With 1 plate and 11 text-figures Printed in Great Britain
CO-ORDINATING SYSTEMS AND BEHAVIOUR IN HYDRA
I. PACEMAKER SYSTEM OF THE PERIODIC CONTRACTIONSBY L. M. PASSANO* AND C. B. McCULLOUGHf Department of Biology, Yale University
{Received 16 January 1964)
INTRODUCTION
This is the first of a series of papers on the physiological mechanisms underlying the behaviour of various species of the familiar fresh-water polyp, Hydra. These hydro-zoan coelenterates have long provided favourable material for diverse biological studies (Lenhoff & Loomis, 1961) as well as being a classical representative type for the introductory study of lower invertebrates. Yet it is noteworthy that in spite of their familiarity to biologists much of their biology remains uninvestigated and poorly known. The neurophysiology of coelenterates is of interest both for its own sake and for comparative reasons, since this group contains both animals with the simplest un-doubted nervous system and also forms with a great variety of levels of nervous organization. Nearly all previous findings have been obtained indirectly. Horridge (1954) has pioneered the direct study of the electrical activity of the coelenterate nervous system, using the scyphozoan medusa Aurelia. Other genera of these large jellyfish have been studied extensively by the present authors. With the notable exception of the work of Josephson (1961, 1962) on electrical activity in the athecate hydroids Cordylophora and Tubularia, the present studies are the first to investigate directly electrical activity and its behavioural correlates in hydrozoans.
The behaviour of Hydra, in contrast to its neurophysiology, has been described extensively ever since it was first treated by Trembley (1744) in the eighteenth century. The polyp's ability to contract itself into a tight ball and to extend again has been known ever since the animal was discovered. Earlier accounts were augmented by the observations of Wilson (1891) who described in some detail the phototaxis of Hydra and its affinity for light at the blue end of the spectrum. A report by Haug (1933) ex-tensively discussed contraction behaviour and orientation of the animal to the direction of the light source. Contraction responses to flashes of strong light and to mechanical shaking have recently been described by Rushforth, Burnett & Maynard (1963).
Although several accounts by the present authors of the nature of electrical activity in Hydra have already appeared (Passano, 1963; Passano & McCullough, 1962, 1963), the present paper describes for the first time in quantitative terms a particular aspect of the polyp's behaviour, the contraction burst, and relates it both to specific electrical events occurring within the animal and to various environmental stimuli impinging upon it. This account is intended as an introduction to a series of reports on integrat-ing and co-ordinatintegrat-ing mechanisms in these animals.
MATERIALS AND METHODS
The greater part of our observations were carried out on two species of Hydra, H. littoralis Hyman and H. pirardi Brien. The former was obtained from the Loomis laboratory while the latter, an exclusively male population, is a European species; our animals were obtained from Western Reserve University. A third species was used in a few experiments; it was obtained from a supply house near Northampton, Massa-chusetts and is probably H. cornea Agassiz.
The animals were maintained in the conventional manner in a culture solution made up to 100 mg. NaHCO3 and 50 mg. disodium-diaminetetra-acetate ('versene') in a
litre of tap water (Loomis & Lenhoff, 1956). Glass containers approximately 20 cm. diameter ('finger-bowls') were used, each filled to a depth of 1 cm. and containing 100-200 polyps. They were kept covered and near a north-facing window so that day-lengths experienced by the animals corresponded to the natural conditions of the district. The laboratory temperature varied more or less randomly within 30 C. above or below a mean temperature of about 180 C.; there was probably a diurnal fluctua-tion in temperature as well, but it was not large.
Stocks were fed on newly hatched, washed Artemia nauplii every 3 or 4 days, in excess, and then transferred to fresh solution after half an hour. Under these con-ditions the animals showed continuous growth and asexual reproduction.
To increase the conducting distance, abnormally long specimens were created surgically by cutting down longitudinally from the hypostome almost to the base. The operated animal healed quickly into a double-columned 'two-headed' organism, whose behaviour seemed normal 24 hr. subsequently.
Electrophysiological recording
The microelectrodes used for electrical recordings consisted of saline-filled glass capillaries of conventional form, but of rather lower impedance than those customarily used in intracellular recording. They were gravity-pulled from 'Pyrex' brand hard glass capillary tubing (O.D. 2 mm., I.D. 1 mm.) and then filled with 2-8M-KC1 at room temperature using the differential vapour pressure method. They were stored filled. For use, an electrode was selected, rinsed briefly in culture solution and placed in a sleeve of larger tubing into which a platinum wire was permanently mounted. When the microelectrode was in place the platinum wire extended into and made contact with the saline solution filling the capillary. The support sleeve in turn was held by a sliding micromanipulator (Zeiss—Jena).
Similar electrodes were also used for delivering electrical stimuli, using a Grass S 4C stimulator and SIU 4A stimulus isolation unit. Single pulses lasting 1-5 msec, and of about 1-3 times threshold were used, with a bare platinum wire touching the animal's base as the other stimulating electrode.
platinum wire held just on the surface surrounding the base. It proved helpful if the animal could be induced to fasten an edge of its basal disk to the surrounding ring. The arrangement of manipulators and low-magnification dissecting microscope is shown in Fig. 1. The insert shows an enlarged view of the recording dish with the intact animal, its basal disk on the surface surrounded by the retaining ring.
