DOI: 10.1534/genetics.104.027631
Rapid Evolution Through Gene Duplication and Subfunctionalization of the
Testes-Specific
␣
4 Proteasome Subunits in Drosophila
Dara G. Torgerson
1and Rama S. Singh
Department of Biology, McMaster University, Hamilton, Ontario L8S 4K1, Canada
Manuscript received February 13, 2004 Accepted for publication July 29, 2004
ABSTRACT
Gene duplication is an important mechanism for acquiring new genes and creating genetic novelty in organisms. Evidence suggests that duplicated genes are retained at a much higher rate than originally thought and that functional divergence of gene copies is a major factor promoting their retention in the genome. We find that two Drosophila testes-specific␣4 proteasome subunit genes (␣4-t1 and␣4-t2) have a higher polymorphism within species and are significantly more diverged between species than the somatic ␣4 gene. Our data suggest that following gene duplication, the␣4-t1 gene experienced relaxed selective constraints, whereas the␣4-t2 gene experienced positive selection acting on several codons. We report significant heterogeneity in evolutionary rates among all three paralogs at homologous codons, indicating that functional divergence has coincided with genic divergence. Reproductive subfunctionalization may allow for a more rapid evolution of reproductive traits and a greater specialization of testes function. Our data add to the increasing evidence that duplicated genes experience lower selective constraints and in some cases positive selection following duplication. Newly duplicated genes that are freer from selective constraints may provide a mechanism for developing new interactions and a pathway for the evolution of new genes.
G
ENE duplication is a source of new genetic function functionalization), and therefore beneficial mutations would not be necessary to retain duplicate copies. and a mechanism of evolutionary novelty (OhnoFunctional divergence may occur through a variety 1970). The classical model of gene duplication is that
of evolutionary processes, including relaxed selective one copy is free to evolve neutrally, thereby
accumulat-constraints, neutral evolution, or even positive selection. ing random mutations that may infrequently result in
Examples of positive selection acting after gene duplica-the uptake of a new function (Ohno1970). Under this
tion include the MADS-box gene family in Arabidopsis model the majority of duplicated genes will be lost, as
(Martinez-CastillaandAlvarez-Buylla2003), xan-most copies will not accumulate the proper combination
thine dehydrogenase (Rodriguez-Trelleset al.2003), of neutral mutations to regain a novel function.
How-CCT proteins (FaresandWolfe2003), phytochrome ever, the generalized fate of duplicated genes is under
A in angiosperms (Mathewset al.2003), and numerous increasing scrutiny, and there is mounting scepticism
others (Ohta1994;Zhanget al.1998, 2002;Johnson that the classical model of gene duplication is able to
et al. 2001;Betra´nandLong2003). Even if acting on explain the large number of duplicated genes that are
a small number of sites for a brief period of time, positive retained in a genome (Massingham et al.2001). The
selection may be an important factor in retaining dupli-duplication-degeneration-complementation (DDC)
cated genes by promoting the acquisition of a novel or model as formally proposed byForceet al.(1999) along
more specialized function. Therefore subfunctionaliza-with similar ideas (Hughes1994;Stoltzfus1999) offer
tion can occur by degenerative mutations resulting in an alternative explanation of the evolutionary fate of
the loss of some functions through either neutral or duplicated genes. If an ancestral gene carries out more
less-constrained nucleotide substitutions and positive se-than one function and undergoes a duplication event,
lection acting on one gene copy can result in a more degenerative mutations could result in each copy
be-specialized function, which together may be significant coming specialized in alternative functions (termed
sub-factors preventing the loss of duplicated genes. Gene duplication allows the study of evolutionary pro-cesses, but also provides an opportunity to examine the evolutionary history of multiunit protein complexes that Sequence data from this article have been deposited with the
EMBL/GenBank Data Libraries under accession nos. AY542377– have arisen through gene duplication. One such exam-AY542432.
ple is the proteasome, which is responsible for
degrad-1Corresponding author:Department of Biology, McMaster University,
ing proteins intracellularly in a highly regulated and 1280 Main St., West Hamilton, ON L8S 4K1, Canada.
