With 11 text-figures Printed in Great Britain
MINIATURE EXCITATORY JUNCTION POTENTIALS IN THE
SOMATIC MUSCLE OF THE EARTHWORM,
PHERETIMA COMMUNISSIMA, IN
SODIUM FREE SOLUTION
Y. ITO, H. KURIYAMA AND N. TASHIRO
Department of Physiology, Faculty of Medicine and Dentistry, Kyushu University, Fukuoka, Japan
{Received 8 October 1968)
INTRODUCTION
Neuromuscular transmission of excitation in invertebrates has been studied exten-sively by many investigators (cf. Bullock & Horridge, 1963). Miniature inhibitory junction potentials (m.i.j.p.s) and miniature excitatory junction potentials (m.e.j.p.s) could be recorded from the longitudinal layer of the somatic muscle in the earthworm. Field stimulation to the peripheral nerves elicited inhibitory junction potentials (i.j.p.s) and excitatory junction potentials (e.j.p.s). Generation of the m.i.j.p. and i.j.p. was blocked by treatment with picrotoxin (icr6 g./ml.) and the equilibrium potentials for them ranged between —52 and —58 mV. The m.e.j.p.s and e.j.p.s were blocked by d-tubocurarine (io~6 g./ml.) and the equilibrium potential for them was o mV. The former was thought to be due to release of y-aminobutyric acid (GABA) from the peripheral inhibitory nerves and to selective increase of chloride permeability. The latter was thought to be due to release of acetylcholine which increases the sodium and potassium permeabilities of the postsynaptic muscle membrane. Generation of the m.e.j.p. was less common than that of the m.i.j.p., in spite of the double innervation to the muscle fibres. However, when the external sodium (Na)0 was replaced with (Tris)0, the membrane was hyperpolarized from — 35 mV. to — 55 mV. and frequently generated the m.e.j.p. (Hidaka et al. 1969b, c). The present experiments were intended to investigate the ionic mechanism involved in generation of the m.e.j.p.s in sodium-free solution, and the results indicated that, in sodium-sodium-free solution, generation of the m.e.j.p in a depolarizing direction is due mainly to increase in calcium permeability and, to a lesser extent, to increase in potassium permeability, brought about by the release of acetylcholine from the nerve terminals.
METHOD
The longitudinal layer of the somatic muscle of the earthworm, Pheretima
cotn-munissima, 5-8 cm. in length, was used. The earthworm was pinned on the plate
io8 Y. ITO, H. KURIYAMA AND N. TASHIRO
The normal solution (Ringer's solution) used for this tissue was of the following composition: Na, 140 mM; K, 2 7 mM; Ca, i-8 mM; Mg, i-o mM; Cl, 148-3 HIM; and pH was adjusted from 7-3 to 7-5. The microelectrode was used for making the electrical recording as well as for stimulating by means of the Wheatstone bridge method. The range of the applied current was between io~10 and 5 x io~9 A. The microelectrodes were filled with 3 M-KC1 for the measurement of the various properties of the mem-brane, and were filled with 2 M potassium citrate for the measurement of the reversal potential for the m.e.j.p. Sodium-free solution was prepared using Tris-(hydroxy-methyl)-aminomethane (C4H11NO3) to maintain isosmoticity. The solution was titrated with a high concentration of HC1, and the pH was adjusted to 7-4. The general procedures of the experiment were the same as those described by Hidaka, Ito & Kuriyama (1969 a) and Hidaka et al. (1969J, c). The experiments were carried out through the year. However, during the winter season (November-March) the effective resistance of the muscle fibres in normal solution was nearly twice that measured during the summer season (April-October).
RESULT
Generation of m.e.j.p.
The membrane potential of muscle membrane was very low. However, the sodium-free solution (sodium was substituted by Tris) hyperpolarized the membrane from — 36 mV. (s.E. = ±0-5, n = 50) to ~54mV. (S.E. = ±o-6, n = 50), and increased
Control Na-free (Tris)
Ca2+ 2 mM
2x10"'A. r-0
L - 5 0 mV.
