.D. Opponents: Dr. Ferenc Gallyas Jr. associate professor, Ph.D.
Dr. Gábor Lotz assistant professor, Ph.D. Final examination committee:
Chairman: Dr. Péter Gergely professor, D.Sc.
Members: Dr. Katalin Monostory senior research associate, Ph.D. Dr. László Csanády associate professor, Ph.D.
Budapest
2010
Introduction
Protein synthesis and maturation in the endoplasmic reticulum
Several proteins are synthesized on the surface of the endoplasmic reticulum (ER), and are translocated into the lumen or membrane during their synthesis. Some proteins are retained in the ER, others are targeted to their final destination (Golgi, lysosome, plasma membrane, extracellular space) by vesicular transport. The proteins reach their native conformation through co- and posttranslational modifications assisted by chaperons (BiP, GRP94, calnexin, calreticulin) and foldases (PDI, ERp57). One of the typical modifications in the ER is N-glycosylation, i.e. the incorporation of an oligosaccharide moiety into the proteins.
Quality control
A complex system ensures that proteins are not transported to their destination until they have gained their native conformation in the ER. The primary quality control mechanism can recognize immature polypeptides by their exposed hydrophobic side chains, free cystein thiols, or aggregation tendencies. Bip (GRP78) – and other chaperons – binds and retains these proteins, which gives them help and time to repair the wrong conformation.
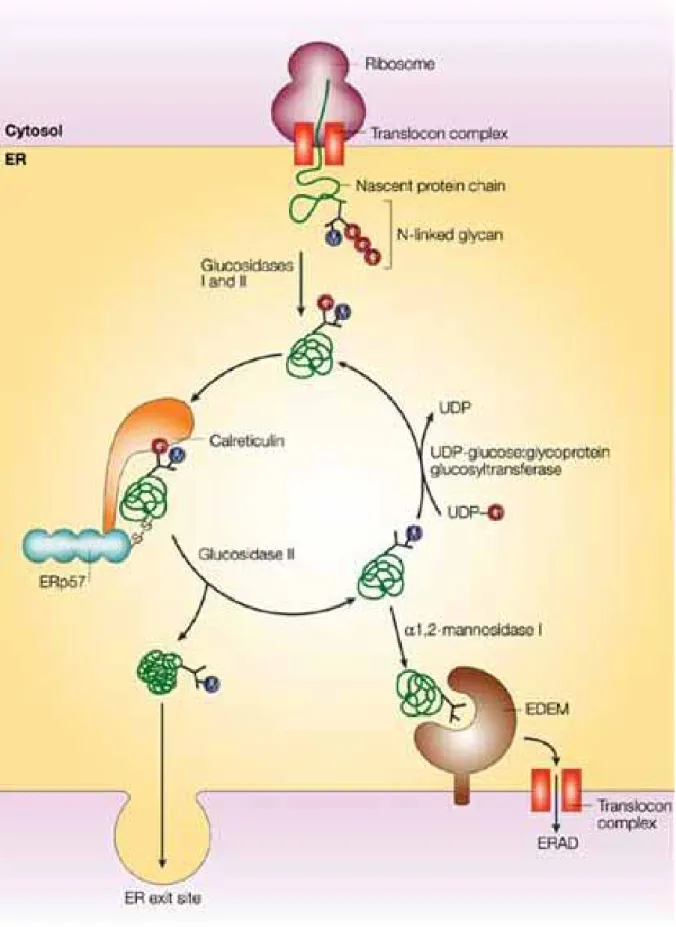
The main components of the best known quality control mechanism are glucosidase II, UDP-glucose:glycoprotein glucosyltransferase (UGGT), calnexin and calreticulin (Figure. 1). UGGT recognizes and glucosylates the misfolded
proteins, so they are labeled as monoglucosylated proteins. These immature proteins are bound by two ER lectins, calnexin and calreticulin, which block the targeting, and help the correction. Glucosidase II cleaves the immaturity signal off, so the protein is released from the lectins. Native proteins can leave the ER towards the secretory pathway, while unfolded proteins are reglucosylated and returned into this cycle (Figure.1). If the protein is unable to fold properly, it is
Endoplasmic reticulum stress
Protein synthesis and maturation in the ER is directly or indirectly affected by conditions interfering with the optimal function of the organelle. Unfolded and/or defective proteins accumulate in the lumen in various cases of ER stress, regardless to the ultimate origin of the disorder. Accumulation of immature proteins activates a specific adaptation mechanism, called „unfolded protein response” (UPR), which attempts to recover the balance between protein synthesis and folding capacity. The four most important elements of the UPR: 1) induction of the ER foldases and chaperones; 2) expansion of the ER by increased synthesis of phospholipids and integral proteins; 3) inhibition of protein synthesis at transcriptional and translational level; 4) stimulation of the ERAD. These responses help immature unfolded proteins obtain their native conformation, or leave the ER and be degraded by the proteasome. If the balance cannot be restored, the UPR can induce apoptosis.