E-mail: [email protected] specific manner. Proteasomes play a critical role in many
basic cellular pathways and are important for regulating evolving rapidly has been found (Singh 1990; Singh andKulathinal2000;SwansonandVacquier2002). most biological processes that take place in an organism
(seeGlickmanandCiechanover2001 for review). The For example, in Drosophila genes expressed in the re-productive tract show higher divergence than those that eukaryotic proteasome is a large 26S multicatalytic
pro-tease composed of two subcomplexes: a 20S core parti- are not (Coulthart and Singh 1988; Civetta and Singh1995), and accessory gland proteins are among cle and a 19S regulatory particle. The core particle is
barrel shaped and is composed of four stacked rings: two the fastest-evolving genes in the Drosophila genome (TsaurandWu1997;Tsauret al.1998;Swansonet al.
identical outer ␣-rings and two identical inner -rings,
each composed of seven distinct subunits. In Archae 2001). Similarly, in mammals some of the more highly diverged proteins are those found in sperm compared the overall structure of the core particle is conserved
with that of eukaryotes; however, it contains only two to other tissues (Torgersonet al.2002), and in Chlamy-domonas sex-related genes are evolving faster than distinct subunits: seven identical ␣-subunits and seven
identical-subunits. Over time the proteasome has be- genes involved in other processes (Ferriset al.1997). Rapidly evolving reproduction-related genes are also come increasingly complex, and the genes coding for
the␣- and-subunits appear to have undergone many found in a variety of other taxa, including centric diatoms (Armbrust andGalindo2001), gastropods (Hellberg gene duplications and specialization leading to at least
14 distinct genes coding for proteins in the eukaryotic et al.2000), abalone (SwansonandVacquier1995), and humans (Wyckoffet al.2000).
core particle (seven unique␣-subunits and seven unique
-subunits). This appears to have occurred early during Due to the large body of evidence that reproduction-related genes evolve rapidly compared to genes not the evolution of Eukarya for the ␣-subunits, resulting
in a variable number of␣proteasome genes across bac- directly involved in reproduction, we hypothesized that the ␣4-t1 and␣4-t2 genes may have their own unique teria, archaea, and eukarya (Bouzatet al.2000).
In Drosophila the proteasome is even more complex evolutionary pathways relative to that of the somatic␣4 gene. Moreover, given that gene duplication can result than initially thought, as several subunits have isoforms
with testes-specific expression patterns (Maet al.2002). in changes in selective constraints that can ultimately lead to functional divergence, we hypothesized that re-The␣4 subunit has undergone at least two gene
duplica-tion events resulting in three paralogs with distinct tissue productive specialization had an effect on the evolution of the␣4 proteasome gene family. In this study we com-expression patterns (Yuanet al.1996). The majority of
cells in Drosophila express the somatic ␣4 gene; how- pare the polymorphism and divergence of the␣4 gene family in Drosophila, test for differences in selective ever, in the testes two different tissue-specific genes (␣
4-t1 and␣4-t2) have replaced the␣4 subunit in the core constraints acting on the testes-specific paralogs, and identify codons among genes that may show functional particle of the proteasome. On the basis of phylogenetic
analysis (Beloteet al.1998), it seems that the first dupli- divergence. cation involved the testes-specific ␣4-t2 gene and the
somatic␣4 gene, followed by a more recent duplication
MATERIALS AND METHODS of the somatic␣4 gene and a second testes-specific gene,
␣4-t1. InDrosophila melanogasterall three duplicated genes Fly strains:Seven lines ofD. melanogasterwere sequenced: one from Hawaii (0231.0) originally from the Bowling Green reside on separate chromosomes, with the somatic␣4
Species Stock Center; one from Peru (0231.1) from the gene on the X chromosome and the two testes-expressed
Bloomington Stock Center; two from Pennysylvania (CPA-46, genes on chromosomes 3 and 2 (Yuan et al. 1996). CPA-129) from Brian Lazzaro at Cornell University; and three At some time following gene duplication both testes- from Zimbabwe [Z(H)-12, Z(H)-16, Z(H)-34] provided by the Andrew Clark laboratory, now at Cornell University, which expressed paralogs became specialized in reproductive
were originally from David Begun at the University of Califor-functioning and are expressed at different times during
nia, Davis. SixDrosophila simulanslines were sequenced: one spermatogenesis. The␣4-t1 subunit is expressed at the
from Madagascar (S-24) and one from Ethiopia (S-23), both primary spermatocyte stage and into spermatid elonga- from John Roote at Cambridge University; one from Florida tion, whereas the␣4-t2 subunit is expressed only during (0251.166), one from an unknown location (Solway-Hochman 1088), and one from Australia (0251.4) from the Bowling spermatid elongation (Yuan et al. 1996). It has been
Green Stock Center; and one from Italy (S-132) from the speculated that during spermatogenesis proteasomes
Umea Stock Center in Sweden. Four Drosophila mauritiana recognize and degrade discarded proteins and regulate
lines were sequenced: two from the Bowling Green Stock the fine structural tuning of the sperm tail (Beloteet Center (0241.1, 0241.7), and two from the Umea Stock Center al.1998). (S-80, S-81). Three lines of Drosophila sechellia, all from the Bowling Green Stock Center (0248.3, 0248.7, 3151), were se-Reproduction-related genes such as␣4-t1 and␣4-t2
quenced. may be subject to alternative evolutionary forces, such
DNA extraction, PCR amplification, and sequencing:Five as sexual conflict and sexual selection, which may
was redissolved in 30l of ddH2O and stored at⫺20⬚until equal todS, then the gene is thought to be under neutral evolution, and will be close to 1. In scenarios where dN PCR amplification.