500 msec.
Fig. 1. Effects of pulses of current delivered to the single muscle fibre by the Wheatstone bridge method in normal and sodium-free (Tris) solution, (a) Control; (6) sodium-free solution.
109
(a) m.e.j.p. sodium-free
in solution
(b) d-tubocurarine
10"* g./ml.
(c) After rinsed by sodium-free solution
r 50 mV.
[image:3.451.86.352.61.289.2]500 msec.
Fig. a. Effects of d-tubocurarine (io~6 g./ml.) on the miniature excitatory junction potentials (m.e.j.p.s). (a) Control: sodium-free solution; (6) d-tubocurarine in sodium-free solution (after 5 min.); (c) after washing out with sodium-free solution.
100
90
80
70
60
i
so
40
30
20
10
-50
Amplitude of
m.e.j.p. 40
30
__
20
10
Tin.
.-Frequency of —1 m.e.j.p.
i
j
-i \
\
\ \
\
vUcn,
0-51 020304050 60 7080 t i f » I ? 1 * » 0-91 -92-93-94-95-96-9 7-98-9
mV.
0 1 2 3 4 5 6 7 8 9
Frequency/5 sec.
[image:3.451.74.356.337.603.2]n o Y. ITO, H. KURIYAMA AND N. TASHIRO
after-hyperpolarization of the membrane was not observed in sodium-free solution due to the hyperpolarization of the membrane.
In sodium-free solution the spontaneously generated miniature depolarizing potentials could be recorded more frequently than those in the normal solution. These small potential changes in the resting membrane were not influenced by treatment with picrotoxin (IO~6-IO~5 g./ml.) and atropine (IO~7-IO~6 g./ml.). However, treat-ment with d-tubocurarine (IO~6-IO~5 g./ml.) completely abolished them. Figure 2
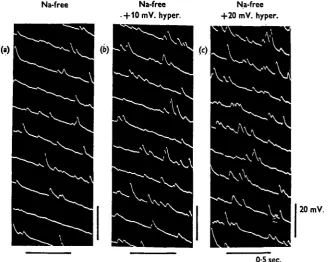
Na-free Na-free +10mV. hyper.
Na-free +20 mV. hyper.
20 mV.
[image:4.451.64.395.164.426.2]0-5 sec.
Fig. 4. Generation of m.e.j.p.s in sodium-free solution, (a) Sodium-free solution; (6) 10 mV. hyperpolarization by inward current in sodium-free solution; (c) 20 mV. hyperpolarization by inward current in sodium-free solution.
shows the effect of d-tubocurarine (io~6 g./ml.) on the the generation of the miniature depolarizing potentials. Therefore, these potential changes are presumably m.e.j.p.s. The spontaneous spike discharges and the spikes elicited by the intracellular depolariz-ing currents could be observed in sodium-free solution because the spike is due to inward movement of calcium ions during the active state of the membrane (Hidaka
et al. 19696). The amplitude and distribution of m.e.j.p.s measured from the single
muscle membrane on the amplitude and frequency of the m.e.j.p.s in sodium-free solution were observed. Since the intensity-voltage relation of the membrane was linear in the ranges of the membrane potential from — 90 mV. to — 20 mV., this indicated that no change of the input resistance of the membrane occurred in the above ranges of the potential changes.
120
100
80
z
6040
20
Na-free solution
Mean 1-9 mV.
1 1 1 1 i I 1-1 1 1 1 1
Amplitude (50 sec.)
Na-free solution + 1 0 mV. hyperpolarization
Mean 2-3 mV.
I I I I
Na-free solution
+20 mV.
h/perpolarization
_n
Mean 2-8 mV.
1 1 1 1 1 1 1 t—«—1—1
0-51 1-9 5 I 5-9 10 i 10-9 0-51 e 1-9 I/I 2 5-9 10 10-9
15 0-51 5
15-9 1-9 5-9 10
10-9 15
15-9
mV. mV. mV.