To polygraph
Fig. 1. The recording arrangement. The animal is suspended from the surface film in a plastic dish and its base is penetrated by one or more recording electrodes supported by a sliding micro-manipulator, while being observed through a dissecting microscope under a safelight (not shown). Localized light stimulation is given with a microscope illuminator fitted with a polished cone of acrylic plastic, painted black to its tip, in another micromanipulator. The entire arrangement is mounted on a heavy steel or concrete plate resting in a box of sawdust. The insert at the upper left shows an enlarged view of the organism.
The microelectrode was brought to the surface of the basal disk at an angle of about 300 from the vertical, while being observed under the dissecting microscope. Usually the tip either just touched or penetrated slightly the basal disk, pressing it against the retaining ring. If a firm tissue-electrode connexion was achieved, and the animal was prevented, by sufficient depth of water, from attaching its tentacles to the bottom of the dish and wrenching away, recordings of several hours duration could be obtained, without further disturbance to the polyp.
Recordings of the pulses associated with contraction bursts could also be made, at greatly reduced amplitude, merely by bringing the recording electrode adjacent to the polyp.
While the recording position at the surface film was preferred for studies of spontaneous activity because it allowed what is an essentially normal position, clear visibility of the animal and an immobile recording site with a minimum of external stimulation, more restrained preparations were also used, especially for dual-channel recordings. The animal was pinned to plates of dental impression wax ('tropical hardness') using the fine barbed spines from the fruit of a species of Opuntia cactus. Prior to any experiment it was necessary to allow the animal at least 30 min. to recover from the excessive stimuli inflicted in setting up the preparation. Handling, illumination, movement of the recording electrode or vibration all had to be avoided. If the recording electrode became dislodged by movements of the animal, a similar recovery period had to be observed after the necessary adjustments had been made, before experimental procedures could be resumed. To reduce vibration as much as possible the dissecting microscope and recording dish containing the polyp were separately supported on the bed of the Zeiss micromanipulator. This in turn rested on a heavy concrete or steel base-plate, weighing approximately 25 kg., imbedded in a sawdust-filled box. The auxiliary micromanipulators also rested on the base-plate. In spite of these precautions, which greatly reduced the vibration transmitted to the animal, all other mechanical disturbances within the laboratory had to be avoided. Most of the results reported here were obtained in a laboratory in an isolated building in quiet surroundings, usually at night. The entire arrangement of manipulators, dissecting microscope and preparation was also within a double-walled shielding cage (Ace Enclosures, Inc.) to reduce RF and AC interference.
Outputs from the recording electrodes and the common reference electrode, a bare platinum wire in one corner of the recording dish 3 cm. from the animal, were fed differentially into high-gain capacitance-coupled Grass P 5 A preamplifiers of a Grass 5 C ink-writing oscillograph, or polygraph. The frequency response of this recording system was flat from 1 to 30 cyc./sec, and off by 3 db. at 70 cyc./sec. An event marker permitted the application of stimuli to be recorded on the paper records. For certain purposes the electrical activity was displayed on a dual channel cathode ray oscillo-scope (Tektronix 502) via two low-level AC preamplifiers (Tektronix 122).
Unlocalized white-light stimuli were given with a battery-fed microscope illuminator (American Optical Co., 'Universal' model) equipped with a Corning Glass heat filter. The light beam passed through a water cell 2 cm. thick and through several centimetres of water in the recording dish before striking the animal. The light was controlled by a hand-operated switch remote from the screening cage. Stimuli of reduced intensity were obtained by interposing neutral density niters between lamp and preparation. Spectrally differentiated stimuli were given by interposing appropriate gelatin filters (Ilford) into the white-light path.
not desired, the preparation was observed under red light (Wratten No. 25). As the animals appear to be insensitive to light of wavelength greater than 600 m/i such illumination allows observation with the dissecting microscope while maintaining 'hydra darkness'. The laboratory was also darkened save for photographic ruby safe-lights during most experiments.
A measuring engine (Fig. 2) was used in this work to obtain exact temporal measure-ments of the occurrences of the various electrical events recorded on the polygraph paper records, in digital form. The apparatus consists of a box with an illuminated viewing surface over which the paper polygraph record is continuously drawn by a
™
<D
<D
d>
10 15
Fig. 2. The measuring engine from above. The device consists of a light box with translucent top, over which polygraph records are pulled between a rubber roller and a steel roller equipped with rubber O-rings. The steel roller is geared to a counter so that i mm. of paper advance is equal to i count. Paper guides are adjustable to accommodate records of different widths. (Covers over gearing and rollers removed for clarity.)
OBSERVATIONS OF BEHAVIOUR AND EXPERIMENTAL RESULTS
Periodic contractions
When undisturbed polyps are observed for extended periods it becomes apparent that Hydra shows more or less regular cycles of activity. These movements are con-siderably faster than those shown by actinians (Batham & Pantin, 1950), yet recording methods that permit the compression of the time scale similarly emphasize the regular occurrence of these events. Examination of the records of electrical activity (correlated with these behavioural changes) over periods of up to 24 hr. or more illustrate this. In the present paper we are concerned with one facet of this continuous behavioural sequence, the periodic co-ordinated contractions of the ectodermal longitudinally arranged muscle sheet of the column. From time to time this muscle shows a series of co-ordinated contractions which reduce the polyp from its usual extended state to a tight contracted ball. All the longitudinal muscles of the tentacles also contract, in approximate synchrony with the column contractions. After shortening progressively to a stumpy spherical shape, the animal again extends, usually in a new direction. This sequence is illustrated in a series of photographs, taken about 10 sec. apart, reproduced as Plate 1.