Primers for PCR amplification were designed using theD. actually exceedsdSandis significantly⬎1, the gene is said to be under positive selection.
melanogastergenome, and in most cases were designed in the
5⬘and 3⬘noncoding region to allow for complete amplifica- We compared the likelihood of different models of selec-tion acting on different branches of a phylogenetic tree using tion of the coding regions of the gene, including the two
introns in both␣4 and␣4-t2. The primer pair used to amplify the program codeml in the PAML package version 3.13 (Yang 1997). This program utilizes the codon substitution model the␣4 gene in D. melanogaster, D. simulans, D. sechellia, and
D. mauritianawas 5⬘-TGCCTGGCGAATTCGAGAAGG-3⬘and of Goldman and Yang (1994) and a maximum-likelihood method to calculate the likelihood of specified models. Twice 5⬘-GTCGCCGAATGCATGGAAAGC-3⬘. The primer pair used
to amplify␣4-t1 was 5⬘-TGCCTGCTAACTAACCCAAAG-3⬘and the difference in likelihoods of two models is then compared to a chi-square distribution, with the degrees of freedom equal 5⬘-GTACCTGCTATCCTGGGTGAC-3⬘. For the␣4-t2 gene, two
primer pairs were designed (external-internal and internal- to the difference in the number of free parameters between the two models. A phylogenetic tree was also generated using external): 5⬘-CCAGTACGCACCTAGCAGGCG-3⬘and 5⬘-ACA
GGACAATCCAAATGGACG-3⬘, and 5⬘-CTGAATTTCGAGAA a single individual of each species (see Figure 1) to avoid polytomy and to avoid estimations of based on polymor-GCCCACG-3⬘and 5⬘-GAACAGAATGGATCAGGGTGG-3⬘. PCR
products were purified using a min-elute QIAGEN (Chats- phism vs.divergence (Figure 2). Trees were also generated with maximum parsimony and neighbor-joining using Kimura worth, CA) gel PCR clean-up kit and sequenced using an ABI
377 Prism DNA sequencer. Sequences were deposited at the two-parameter distances in MEGA (Kumaret al.2001), which gave identical tree topologies as in Figure 2 (data not shown). National Center for Biotechnology Information (NCBI) under
the accession nos. AY542377–AY542432. The first evolutionary model that we tested was a one-ratio model (model 0) that assumed a singlefor all branches in The␣4 and␣4-t2 genes forDrosophila viriliswere obtained
from NCBI [http://www.ncbi.nlm.nih.gov/; accession nos. the tree, whereas the other three models allowed for either two or three different values of along separate branches AF017649 (␣4) and AF017650 (␣4-t2;Belote et al.1998)];
however, alignment of the testes-specificD. virilis␣4 gene to (models 2A, 2B, and 2C, Figure 3). The first two-ratio model (model 2A, Figure 3) assumed onefor the single branch members of themelanogastergroup was ambiguous, so we did
not include this gene in our analysis. Sequences forDrosophila immediately following gene duplication (d), with another for all other branches (r). The second two-ratio model pseudoobscurawere obtained through a BLAST search of the
D. pseudoobscura genome using the ␣4, ␣4-t1, and ␣4-t2 D. (model 2B, Figure 3) assumed onefor all terminal branches of the gene being tested for selection (g) and another value melanogaster sequences, and neighbor-joining trees were
generated to confirm orthology and exclude paralogs. Genes offor all other branches in the tree (r). The three-ratio model (model 2C, Figure 3) assumed onefor the branch inD. pseudoobscurawere confirmed by comparing
correspond-ing start and stop codons with theD. melanogasteralignment immediately following duplication (d), onefor all terminal branches of the gene being tested for selection (g) and a and by examining the translated sequence for any stop codons.