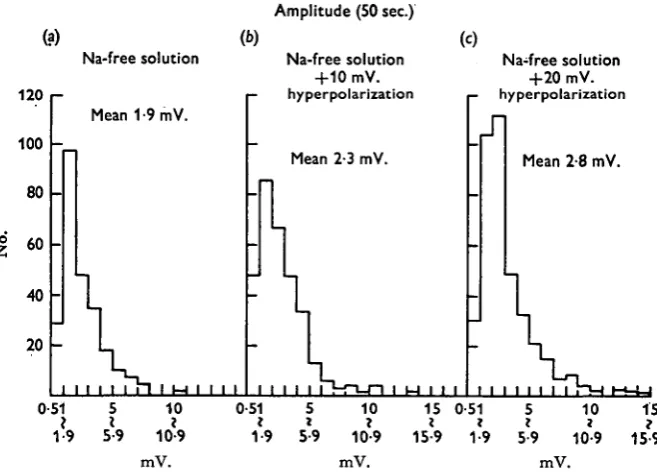
Fig. 5. Histograms of the amplitude of m.e.j.p.s generated within 50 msec, in sodium-free solu-tion, (a) Sodium-free solution; (b) 10 mV. hyperpolarization by inward current in sodium-free solution; (c) 20 mV. hyperpolarization by inward current in sodium-free solution.
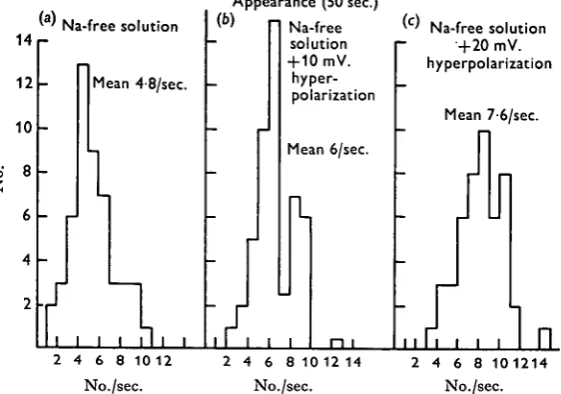
Figure 4 shows the appearance of the m.e.j.p.s in sodium-free solution with 10 mV. and 20 mV. hyperpolarization of the membrane. Figure 5 shows the histograms of the amplitude of the m.e.j.p.s which appeared within 50 sec. in the sodium-free solution and with both 10 mV. and 20 mV. hyperpolarization of the membrane, and Fig. 6 shows the histograms of appearance of the m.e.j.p. within 50 sec. at the different membrane potential levels. T h e hyperpolarizing currents were applied as intracellular polarizing currents in the sodium-free solution. T o prevent the generation of m.i.j.p.s the tissue was previously treated with picrotoxin (io~6 g./ml.). Because the reversal potential level for the m.i.j.p.s was about —55 mV. and conditioning hyperpolariza-tion of the membrane was up to —60 mV., the polarity of the m.i.j.p.s was reversed to the same direction as that of the polarity of the m.e.j.p.s.
In this particular cell the mean m.e.j.p. amplitude was 1*9 mV. and the mean frequency was 4/8/sec. in the sodium-free solution. When the membrane was hyper-polarized from —55 to —65 mV., the mean amplitude and frequency were increased to 2-3 and 6-o mV./sec. respectively, and when the membrane was hyperpolarized to
— 75 mV. these values increased to 2-8 and 7-6 mV./sec. respectively.
[image:5.451.59.388.146.382.2]i i 2 Y. I T O , H . KURIYAMA AND N . TASHIRO
to the increased driving force of the membrane, thus inducing the increased appear-ance of the m.e.j.p.s which were previously lost in the noise level.