The mechanism of UPR
Proteins specialized to sense ER stress and trigger the UPR („inositol-requiring enzyme 1”: IRE1, „pancreatic ER kinase (PKR)-like ER kinase”: PERK and „activating transcription factor 6”: ATF6) show a number of similarities with hormone receptors of the plasma membrane. The cytosolic domain of these transmembrane proteins transmits the signal, which is detected by the luminal domain (analogous with the extracellular domain). IRE1, PERK and ATF6 are kept in inactive state by BiP chaperone attached to their luminal domains and are activated when BiP dissociates due to luminal accumulation of immature and unfolded proteins in the ER.
IRE1 dimerizes and autophosphorylates and then splices XBP1 mRNA, creating a template for the active transcription factor (sXBP1). ATF6 is translocated to the Golgi, where its cytosolic fragment is liberated by proteolysis and can act as a transcription factor. After dimerization and autophosphorylation,
PERK phosphorylates eukaryotic Initiation Factor 2α (eIF2α), which attenuates general protein synthesis but enhances translation of ATF4 transcription factor.
ATF6, ATF4 and sXBP1 induce among others ER chaperons (BiP, PDI), components of ERAD and phospholipid synthesis, which favors adaptation and survival; but they also induce the proapoptotic protein CHOP, which can contribute to apoptotic cell death.
Apoptosis induced by endoplasmic reticulum stress
Investigations in the last few years revealed that signal transduction pathways activating apoptosis can be initiated by nearly all the orgenelles. ER stress is a severe functional disorder and alarm state in the cells, just like the multiple DNA damage or deficient ATP production. It is not surprising; therefore, that ER stress might cause or promote apoptosis, too.
The above mentioned CHOP transcription factorincreases the synthesis of Bak and Bax proapoptotic proteins (activators of „intrinsic” apoptotic pathway); in addition, it decreases the level of Bcl-2 antiapoptotic protein. Bak and Bax are translocated into the ER membrane, and permit Ca2+ efflux. The elevated
cytoplasmic Ca2+ concentration in turn can disrupt mitochondrial electrochemical gradient and activates calpain. Calpain directly activates procaspase-12 – procaspase-4 in human – and mitochondrial dysfunction triggers the „intrinsic” apoptotic cascade.
Tea as an antitumor herb
The beneficial health effects of green tea got into the focus of scientific interest in the last 20-30 years. Incidences of various cancers and cardiovascular diseases are lower in the Far East countries of Asia, despite high consumption of tobacco. This phenomenon has been postulated as the „Asian Paradox”, and widely attributed to regular consumption of large quantities of green tea.
Tea derives from dried leaves of the evergreen plant (Camellia sinensis).
Tea leaves are remarkably reach in polyphenols (catechins, flavanols), which are the major components of green tea. The most important representative of these
molecules is epigallocatechin-3-gallate (EGCG), which gives 59 % of the total cathechin content, but epigallocatechin (EGC; 19 %), epicatechin-3-gallate (ECG; 13 %) and epicatechin (EC; 6.5 %) are also important.
A number of potential targets have been identified where catechins can prevent tumor cell formation or block tumor progression and metastasis. Besides their impressive antioxidant activity, flavanols inhibit cell migration, proliferation and angiogenesis, and can initiate apoptosis in various types of tumor cells. Among these complex antitumor activities, apoptosis induction is especially important, because it can kill the existing tumor cells and reduce tumor mass. The exact mechanism of the proapoptotic effect of catechins has not been fully clarified, and the role of signal pathways from subcellular organelles cannot be ruled out.