Between these periodic contractions the animals usually remain extended, seemingly inert, in what appears to be a' fishing' attitude. Occasionally, however, they may take a locomotor step or they may circle slowly about the attached basal disk (Haug, 1933) sweeping the substratum with their tentacles. After 5-10 min. of extension the animal will usually begin to contract again. The longitudinally arranged muscles contract and compress the column down towards the base. The contraction phase typically lasts for 30-60 sec, after which extension, effected by the gastrodermal circular muscles,
follows as before.
These periodic contractions consist of a series of individual contractions, although this fact is not at first obvious from simple observation. Close observation, however, confirms what is easily apparent from recordings of the associated electrical activity, that each contraction consists of a regular succession of individual contractions. The number of contractions is characteristic for each species. We shall return to this point later. Since each ' behavioural' contraction is made up of a series of contractions, we shall refer to the overall periodic contraction as a 'contraction burst'.
In contrast to the step-wise contraction process extension appears to take place in one smooth slow movement. It has been reported (Semal-Van Gansen, 1952) that H. attenuata Pallas extends in a step-wise manner, but although we have looked for this on many occasions in animals of the species we have used, both by simple observa-tion and by time-lapse photomicrography, we have failed to find such extension save, possibly, in a single case. We have been unable to find any electrical correlates to extension.
'Reflex' contractions
are situated. All the animals 'reflexly' contract whenever their bowl is picked up. When the tentacles are removed or when the individual animal is in depression, shak-ing fails to elicit the normal reflex contraction response. It has recently been shown that the thresholds to strong mechanical agitation and to strong light stimuli adapt at markedly different rates (Rushforth et al. 1963).
I M i l l M i l l I I I
I I * I
Simultaneously recorded from two animals in the same dish .
i M i l l I I I I I I M I I
I I I
Pinned and fully illuminated
111 mill 11 mil Minn 1 1111111 nun 1 111 nm M
Illuminated Light off Light on
1111 11 ' i i 1 1 r nm 11
1 1Undisturbed in dark
E
P-I P-I P-I P-I P-I P-I P-I P-I P-I P-I P-I P-I P-I P-I M l
1Hypostome removed I I I I
Hypostome removed I I I I
F I I I 0 1 2
Time (hr.)
Fig. 3. Chronological records of the occurrence of contraction bursts of various individual animals. A! and A2: simultaneous records from two animals (H. littoralis), 2 cm. apart in the
same dish, showing asynchrony of behaviour patterns and phases of excitation and quiescence. B, Hyperactivity by an illuminated and pinned down individual (H. pirardi). C, Effect of illumination on contraction-burst frequency (H. pirardi). At •, locomotor contraction burst occurred and recording was disrupted. D, Undisturbed H. pirardi in the dark (compare with B above). E, Effect of hypostomal extirpation (H. pirardi). F, Same as E, in another species (H. earned).
Spontaneity
Although the contraction bursts of a group of animals may be synchronized by strong external stimuli, the occurrence of a contraction burst is normally inherent, not requiring triggering by any external stimulus. The activities of each of a group of animals in a culture vessel are unsynchronized and particular to each individual. This observation was made repeatedly and without exception. Results of a typical experi-ment with H. littoralis are shown graphically in Fig. 3 Ax and A2. The occurrences of
period, show no co-ordination. It is also evident that the contraction bursts of any individual animal occur periodically but not necessarily at regular or predictable intervals. Intervals between spontaneous contraction bursts are usually irregular and variable rather than rhythmic, although regularity of contraction-burst initiation has been found often enough to suggest that rhythmicity is a valid behaviour pattern for certain individuals at certain (as yet unspecified) times. Usually two or three contrac-tion bursts will take place in the course of 10 min. interspersed with comparable periods of inactivity. Examples of this phasic nature of contraction-burst occurrences can also be seen in Fig. 3.
Other types of contraction behaviour
There are several other behavioural events which involve contractions of the ecto-dermal longitudinal muscles in addition to the normal contraction burst. Single con-tractions comprise approximately 10% of the individual's contraction behaviour. Such single contractions cause only a slight shortening of the animal after which it re-extends and resumes its previous posture without any appreciable activity change. Such contractions could reflect adventitious impulses in the contraction-burst con-ducting system.
A single electric shock given to any one of a variety of the regions of the polyp usually causes a single contraction by the longitudinal musculature rather than a typical contraction burst. These contractions appear to originate from the area stimulated and spread throughout the column. When effective shocks are given 15-30 sec. apart, causing a series of such single contractions, the organism rarely shows regular contraction bursts. Such artificially induced contractions may supplant endo-genous activity for an hour or more. Conventional contraction bursts are more likely to ensue following artificially induced contractions when the animal has been subjected to strong stimulation, either electric shocks of excessive strength, or excessive handling or mechanical agitation.
On certain occasions when individual animals were injured, as by the insertion of cactus spines or by other operative procedures, they exhibited regularly recurring double or triple body contractions in concert at 5-10 sec. intervals for several minutes before returning to normal periodic contraction-burst behaviour (Fig. 7C). Such double contractions at short regular intervals, maintaining the column in a more-or-less tetanically shortened state, give an impression of extreme stress and excitation and last only a few minutes. Since this behaviour was only shown three times it is not known whether some specific parameter of the stimulus led to this behaviour, or whether it was peculiar to certain individuals.
Besides this asymmetrical movement the animals show a variety of simple one-sided contractions of the longitudinal muscles. For example, one quadrant of the longi-tudinal musculature below the hypostome can contract locally, causing the 'nodding' movement which is characteristic during the feeding response. Asymmetrical con-tractions near the base often proceed around the column to cause circling movements. The electrical events associated with these movements appear to be localized in the area concerned and not conducted throughout the animal. Their forms are rather inconsistent.