When the mean amplitude of the m.e.j.p. was plotted against the membrane poten-tials, the reversal potential level could be estimated from the extrapolated line, and this value was calculated to be - 1 6 mV. from Fig. 5. On the other hand, when the reversal potential level for the m.e.j.p. was measured by the application of the inward current, this value was — 20 mV. (s.E. = + 1-2, n = 11). The difference in the results obtained from these two methods was not significant. However, the main explanation
14 12 10 8 6 4 2
' ' Na-free solution
-_ -- - r -i Mean 4-8/sec.
1
/ A,
Appearance (b) -_--J
2 (50 sec.)
' Na-free
solution + 10 mV.
hyper-polarization
Mean 6/sec.
n i
'c' Na-free solution
+20 mV.
hyperpolarization _
Mean 7-6/sec.
r
- r
1
-I
J
fl
"..rf...
In
2 4 6 8 10 12 No./sec.
2 4 6 8 10 12 14
No./sec.
[image:6.451.87.368.183.380.2]4 6 8 101214 No./sec.
Fig. 6. Histograms of the appearances of m.e.j.p.s within 50 msec, in sodium-free solution, (a) Sodium-free solution; (6) 10 mV. hyperpolarization by inward current in sodium-free solution; (c) 20 mV. hyperpolarization by inward current in sodium-free solution.
for the low value of the reversal potential level measured by the conditioning hyper-polarization of the membrane might be the appearance of the small m.e.j.p.s which were previously lost in the noise level.
Effect of acetylcholine on the input resistance and membrane potential
experiments. Treatment with acetylcholine (io~6 g./ml.) consistently reduced the input resistance of the membrane and pre-treatment with d-tubocurarine (io~6 g./ml.) prevented this. This increased membrane conductance could not be explained by increased potassium conductance alone, because the membrane was slightly depolar-ized by treatment with acetylcholine (io~6 g./ml.) from — 51-2 mV. (S.E. = ±o-6,
n = 25) to —48-2 mV. (S.E. = + 0-7, n = 25), thus indicating that increased
permea-bility to another ion, namely calcium ion, must be involved. However, chloride ion is probably not involved, because (i) picrotoxin (IO~6-IO~5 g./ml.) did not influence the reversal potential level for the m.e.j.p.s, and (ii) in 5-6 mM chloride solution ((Cl)0 was substituted by d-glutamate) the reversal potential level for the m.e.j.p.s remained the same as in normal solution.
o-'A 10
l T ^^~ (a)
1 1 1 1
• ACh 10"' g./ml. ^ •oo ^0
o Control
t # • •*
•
0
/ oo° -•_• _o
A. • O * *
w O
• *° $
00 0 0
o
0 0
°o
O°
o0
10 (b)
• D-TC 10-6 g./ml. 0 Control
••b
10
20
- 3 0
- 4 0
50
Fig. 7. Current-voltage relation of the muscle fibres measured in the presence of acetylcholine (io~fl g./ml.) alone and in the presence of acetylcholine (io~6 g./ml.) together with D-tubo-curarine (io"0 g./ml.). Six different fibres were used, (a) Control (O), acetylcholine (O); (6) control (O), effects of acetylcholine in the pretreatment with d-tubocurarine ( • ) .
Effects of calcium and potassium ions on the reversal potential for m.e.j.p.
Effects of various concentrations of calcium ion on the reversal potential level for the m.e.j.p. were observed in the presence of picrotoxin (icr5 g./ml.).
In the calcium-deficient solution (i, 0-5, 0-2 mM) the muscle membranes were depolarized from — 35-2 mV. (S.E. = ±o-8, n = 25) to — 32-4 mv. (S.E. = ±0-9,
n = 25), to —29 mV. (S.E. = +o-8, n = 25) and to —21 mV. (S.E. = ±o-6, n = 25) respectively, and the membrane resistances were reduced from 68 MQ (S.E. = + 2-4,
n = 6) to 54 MQ (S.E. = ± 2-5, n = 8), to 42 MQ (S.E. = + 2-3, n = 10) and to 37 MQ, (S.E. = ±2-1, n = 8) respectively.