Aims
The antitumor effects of tea catechins are manifested through various mechanisms and target points. The enhanced apoptosis of tumor cells might be due to ER dysfunction and activation of the UPR. We hypothesized that tea flavanols can cause ER stress by affecting the protein folding and quality control mechanism, and that the proapoptotic mechanisms initiated by the ER contribute to the apoptosis of the tumor cells.
The aim of the present study, therefore, was to elucidate whether the disturbance of protein maturation in the ER and activation of the UPR can play an important role in the antitumor effects of tea flavanols. After answering this fundamental question, we attempted to identify the molecular target of tea flavanols in the ER. We wanted to answer the following questions:
- Can we detect ER stress and components of the UPR in EGCG treated hepatoma cells? Are the proapoptotic components of the UPR activated? After we confirmed the apoptosis inducing effect of EGCG in our hepatoma cells, we investigated the expression of ER chaperons and foldases, phosphorylation of eIF2α, morphological alterations in ER as
typical elements of the UPR. In addition, we studied the proapoptotic elements: CHOP-induction, procaspase-12 cleavage, slow depletion of the calcium stores of the ER, which have great importance regarding our hypothesis.
- Can EGCG – and other polyphenols – inhibit glucosidase II enzyme, which plays an important role in protein processing and quality control in the ER?
Glucosidase activity was measured by using artificial substrates in hepatoma cells and rat liver microsomes. After confirming the inhibiton of glucosidase II in hepatoma cells the phenomenon was further characterized in rat liver microsomes. Besides determining the concentration-dependence, the type of inhibition was also analyzed by assessing alterations in the kinetic parameters. The IC50 and Ki values
characterizing the inhibitory effects of the most important tea flavanols and a synthetic gallate derivative were calculated and compared with those of a widely known glucosidase inhibitor.
Materials and methods
Cell culture: Hepa1c1c7 mouse hepatoma cell line was grown in α-MEM
supplemented with 10 % fetal bovine serum, 2mM L-glutamine and 1 % antibiotic/antimycotic at 37° in 95 % air and 5 % CO2 in 10 cm diameter cell
flask. EGCG was dissolved in culture medium and added to the cell culture at 70–80 % confluence at 100-fold dilution.
Apoptosis assay: The cells were grown on sterile Burker counting chamber
coverslip. Fluorescent dyes (2 % annexin V-FITC and 2 % propidium iodide) (Annexin-V-FLUOS Staining Kit, Roche) were added to the cell culture after 70 % confluence. After 15 minutes incubation at room temperature, the coverslip with dyed cells was placed in Burker counting chamber, and annexin labeled apoptotic green cells and propidium iodide labeled necrotic red cells were
counted using fluorescence microscope. The apoptosis index was determined as apoptotic cells / bodies per 100 cells.
Cell lysate: Cells were washed once with PBS, then 200 µl lysis buffer (250
mM NaCl, 50 mM HEPES-Na pH 7.4, 1 mM EDTA-Na, 1 mM EGTA-Na, 1.5 mM MgCl2, 0.1 % Nonidet-P-40, 1 % protease-inhibitor coctail, 1 mM PMSF,
10 mM benzamidin, 20 mM NaF, 1 mM para-nitrofenil-phosphate) was spread on the culture. Cells were kept on ice during the whole process of harvesting. After incubation for 1-2 min, the cells were scraped off the plates, suspended, sonicated and centrifuged (10 minutes, 10000 x g, 4 °C). The supernatants were stored at -20°C until used. Protein concentration of cell lysates was measured by using BioRad “micro protein assay kit”.
Western blot assays: The samples were incubated in a sample buffer including 2
% β-mercaptoethanol for 10 minutes at 95°C. Equal amounts of proteins (20 μg) were separated in 9% SDS-PAGE and transferred to NC or PVDF membrane (60 minutes, 100 V). Equal protein loading was verified by red Ponceau-staining of the membrane. Monoclonal (for procaspase-12 and CHOP) and polyclonal (for other investigated proteins) primary antibodies were used and detected with appropriate horseradish peroxidase conjugated secondary antibodies.