Diurnal fluctuation in contraction-burst frequency
A variety of environmental variables affect the frequency of spontaneous contraction bursts, and the most important of these is ambient illumination. Contraction-burst frequency under natural daylight is 1*5-2 times that in the dark. Examples of records of contraction bursts in individual animals, in the dark and illuminated, are shown in Fig. 3 B and D. Individual animals show considerable variability in their frequency of contraction bursts, so that the following experiment was carried out using groups of healthy, well-fed hydras (H. littoralis) exposed to daylight from the north sky. During a period of several weeks, observations were made daily for periods of 30 min. each, at 08.00, 13.00 and 22.00. Two observations were carried out at 05.45 (dawn). An area of approximately 25 cm.2 was marked off on the underside of the glass culture vessel. Usually 15-30 individual polyps were found to be attached within this area. Floating animals were not used. The total number of contractions of every animal within this area was counted for the 30 min. period and then the average number of contractions per hour was calculated. The same culture was used throughout the experiment, but, presumably, different animals occupied the count area on different occasions. For any contraction to count as a contraction burst, the animal had to show at least four separate longitudinal contractions and to have shortened to the 'stumpy-ball' stage. Counts at night were observed under a 25 W. ruby photographic safelight, 25 cm. from the vessel. Extreme care was taken to avoid disturbing the culture.
1 5
S 1 0
-t
Fed 0
-o -
o;-
g •
Q
Dawn Noon Sunset Midnight Dawn
Time
Fig. 4. Diurnal variation in contraction-burst frequencies in well-fed, naturally illuminated H. littoralis. Points represent mean values for groups of approximately 25 individuals (see text).
[image:9.451.64.391.440.609.2]652 L. M. PASSANO AND C. B. MCCULLOUGH
The results of this experiment are shown graphically in Fig. 4 and Fig. 5 a. Con-traction burst frequency rose from the lowest value prior to dawn to the highest value soon after sunrise. This high level of contraction-burst activity continued for several hours and then commenced to decline slowly during the day. By late evening activity had declined to approximately one-half of that of the morning levels.
That this diurnal variation in contraction-burst frequency is due to ambient illumination levels is confirmed by counts made on a control culture of H. littoralis maintained in the same manner save that they were continuously illuminated. No residual circadian rhythm was evident under these conditions.
A. Fed daily
L
Contraction bursts per hr.
14-1 + 0-7
11-8+0-7
7-2 + 0-5
B. Fed every third (or fourth) day
r - - - • • :
First day 7-8 + 0-4
6-6 + 0-4
5-1 + 0-4
Second day 7-5+0-3
6-5 + 0-4
4-9 + 0-4
Third day
6-8 + 0-4
6-0 + 0-5
4-8 + 0-5
Fourth day
5-4 + 0-4
4-4 + 0-5
Fig. 5. Contraction-burst frequencies in relation to time of day and feeding regime, for naturally illuminated H. littoralis. Mean values with s.E. were obtained in the morning (light bar), afternoon (medium bar) and evening (dark bar), see text. A, Fed daily after the afternoon count. B, Fed every third (or fourth) day, showing depressed contraction-burst frequency and further reduction on days following feeding.
Starvation
Animals starved longer than a week show little if any contraction-burst activity; in fact almost all overt behaviour of any kind ceases.
The 'light response' to sudden illumination
Besides the daily fluctuation in contraction-burst frequency due to natural daylight illumination, there is the immediate, and dramatic, response of the dark-adapted culture to abrupt, strong illumination. This behavioural sequence has already been described (Passano & McCullough, 1963).
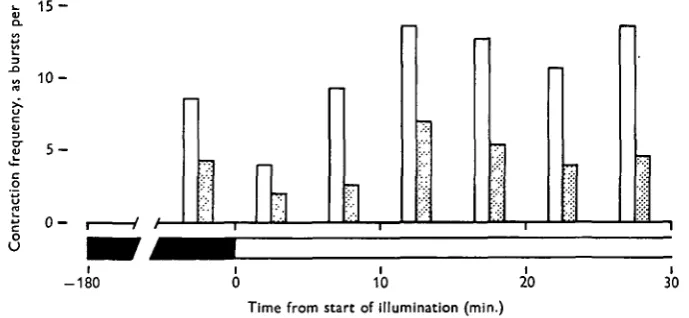
A culture of H. littoralis such as described above was given natural daylight illumination for some days. After the 23.00 hour count in the dark (i.e. 'hydra darkness' using a ruby safelight), the culture was illuminated with a white 60W. incandescent lamp 25 cm. from the culture vessel. Contraction bursts were counted, and mean frequencies were determined, for each 5 min. period during the following 30 min. These results are shown in Fig. 6, which also gives the results of a parallel
1 5
1 0
5
0
-10
Time from start of illumination (min.) 1
[image:11.451.54.399.271.430.2]20 30
Fig. 6. The effect of illumination on contraction-burst frequency in well-fed (open bar) and starved (filled bar) H. littoralis. Frequency rises 75 % following initial inhibition.
experiment with an unfed culture of the same species. Temperatures were not con-trolled in these experiments, but the rise in temperature in the culture vessel was no more than 0-3° C. after 30 min.