However, in the sodium-free solution calcium concentrations ranging from five times to one fifth the normal concentration change neither the input resistance of the membrane nor the membrane potential. Furthermore, the intensity-voltage relation observed by the intracellular polarizing method under various calcium concentrations
114 Y. ITO, H . KURIYAMA AND N . TASHIRO
in the sodium-free solution showed no rectification by the membrane within ranges of membrane potential from - 9 0 to - 2 0 mV.
Excess calcium concentrations lowered the reversal potential level from — 20 mV. (s.E. = +1-2, n = 11) with 2 mM calcium ion to — 13 mV. (s.E. = + 1-4, n = 10)
>-20
- 4 0
0-4 1 2 2-7 5 10
0-27 0-54 . m M
[image:8.451.124.323.125.327.2]• K-concentration oCa-concentration
Fig. 8. Reversal potential level for the m.e.j.p.s under various calcium (O) and potassium (•) concentrations in sodium-free solution.
+20 T
0
-500 msec.
- 5 0
- 7 0
Fig. 9. Effects of intracellular polarization on the amplitude and polarity of the m.e.j.p.s in sodium-free solution. Continuous line indicates extracellular potential level. KC1, 271HM; CaCl2, 2 mM. Note: m.e.j.p. elicited the spike with overshoot.
with 5 mM calcium ion. When the external calcium was reduced to one fifth of the normal concentration the reversal potential it was increased up to — 36 mV. (s.E. =
[image:8.451.56.398.352.540.2]various external concentrations of calcium and potassium ions. Horizontal bars show twice the s.D. When the external potassium was reduced in the presence of picrotoxin (io~6 g./ml.), the reversal potential level shifted in a more negative direction, i.e. in the solution containing one tenth the normal potassium concentration (0-27
+ 2 0 •-,
0 5 0 0
6 0
-Fig. 10. Legend as for -Fig. 9. KC1 and CaCls are 0-54 and 2 mM respectively.
+20
500 msec.
- 5 0
- 7 0
Fig. i i . Legend as for Fig. 9. KC1 and CaCl2 are 2#7 and 5 mM respectively.
the reversal potential level was measured to be —44-5 mV. (s.E. = ±2-4, n = 5). The change of the reversal potential level produced by a tenfold change of the external potassium concentration was 24-5 mV. in solutions containing 2 mM calcium.
Figures 9-11 show the actual reversal potential levels for the m.e.j.p. measured in sodium-free solution, i.e. Fig. 9 shows the reversal potential level for the m.e.j.p. measured in 2 mM calcium and 2-7 mM potassium by application of the intracellular polarizing currents, in 2 mM calcium and 0*54 mM potassium (Fig. 10) and in 5 mM
Lcalcium and 27 mM potassium solutions (Fig. 11). During these experiments, to
[image:9.451.62.395.136.307.2] [image:9.451.61.401.336.514.2]u 6 Y. ITO, H. KURIYAMA AND N. TASHIRO
prevent the generation of repetitive spikes by the depolarization of the membrane during the application of the currents, the current was applied to the fibre with the minimal gradient. The shift of the reversal potential level in one tenth normal potassium concentration was at 24 mV., and in ten times normal calcium concentration was at 17 mV.
DISCUSSION
i.e. in 2 7 mM (KC1)O, a = 8 in 2 mM (Ca)0; a = 7 in 5 HIM (Ca)0, and a = 7 in 10 mM (Ca)0. These values are mutually in agreement, thus indicating that the permea-bility coefficient of calcium is nearly eight times higher than that of potassium in the presence of acetylcholine. On the other hand, when the external calcium concentration was kept at 2 mM a = 8 in 2-7 mM (K)o as described previously, and a = 5 in 0-54 mM (K)o, and a = 4 in 0-27 mM (K)o. The calculated values varied from 8 to 4. However, the values observed in the various concentrations of potassium indicated that the calcium permeability is much higher than the potassium permeability in the presence of acetylcholine, thus generating the m.e.j.p.s in the depolarizing direction. More information about the mechanism involved on the generations of the m.e.j.p.s will be necessary for further theoretical considerations. It might be possible to conclude from the present experiment that acetylcholine increased sodium, potassium and calcium permeabilities of the longitudinal muscle membrane of the earthworm in physio-logical solution as observed in the frog skeletal muscle (Takeuchi & Takeuchi, 1959, 1960a; Takeuchi, 1963). However, in sodium-free solution generation of the m.e.j.p. is due to increase in calcium permeability and, to a lesser extent, to increase in potas-sium permeability, by spontaneous release of acetylcholine from the nerve terminals.