Fluorescence microscopy with endoplasmic reticulum labeling: The
morphology of the ER in the hepatoma cell cultures was studied using a Nikon Eclipse TE300 microscope equipped with Nikon S Fluor40X or Planapo100X objectives and a FITC filter set and Photometrics Cool SNAP HQ CCD camera. Equipments were driven by MetaMorph, Meta Imaging SeriesTM software. The fluorescent images were acquired after labeling the ER with 0.1 μM BODIPY® FL thapsigargin.
Measurement of cytosolic free calcium levels: Alterations in the cytosolic free
calcium concentration were detected with microscopy using a fluorescent probe. Control and EGCG-treated hepatoma cells were loaded with 3 µM Fluo-3 acetoxymethyl ester for 30 minutes at 37 °C as detailed earlier. After loading,
cells were kept at 25 °C in Krebs buffer (140 mM NaCl, 5.4 mM KCl, 1 mM MgCl2, 1 mM CaCl2, 15 mM Hepes pH 7.4). In order to prevent leakage of
intracellular Fluo-3, 100 µM sulfinpyrazone was included in the medium. The results were not affected by sulfinpyrazone. Thapsigargin (1 μM) or EGCG (10-100 μM) was added to the cells and fluorescence microscopy was carried out using the equipment described above driven by MetaFluor, Meta Imaging SeriesTM software. At the end of each incubation, ionomycin (10 μM) plus CaCl2
(10 mM) followed by addition of EGTA (20 mM) were added to determine the maximal and minimal fluorescence values, respectively.
Measurement of glucosidase II activity in hepatoma cells: α-Glucosidase
activity of Hepa 1c1c7 cells was determined as the rate of 4-methylumbelliferone production by enzymatic hydrolysis of methylumbelliferyl-glucopyranoside (MUG). Confluent cells were trypsinized, counted, diluted to about 107 cells/ml and divided into two aliquots. The cellular membranes were permeabilized in one aliquot by incubation with the pore-forming antibiotic alamethicin (0.2 mg/ml) at 37 ºC for 5 min. The cells were then centrifuged (250 x g for 4 min) washed with PBS. Finally, the cells were resuspended in serum-free α-MEM at 5 x 106 cells/ml and pre-incubated at 37 ºC in the presence or absence of 100 μM EGCG for 0-12 hrs. The reaction was started by the addition MUG at 50 μM final concentration. Samples were taken after 0, 15, 30, and 45 minutes, mixed with two volumes of ice-cold methanol and stored at -20 ºC until analysis. They were centrifuged (10 min, 4 ºC, at 20 000 x g) and the 4-methylumbelliferone content of protein-free supernatants was measured by HPLC.
Preparation of rat liver microsomal vesicles: Microsomes were prepared from
livers of overnight fasted male Wistar rats (180-230 g), using fractional centrifugation. The ER vesicles were washed and re-suspended in MOPS-KCl buffer (100 mM KCl, 20 mM NaCl, 3 mM MgCl2, 20 mM MOPS; pH 7.2) then
immediately frozen in liquid nitrogen and kept in liquid nitrogen until use (within 6 months). The protein concentration in microsomal samples was
determined using the method of Lowry. Purity of the microsomes was assessed by a marker-enzyme analysis (mitochondria: cytochrome c oxidase, ER: glucose 6-phosphatase, cytosol: 5’-nucleotidase). The integrity of the microsomal membranes was assessed using the mannose-6-phosphatase assay,which showed a latency greater than 95 %.
Measurement of glucosidase II activity in microsomes: Two specific artificial
substrates of glucosidase II were used: MUG and 4-nitrophenyl-glucopyranoside (NPG). Methylumbelliferone was detected by Cary-Varian Spectrofluorimeter, 4-nitrophenol was quantified by using HPLC. Microsomal membranes were permeabilized with the pore-forming antibiotic (0.1 mg/mg microsomal protein). Inhibitors were added to the microsomal suspension 1-2 minutes prior to the substrate (MUG or NPG).
Statistical analysis: The enzyme activities are presented as pmol/min/million
cells and nmol/min/mg. Derived parameters (Km, Vmax, IC50 and Ki) were
calculated, statistical analysis (ANOVA) and diagrams were made by GraphPad Prism 4.01 software.