The response to sudden illumination may be summarized:
(1) immediate cessation of contractions upon illumination. A contraction burst in progress is abruptly halted;
(2) extension of the column to twice the length of dark-adapted animals; (3) after 3-6 min. either (a) a contraction burst, or (b) locomotion;
(4) contractions (or locomotion; seen more commonly in starved animals) recur at approximately twice the frequency of spontaneous contraction bursts in the dark, for the next 20 min.;
(5) activity declines after 30 min., but remains above dark-adapted levels if illumination is continued.
This light response will be discussed in more detail in a subsequent paper.
654
Electrical correlates of the contraction burst
By arranging an animal in the manner described above (under Materials and Methods) it is possible to obtain records of the electrical changes occurring within the animal for extended periods without serious interference with its behaviour. By simultaneously recording and observing the animal (using red light as ' hydra darkness' when appropriate), it can be shown that whenever the animal undergoes the series of longitudinal contractions making up the contraction burst, each contraction is pre-ceded by a large (30 mV.), slow (250-500 msec.) compound potential. After these pulses cease the polyp slowly re-elongates.
Several examples of polygraph records of such preparations, using H. littoralis, are shown in Fig. 7, and comparable records obtained from H. pirardi are given in Fig. 8.
• \ \
Fig. 7. Polygraph records of impulses from individual H. littoralis. A, Consecutive regular contraction bursts (portion of interburst record deleted); c c indicates electrical poten-tials which preceed contractions. B, Locomotor contraction burst, with an attempt to wrench away the pedal disk; note the greater number and frequency of potentials, c' c', associated with these contractions. C, Double potentials (c) found with rhythmic contractions following injury with cactus spines. • , 'Rhythmic potentials' (Passano & McCullough, 1962) conducted on separate pathways, a, Asymetrical contraction potential, not through-conducted. / /, Special burst of rhythmic potentials associated with tentacle attachment and pre-locomotor behaviour, u, Contraction potentials associated with asymmetrical uncoiling (see text). Scales at lower left: vertical; 100/tV., horizontal; 10 sec.
It should be noted that the polygraph recording speed used in the records of H. pirardi is less than that used for the records of H. littoralis.
much more rapid and contain more pulses. We shall return to these patterns in a later section of this paper.
[image:13.451.71.390.235.426.2]Individual pulses are similar and recognizable. There is a rapid initial deflexion, negative with respect to the reference electrode, followed by a slower positive swing and subsequent return to the base line. A more precise determination of the un-distorted shapes of these potentials has not been made; since we were simultaneously interested in other electrical activity of a much lower level our records give a distorted picture of these pulses. In a given recording situation, however, superimposed cathode ray oscilloscope traces of the consecutive contraction impulses in a burst did not disclose any notable variation in the form, amplitude or duration of the pulses. Such determinations are not very satisfactory because the animal's movements nearly always result in some movement of the recording electrode tip relative to the impaled tissue.
Fig. 8. Polygraph records of impulses from individual H. pirardi. A, Regular endogenous contraction bursts. B, Locomotor contraction burst. Lower pen shows time in min. • , Vertical scale: rhythmic potential impulses approximately ioo /tV. Symbols as in Fig. 7.
The conduction velocity of the contraction pulse was difficult to measure exactly because of the small size of the polyp and the slow rise-time of the impulse. To achieve a longer conducting path, a two-headed, double columned H. pirardi, joined at the base, was created surgically in the manner described above. The animal thus prepared was pinned horizontally to the bottom of the preparation dish and recording electrodes were inserted near the tentacle insertions of the two 'heads'. Spontaneous contraction bursts arose at either end indiscriminately, seemingly at a normal fre-quency for a restrained preparation (see Fig. 3B). The mean conduction velocity obtained from this preparation was 15 cm./sec, which is approximately what has been found in normal animals when recording simultaneously from two points on the column.
Point of origin of potentials
confirm this localization by other means. Two methods have been used: extirpation and localized light stimulation.
Removal of the entire hypostomal region (and also the tentacles) by a transverse 'decapitation' proximal to the point of insertion of the tentacles eliminates all co-ordinated contractions of the longitudinal muscles for 45-60 min., or even longer. Cutting the column and base, etc., does not appear to affect contraction bursts unless the hypostome is removed. Removal of a few or all of the tentacles does not affect endogenous initiation of contraction bursts, although the polyps do then become in-sensitive to strong mechanical stimuli, such as that caused by disturbing the culture dish, which causes 'reflex' contractions in intact animals. Our findings here confirm those of Rushforth et al. (1963).
After about an hour of inactivity contractions recommence, but at a greatly reduced frequency. The effect of hypostomal removal is shown diagramatically in Fig. 3E and F. Burst patterns are initially abnormal. Contraction-burst frequencies remained permanently depressed, at 2-3 per hour, in cases where the tentacles and hypostome failed to regenerate. Otherwise the burst frequency was again normal in 2-3 days.
Localized light stimulation provides a more precise method of localizing the burst-initiation area. As previously noted, the initial effect of illuminating a dark-adapted animal is to bring about a cessation of concentrations. Contraction bursts under way are interrupted (see following paragraph). With the use of the plastic cone attachment to a microscope illuminator, described in the previous section of this paper, it proved possible to give localized light stimuli to circumscribed regions. A number of such experiments were carried out, using animals either floating on the surface film or restrained by being pinned to wax plates. The former preparations showed more nearly undisturbed behaviour but were harder to stimulate accurately. Such experi-ments all showed that illumination of the hypostomal region alone could halt contrac-tion bursts. Contraccontrac-tions were not affected by illuminating the column below the point of tentacle insertion, nor anywhere further down the column to the basal disk.