SUMMARY
1. Miniature excitatory junction potentials (m.e.j.p.s) could be recorded from the longitudinal muscle layer of earthworm in sodium-free solution.
2. The amplitude and frequency of the m.e.j.p.s indicated the diffuse innervation and random release of the chemical transmitter from the nerve terminals.
3. Generation of the m.e.j.p.s was prevented by treatment with D-tubocurarine, but not by atropine and picrotoxin.
4. Hyperpolarizations of the membrane by applications of inward current increased the frequency and amplitude of the m.e.j.p.s in sodium-free solution.
5. The reversal potential level for the m.e.j.p.s in sodium-free solution was — 20 mV., and this value was 20 raV. negative to that measured in physiological solution. Low-potassium solution shifted the reversal potential levels in a more negative and high-calcium in a less negative direction.
6. The change of the reversal potential produced by a tenfold change of the external potassium concentration was 24-5 mV., and that by change of the external calcium concentration was 17 mV.
REFERENCES
BULLOCK, T. H. & HORRIDGE, G. A. (1965). Structure and Function in the Nervous System of
Inverte-brates, vols. 1 and 11. San Francisco and London: W. H. Freeman and Co.
GRUNDFEST, H. (1966). Comparative electrobiology of excitable membrane. In Advances in Comparative
Physiology and Biochemistry, vol 2, pp. 1—116. Ed. O. Lowenstein. New York: Academic Press.
HIDAKA, T., ITO, Y. & KURIYAMA, H. (1969a). Membrane properties of the somatic muscle (obliquely striated muscle) of the earthworm. J. exp. Biol. (in the Press).
HIDAKA, T., ITO, Y., KURIYAMA, H. & TASHIRO, N. (19696). Effects of various ions on the resting and active membrane of the somatic muscle of earthworm. J. exp. Biol. (in the Press.)
HIDAKA, T., ITO, Y., KURIYAMA, H. & TASHIRO, N. (1969 c). Neuromuscular transmission in the
longitudinal layer of somatic muscle in the earthworm. J. Exp. Biol. (in the Press).
n 8 Y. ITO, H. KURIYAMA AND N. TASHIRO
IKEMOTO, N. (1963). Further studies in electron microscopic structures of the oblique-striated muscle of the earthworm, Eisenia Faetida. Biol.J. Okayama Univ. 9, 81-126.
KATZ, B. (1948). The electrical properties of the muscle fibre membrane. Proc. R. Soc. B 135, 506-34. KAWAGUTI, S. & IKEMOTO, N. (1958). Electron microscopy on the smooth muscle of the leech Hirudo
Nipponia. Biol. J. Okayama Univ. 4, 79-91.
NISHIHARA, H. (1967). The fine structure of the earthworm body wall muscle. Ada anat. nippon. 42, 38-9.
TAKEUCHI, A. & TAKEUCHI, N. (1959). Active phase of frog's end-plate potential. J. Neurophysiol. 22,
395-41i-TAKEUCHI, A. & 395-41i-TAKEUCHI, N. (1960a). On the permeability of the end-plate membrane during the action of transmitter. J. Physiol. 154, 52-67.
TAKEUCHI, A. & TAKEUCHI, N. (19606). Further analysis of relationship between end-plate potential and end-plate current. J. Neurophysiol. 23, 397-402.