Results
EGCG activated ER-derived proapoptotic mechanism
Activation of apoptosis:The number of annexin-positive (apoptotic) hepatoma
cells did not change in the first 6 hrs of EGCG treatment. However, the frequency of apoptosis increased remarkably 12 hrs after the addition of EGCG and further elevated in the next 12 hrs. The apoptotic index of untreated cells did not change during the one day long parallel incubations.
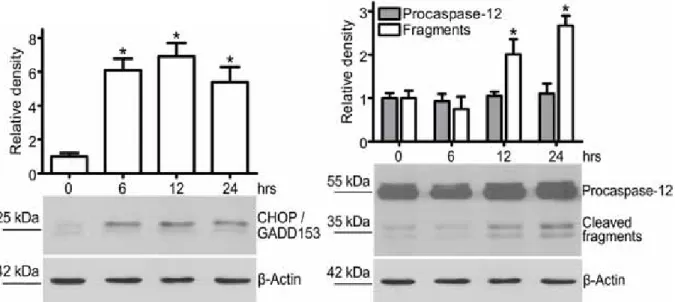
CHOP induction and procaspase-12 cleavage: In accordance with the
stimulation of apoptosis, activation of ER-related death-promoting mechanisms was observed in the EGCG-treated cells. Western blot analysis revealed the induction of CHOP protein and the proteolytic activation of procaspase-12
(Figure 2) with different time-courses. The amount of CHOP reached its
maximum at 6-7-times above the control level within 6 hrs and it was sustained for at least 24 hrs after the addition of EGCG. While the expression of procaspase-12 protein increased slightly (but not significantly), the amount of cleaved caspase-12 fragments changed parallelly with the apoptotic index, as its elevation was evident after 12 hrs and became even more pronounced after 24 hrs.
Figure 2 CHOP induction and procaspase-12 cleavage
Chaperone and foldase expressions: Since the previous findings indicated the
activation of ER stress response in the EGCG-treated cells, further ER stress markers were investigated under the same conditions. The amount of five investigated proteins (GRP94, BiP, calnexin, PDI and ERp72) was monitored using Western blot analysis; and a steady expression of all the five proteins was revealed without any obvious alteration during the 24 hrs of incubation.
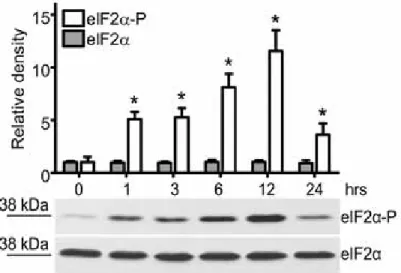
Increased phosphorylation of eIF2α: The amount of phosphorylated eIF2α
subunit increased after 1 hr and further elevated until the 12th hour of EGCG-treatment then returned to the initial level (Figure 3). The effect was caused by
enhanced protein phosphorylation rather than protein induction because the immunoblot band intensities were unaltered when a phosphorylation-unspecific anti-eIF2α antibody was used.
Figure 3 Increased phosphorylation of eIF2 α
Depletion of ER calcium stores: The cytosolic free Ca2+ concentration in the
hepatoma cells was assessed by fluorescent microscopy in the absence of extracellular calcium to avoid influx. Addition of EGCG (100 µM) did not cause an instant calcium release from the internal stores and did not increase the cytosolic free calcium level within 5 min. The effect of longer EGCG-treatments on the ER calcium stores was studied by measuring the thapsigargin-releasable Ca2+ in the same conditions.
Treatment of the cells with increasing doses of EGCG (10 to 100 µM) for 18 hrs caused a progressive reduction of ER calcium content (Figure 4 left). The
luminal calcium stores were almost completely depleted at 50 µM and virtually no thapsigargin-releasable Ca2+ remained at 100 µM EGCG concentration. The time-dependence of the observed effect was investigated at 100 µM EGCG. The initially slow Ca2+ store depletion was accelerated after 2 hrs and was practically complete within 4 hrs after the administration of EGCG (Figure 4B right).
The alterations in ER morphology: EGCG-treatment induced evident alterations
in the ER network morphology as detected by fluorescence microscopy. Comparison of the cells before and after EGCG-treatment clearly revealed derangement of the reticular structure. The most obvious alterations were the formation of ER-derived vacuoles and retraction of the ER network from the plasma membrane.