Interruption of contraction bursts by light
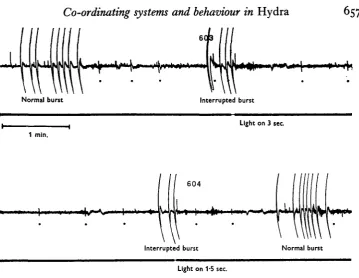
Results of further experiments on the interrupting of the contraction burst by strong light stimulation may be mentioned briefly at this point, although this aspect of the investigation requires further consideration. General illumination with a micro-scope lamp could halt a contraction burst in about 90% of all animals tested. In exceptional cases bright flashes of no more than 0-2 sec. duration were consistently effective. An example taken from such experiments is shown in Fig. 9, where following a series of normal bursts, two successive bursts were halted. The next burst was un-interrupted, showing that bursts of normal pattern follow such interruptions. Very short, intense, flashes given by photographic flashbulbs were ineffective even when given to the most sensitive individual polyps. Reducing the intensity of the light stimulus leads to variable responses; often contractions are slowed down rather than halted completely, so that there were abnormally long mid-burst intervals after which the burst continued its normal course. In other cases the inhibitory effect was delayed, one pulse occurring regularly after the stimulus was given.
,.+\,i fu.
6C
657
Normal burst Interrupted burst
Light on 3 sec.
1 min.
604
Interrupted burst
\ \\w\\\
Normal burst [image:15.451.48.408.45.323.2]Light on V5 sec.
Fig. 9. Continuous polygraph record from dark-adapted H. pirardi. Contraction bursts are twice halted by short periods of general illumination, as shown on the lower trace. # , 'Rhyth-mic potentials' (Passano & McCullough, 1963) conducted on separate pathways.
other effects of light on Hydra (Passano & McCullough, 1962). Even yellow-pass and green-pass filters (at the intensities available to us) failed to influence bursts in progress. Blue light (wavelength shorter than 500 va/i) and blue-violet light consistently showed the same efficiency as white light in halting contraction bursts even at energy levels much reduced from that available from the unfiltered source, for threshold intensities were not appreciably altered by the interposition of these pass filters into the white-light beam.
Contraction-burst pattern
6
58
L. M. PASSANO AND C. B. MCCULLOUGHSeveral aspects of the patterned nature of contraction bursts can be seen from such graphic representation:
(1) In both species the occurrence of pulses in the regular contraction bursts accelerates and decelerates at a fairly uniform and predictable rate.
(2) Contraction-burst pulses are normally not generated more frequently than about one per sec. The shortest interval, usually occurring near the middle of the burst, is found to be between 1 and 2 sec. Exceptions to this characteristic refractory period in normal bursts are the double or even triple potentials occasionally encountered (Fig. 7C ) .
Regular bursts
Locomocor bursts
30 sec. 30 sec.
Fig. 10. Graphic representations of individual contraction bursts, illustrating the two different patterns of pacemaker output (associated with regular and locomotor contraction bursts) in each of two species of Hydra, H. pirardi (above) and H. littoralis (below). Intervals between contractions are plotted vertically, and the shortest interval of each burst has been positioned centrally on the ordinate.
(3) Despite variations, pattern differences between species are readily apparent. In seemingly identical situations contraction bursts of H. pirardi last less than 30 sec. and consist of 5—8 pulses, whereas in H. littoralis contraction bursts last about 45-60 sec. and contain 9-12 pulses. Visually observed individuals of both species in undisturbed culture vessels showed the same species characteristics as those shown under recording conditions.
The greater excitation of the locomotor contraction burst is also demonstrated by the fact that light stimuli of intensities and durations sufficient to halt an animal's regular contraction burst were ineffective when applied during locomotor bursts.
Records from unpinned animals showed the same characteristics as those from the more restrained preparations. In several cases the animal attempted to attach its tentacles to the surface film, although when the somersault was attempted the tentacles lost their grip and the recording site was not dislodged. Even when the animal succeeded in its locomotion attempt, the pattern recorded before the interruption resembled that already described.
The electrical events and the contraction
Recordings were carried out on a series of H.pirardi in culture solution to which 1 o~2 M chlorotone solution was added slowly from time to time. Each animal was observed by one of us while the other recorded the observations on the polygraph record. Such experiments show that as the concentration of chlorotone increased to about io~3M (10% of the lethal concentration under these conditions), contractions of the longi-tudinal musculature became very lethargic. In response to each impulse the column shortened only slightly rather than in its normally vigorous fashion, so that at the completion of the burst the polyp was no more than one-half to two-thirds contracted. The tentacles did not contract at all with the column, but remained inert and more or less rigidly extended. The electrical burst pattern was entirely unaffected by the chlorotone treatment. After being returned to fresh culture solution, contractions returned to normal within a few minutes. Essentially similar results have also been obtained in experiments using H. canadensis Rowan.
Spontaneity and the endogenous nature of contraction bursts
In the light of the accumulated evidence that contraction-burst potentials are generated by a unit or units made up of light-sensitive elements and that these potentials are independent of the actual process of contraction, it was of interest to learn more about the properties of this 'unit'. The generative element shows charac-teristics usually ascribed to a 'pacemaker'.