Effect of various tea flavanols on glucosidase II
Our results suggest that EGCG causes the derangement of luminal protein processing and the immature, unfolded proteins accumulate in the ER lumen of the hepatoma cells. Inhibitors of glucosidase II are known to be polyhydroxylated compounds like the flavanols, and hence we assumed that the observed ER stress may be due to inhibition of glucosidase II.
Inhibition of glucosidase II activity in mouse hepatoma cells: The effect of
EGCG on glucosidase II was first investigated in intact and permeabilized Hepa 1c1c7 cells by measuring the rate of MUG hydrolysis in the presence and absence of the flavanol. Glucosidase activity was significantly inhibited by EGCG. The extent of inhibition was 81 % and 83 % in intact and permeabilized cells, respectively.
Target and concentration-dependence of inhibition: The effect of EGCG was
studied in rat liver microsomes by using MUG substrate (40 μM). Concentration dependent inhibition (IC50 = 50.9 μM; Ki = 29.5 μM) was observed in the
The latency of glucosidase activity decreased with the increasing EGCG concentration; the latency was practically abolished above 150 µM EGCG. Our results indicate direct inhibition of glucosidase II by the flavanol.
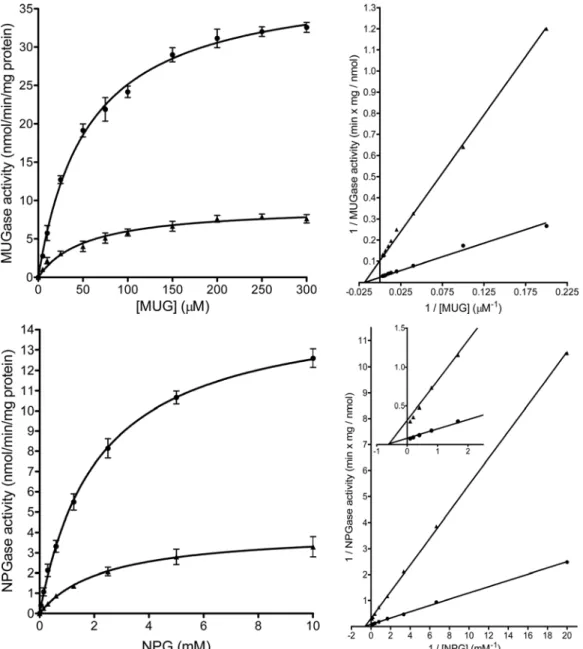
Kinetic characteristics of the inhibition: Permeabilized microsomes were treated
with 100 μM EGCG, and the enzyme activity was measured using MUG (5-300 μM), and NPG (0.05-10 mM) (Figure 5). EGCG treatment resulted in a
four-fold decrease in vmax, while Km values did not change significantly.
Figure 5 Kinetic analysis of the inhibitory effect on glucosidase II
Comparison of the inhibitory effect of tea flavanols: It was also investigated
whether glucosidase II can be inhibited by other tea cathecins like epicatechin gallate (ECG), gallocatechin (GC), epigallocatechin (EGC), and gallocatechin
gallate (GCG). Inhibitory effect of these agents was compared with that of N -butyldeoxynojirimycin (NBDJ) on the basis of the results obtained using MUG and NPG substrates. All investigated flavanols inhibited the enzyme, although with different efficiencies (Table 1). The effect of EGCG can be considered as
intermediate compared with the other agents. The enzyme activity was almost totally blocked at higher concentrations of GCG and ECG, similarly to NBDJ. Even the least effective tea flavanols (GC and EGC) caused a nearly 30% decrease in the activity at 200 μM concentration.