Certain of our experimental results had suggested that individuals tended to increase their contraction-burst frequency when these contractions were repeatedly blocked by light, as described above. Systematic inquiry showed that this was so. For example: an intact full-sized H. littoralis was allowed to dark-adapt undisturbed for an hour. For the succeeding 30 min. period every contraction burst was given an inhibiting light stimulus consisting of 5 sec. of general bright illumination immediately after the first spontaneous contraction pulse occurred. Following a further 30 min. period without stimulation, the process was repeated save that three impulses were permitted prior to the inhibiting stimulus. Another 30 min. period of recovery was allowed and then the light-inhibition was carried out yet a third time, in which two contraction pulses were permitted.
normal spontaneous contraction bursts was obtained from the latter 30 min. of the initial pre-experimental period, but comparable levels were attained during the inter-mediate recovery periods. It is noteworthy that, although all of the 'blocked' periods showed greatly heightened frequencies, bursts were initiated most frequently when only a single pulse was permitted and that even where two were allowed the burst frequency was slightly higher than that for three.
294 ±42 sec.
41 ±10 sec.
B
2 | I 58 ± 4 sec.
6S±4sec.
1
0 1 2 3 4 5
Time between contraction bursts (min.)
Fig. I I . The effect of repeated interruption of contraction bursts by illumination on the interval between contraction bursts. A: control: mean interval (and s.E.) for consecutive uninterrupted contraction bursts, with 8-10 contractions per burst. B^ B2, and Ba, Mean
intervals for consecutive bursts interrupted after one (BO, two (Ba) and three (B3)
con-tractions. The interval length between bursts is directly related to the number of contractions permitted to occur in the previous contraction burst.
When consecutive contraction bursts are halted in this manner, blocking thresholds gradually rise, until eventually a contraction burst proceeds in spite of the inhibitory stimulus. In the experiment just described, 12-8% of the blocking stimuli were ineffectual during the one-pulse period and 7-6% ineffectual during the two-pulse period; only in the period where the light stimulus was given after the third sponta-neous pulse was the light inhibition 100% effective.
The same enhancement of contraction-burst frequency occurs even if the inhibiting light stimulus is given locally to the hypostome rather than to the entire animal.
DISCUSSION AND CONCLUSIONS
these animals' behaviour. Activities in Hydra, as in other coelenterates and in many sessile forms, require prolonged observation to reveal their behaviouristic significance. Reliable recording techniques, although involved with pathway areas and entire nerve-nets rather than single neurons or bundles of axons, provide the means by which correlations between behaviour and electrical activity may be ascertained.
The periodic contractions of Hydra are characterized as much by their variability of their occurrence as by their repetition. This may make the study of them more difficult, but it strengthens our belief that contraction bursts are a visible manifestation of the complex integrated behaviour of the organism. Such variability is always characteristic of an animal's behaviour. The repeated observation of asynchronous contraction bursts among individual polyps in a common undisturbed environment shows that bursts are initiated endogenously. Yet contraction-burst frequencies are very sensitive to mechanical disturbances in the environment and to ambient illumination levels. The adaptive significance of both of these responses for a more or less sessile, slow-moving carnivore is obvious. The circadian rhythm of activity imposed by the effect of illu-mination levels on burst frequency gives polyps a cyclic activity to correspond to that shown by their prey, mainly crustacean plankters, whose activity rhythm in turn is synchronized to their phytoplankton food source.
Contraction bursts are also used as a component of Hydra!s locomotor sequence. Quantitative study of the patterns of the individual bursts in two species of Hydra show that such locomotor bursts are distinct modifications of the regular burst. The polyp's distinctive somersaulting locomotion has thus been achieved by evolving a behavioural sequence of individual components adapted and modified from their more usual form. Both regular and modified patterns exist simultaneously.
Turning from the behavioural to the physiological level of inquiry shifts attention to the mechanisms controlling the individual contraction burst. Is the patterned sequence of longitudinal muscle contractions, that makes up such a burst, each preceded by a through-conducted electrical pulse a patterned output from an auto-nomous pacemaker component of the co-ordinating system? Alternatively, is a burst simply an automatic chain-reflex response to proprioceptive feedback from internal stretch, engendered by a single random contraction?
Experiments with chlorotone anaesthesia show that the electrical pulse train occurs independently of muscle timing and of the extent of shortening. Furthermore, roughly 10% of all spontaneous contraction behaviour consists of single contractions, without a patterned burst ensuing. Single electric shocks cause single contractions, not bursts, with few exceptions. If the burst components were simply linked mechanically, we presume that the entire sequence would inevitably be set off by the initial event and that the sequence would be unvarying and predictable in much the way that a line of bowling pins falls over when the first is toppled. This does not in fact occur. The only component of the contraction burst that might be mechanically linked is the co-ordination between column contraction and tentacle contractions; here chlorotone anaesthesia does unlink these components of the response pattern.
The two different contraction-burst patterns—the regular burst and the locomotor burst—also argue against the 'chain-reflex' explanation of the burst. It is difficult to imagine the same musculature bringing about two, distinctively different, contractions without postulating the existence of a centre for integrating various sensory stimuli and effecting a co-ordinated response appropriate to these conditions. The problem of locomotion in Hydra will be considered in another paper; however, it is noteworthy that moderately starved animals show more locomotion than do well-fed polyps, although the latter show a higher frequency of regular contraction bursts. Sudden stimulation, even that localized on a portion of the animal not directly concerned with contraction bursts (Passano & McCullough, 1963), often causes an animal to show one or more locomotor contraction bursts. It seems likely that such sensory information has been processed either indirectly or directly through a pacemaker unit which, in turn, is responsible for the patterned contraction-burst pulses and through them the resulting muscular contractions.
The properties of the contraction-burst pacemakers may be summarized as follows: (1) They are localized in the hypostome or immediately adjacent to it. This has been shown by extirpation, dual-recording and localized stimulation experiments.
(2) They are endogenously active and not dependent upon some subtle external stimulus; they are, however, affected by the internal state of the organism and by external stimuli of various kinds.
(3) They are directly inhibited by light, with the blue wavelengths of the spectrum most effective. As no specialized light-receptor structures, at any morphological level, are as yet known in Hydra, this sensitivity falls into the general category of ' dermal light sense'. It suggests, but does not prove, that the pacemakers are neurons.
(4) There are two (kinds of) pacemakers, those producing regular contraction bursts and those causing locomotor contraction bursts. They are differentially affected by nutritional status, and perhaps by ambient illumination.
(5) Output from one pacemaker suppresses that of others. This could be seen in the 'two-headed' animals created surgically, and even by the effect of suppression by repeated electric shocks.
(6) Prevention of regular pacemaker output, as by the application of inhibiting flashes of light, drastically increases the pacemaker firing frequency. It can also decrease its sensitivity to light inhibition.
pacemaker system under 'higher' control. Nevertheless, contraction-burst fre-quencies afford a simple and quite reliable means of assessing the individual polyp's 'psychological state'. They are easy to monitor electrically and are clearly visible to the observer. In contrast to the slow behaviour shifts of Metridium (Batham & Pantin, 1950), the periodic contractions of Hydra recur at a lively rate. They provide a clear quantitative measure of behaviour phase. The sensitivity of their pacemakers to flashes of light provides an easily manipulated tool with which to vary the animal's state of excitation. These properties of the contraction-burst system will be of aid in attempting to understand the relationship between the contraction burst and the rhythmic potential systems.
SUMMARY
1. A technique of recording electrical activity from an intact, essentially normal specimen of Hydra is described.
2. The existence of regularly recurring co-ordinated longitudinal contractions of the ectodermal muscles is confirmed and re-emphasized.
3. Such contractions are found to consist of a patterned series of individual co-ordinated contractions, each preceded by a single large, compound potential. The overall contraction, consisting of a variable number between 5 and 12 or more con-tractions depending on species, is thus called a contraction burst.
4. These contraction-burst potentials originate endogenously; they are considered to be the most important effector activity in Hydra.
5. Contraction-burst potentials originate in the hypostome and are conducted throughout the column at approximately 15 cm./sec.
6. Contraction-burst patterns have been studied quantitatively in two species, showing interspecific differences between both regular contraction bursts and those associated with locomotion.
7. Certain extrinsic and intrinsic variables affect contraction-burst frequency. Day-light, and nutritional state, both modify this rate, with the former giving rise to a circadian activity cycle under natural conditions.
8. Single electric shocks usually cause a single co-ordinated muscle contraction. Such stimuli can markedly reduce endogenous contraction-burst activity.
9. Sudden illumination interrupts contraction bursts temporarily, even halting those in progress. Blue light is most effective. This stimulus has been used as a tool to investigate the properties of the pacemaker systems concerned with contraction bursts.
10. The nature and properties of these pacemakers is discussed.
664 L. M. PASSANO AND C. B. MCCULLOUGH
REFERENCES
BATHAM, E. J. & PANTIN, C. F. A. (1950). Phases of activity in the sea-anemone Metridium senile (L.), and their relation to external stimuli. J. Exp. Biol. Z7, 377-99.
HAUG, G. (1933). Die Lichtreaktionen der Hydren. Z. vergl. Physiol. 19, 246-303:
HORRIDGE, G. A. (1954). The nerves and muscles of medusae. I. Conduction in the nervous system of Aurellia aurita Lamarck. J. Exp. Biol. 31, 594-600.
JOSEPHSON, R. K. (1961). Repetitive potentials following brief electrical stimuli in a hydroid. J. Exp. Biol. 38,
579-93-JOSEPHSON, R. K. (1962). Spontaneous electrical activity in a hydroid polyp. Comp. Biochem. Physiol. 5, 45-58.
LENHOFF, H. M. & LOOMIS, W. F. (1961). The Biology of Hydra and some other Coelenterates. Coral Gables: University of Miami Press.
LOOMIS, W. F. & LENHOFF, H. M. (1956). Growth and sexual differentiation in Hydra in mass culture. J. Exp. Zool. 13a, 555-74.
PASSANO, L. M. (1963). Primitive nervous systems. Proc. Nat. Acad. Sci., Wash. 50, 306-13. PASSANO, L. M. & MCCULLOUGH, C. B. (1962). The light response and the rhythmic potentials of Hydra.
Proc. Nat. Acad. Sci., Wash. 48, 1376-82.
PASSANO, L. M. & MCCULLOUGH, C. B. (1963). Pacemaker hierarchies controlling the behaviour of hydras. Nature, Land. 199, 1174-5.
RUSHFORTH, N. B., BURNETT, A. L. & MAYNARD, R. (1963). Behaviour in Hydra: contraction responses of H. pirardi to mechanical and light stimuli. Science, 139, 760.
SEMAL-VAN GANSEN, P. (1952). Note sur le systeme nerveux de l'hydre. Bull. Acad. Belg. Cl. Sci., V, 38,
718-35-TREMBLEY, A. (1744). Mhnoirespour servir dl'histoire d'ungenre de polypes d'eau douce a bras en forme de comes. Leyden: J. and H. Verbleck.
WILSON, E. B. (1891). The heliotropism of hydra. Amer. Nat. 25, 413-33.
EXPLANATION OF PLATE