MUGase NPGase inhibitor IC50 (μM) Ki (μM) IC50 (μM) Ki (μM) GCG 3.502 2.027 3.702 2.545 ECG 15.14 8.763 19.06 13.10 EGCG 50.92 29.48 47.72 32.81 EGC 117.7 68.15 110.5 75.99 GC 136.3 78.93 102.7 70.62 NBDJ 8.965 5.190 3.268 2.247
Table 1: IC50 és Ki values of various inhibitors of glucosidase II
In summary, EGCG treatment induces ER stress activates the UPR in the hepatoma cells studied. The exact mechanisms in the background need to be elucidated. Earlier studies suggested a direct inhibition of GRP78/BiP chaperon, but the disturbance of ER functions can evolve through other molecular targets. Our study revealed that EGCG and other tea flavanols directly inhibit glucosidase II, a key enzyme of protein processing and quality control in the ER. Activation of the apoptosis inducing elements of the UPR – irrespectively to the origin of ER stress – can contribute to the antitumor effects of catechins observed in vivo and in vitro. Our results raise the possibility of developing novel antitumor drugs inducing tumor cell apoptosis through interfering with protein maturation and quality control in the ER.
Conclusions
We studied the apoptosis inducing effect of EGCG, which is one of the most important elements of the antitumor effects of tea flavanols. We aimed to answer the question whether EGCG can initiate proapoptotic signals from ER, and also wanted to identify the molecular target, through which the stress response is induced in the organelle. We can draw the following conclusions:
1. EGCG treatment induces apoptosis in Hepa1c1c7 hepatoma cells. Beside the activation of programmed cell death, ER-derived proapoptotic signals (CHOP-induction, procaspase-12 cleavage, slow depletion of ER calcium stores) were also detected, so the induced apoptosis was at least partly of ER origin.
2. EGCG treatment induces ER stress in the hepatoma cells, which was well indicated by the morphological changes of the organelle. The most important proapoptotic elements of the UPR as well as the enhanced phosphorylation of eIF2α were detected, although the activated partial UPR did not involve induction of the major ER chaperones and foldases.
3. The ER stress induced by EGCG might be due to the inhibition of glucosidase II enzyme by this and other tea flavanols, since the enzyme plays a key role in glycoprotein processing and quality control.
4. Inhibition of glucosidase II was be demonstrated in hepatoma cells and rat liver microsomes. The presence of gallate group in the compounds seems to be necessary for an efficient inhibition of glucosidase II, and the configuration of the gallo moiety also influences the efficiency.
IC50 and Ki parameters of the most efficient teaflavanol (GCG) were
Our results collectively lead us to the conclusion that EGCG interferes with the protein maturation and quality control in the ER lumen by inhibiting glucosidase II enzyme. This leads to a stress in the organelle, and the consequently triggered partial UPR contributes to the apoptotic cell death. On the one hand, it may play a significant role in the well-known antitumor effect of polyphenols. On the other hand, it raises a novel approach in antitumor therapy, i.e. it provides endoplasmic reticulum and the components of the local protein processing machinery as potential drug-targets.
List of publications
Publications related to the Ph.D. thesis
1. Gamberucci A., Konta L., Colucci A., Giunti R., Magyar J.É., Mandl J.,
Bánhegyi G., Benedetti A., Csala M.: Green tea flavonols inhibit glucosidase II. Biochem. Pharmacol. 72, 640-646, 2006; IF: 3.581
2. Magyar J.É., Gamberucci A., Konta L., Margittai É., Mandl J., Bánhegyi
G., Benedetti A., Csala M.: Endoplasmic reticulum stress underlying the pro-apoptotic effect of epigallocatechin gallate in mouse hepatoma cells.
Int. J. Biochem. Cell Biol. 41, 694-700, 2009; IF: 4.887
3. Magyar J.É., Konta L., Bánhegyi G., Mandl J., Csala M.:
Zöldtea-flavonolok hatása a glukozidáz II enzim aktivitására. (absztrakt) Biokémia
3, 74, 2006 Other publications
1. Révész K., Tüttő A., Margittai É., Bánhegyi G., Magyar J.É., Mandl J.,
Csala M.: Glucuronide transport across the endoplasmic reticulum membrane is inhibited by epigallocatechin gallate and other green tea polyphenols. Int. J. Biochem. Cell Biol. 39, 922-930, 2007; IF: 4.009
2. Csala M., Marcolongo P., Lizák B., Senesi S., Margittai É, Fulceri R, Magyar J.É., Benedetti A., Bánhegyi G.: Transport and transporters in the endoplasmic reticulum. Biochim. Biophys. Acta (Biomembranes) 1768,