Sequence analysis: DNA and protein sequences were thirdfor all other branches in the tree (r).
By comparing the likelihood of all of these models, we aligned using ClustalX (Thompsonet al.1997) and confirmed
by eye through a comparison of DNA alignments to the trans- tested (1) whether there were different selective pressures immediately following gene duplication (model 2A vs. 0); lated amino acid alignment. Measurements of polymorphism
and divergence for each gene were calculated using the pro- (2) whether there were different selective pressures along terminal branches of the gene being tested for selection gram DNAsp version 3.53 (RozasandRozas1999).
Polymor-phism was measured using two estimates:, which is deter- (model 2Bvs.0); and (3) whether there were different selec-tive pressures immediately following gene duplication, as well mined from the number of segregating sites in a sample of
genes, and, the average pairwise difference between haplo- as different selective pressures along terminal branches of the gene (model 2Cvs.2A and 2B).
types. Divergence was measured in terms of nonsynonymous
(dN) and synonymous (dS) nucleotide substitution rates as esti- Because we found significant heterogeneity in selective pres-sures within genes through a comparison of models that con-mated using the method of Yang and Nielsen (2000) in
PAML (Yang1997), which accounted for both a transition/ strainedto a single value (model 0), to a model that allowed for three different classes ofwithin a gene (model 3), we transversion bias and a codon usage bias. Differences between
polymorphism and divergence of the duplicated testes- tested the likelihood of branch-site models (YangandNielsen 2002). We used the program codeml to test whether each expressed genes (␣4-t1 and ␣4-t2) and the ancestral gene
(␣4) were compared using a Student’st-test. A phylogeny was amino acid falls into one of four site classes with three esti-mates of : o, including amino acid sites that are highly constructed using Bayesian analyses with the program MrBayes
version 3.0 (HuelsenbeckandRonquist2001), and the re- conserved across all branches;1, including amino acid sites that are weakly constrained or neutral across all branches; sulting tree was viewed using TreeView version 1.6.5 (Page
1996; Figure 1). To test for the significance of each node, and2, including two classes of sites, those either conserved or neutral on background branches ( ⬍1 or ⫽1) but posterior probabilities were calculated from a consensus tree
of all trees sampled after the Markov chain reached stationar- with ⬎1 on the branch being tested for selection. Once again we compared the likelihood of two models: model A ity, which was estimated to be at 65,000 generations. The␣7
subunit in D. melanogaster was chosen as an outgroup as it that allowed for two site classes witho⫽0 and1⫽1 and model B, a more flexible distribution that allowedoand1 diverged from the␣4 gene with early eukaryotes (Bouzatet
al.2000). to vary. Both model A and B estimate2from the data, and
the likelihood of these models was then compared to the
Tests for selection:We compared the rates of
nonsynony-mous (dN) and synonymous (dS) nucleotide substitution be- likelihood of models 1 and 3 that assumed oand 1were the same across all branches. Model 1 is a neutral model that tween duplicated genes to determine which mode of selection
played a role in the divergence of the testes-specific␣4 iso- constrainso⫽0 and1⫽1 across all branches. By comparing model A to model 1, and model B to model 3, we tested for forms. IfdN⬍dS, it suggests selective constraint against
Figure1.—Bayesian analysis of the ␣4 proteasome subunit gene family in Drosophila conducted using MrBayes version 3.0 (Huelsenbeck and Ronquist 2001). The tree was rooted using the␣7 subunit sequence fromD. melanogasteravailable at Gen-Bank (accession no. AF025793). The numbers at the nodes are the poste-rior probabilities of support for that node calculated from a consensus tree of all trees sampled after the Mar-kov chain reached stationarity at
ⵑ65,000 generations. Nodes were collapsed if the posterior probability was⬍0.70. mel,D. melanogaster; sim, D. simulans; mau,D. mauritiana; sec, D. sechellia.
B, the probabilities were then estimated that amino acid sites in the other, giving as a measure of rate correlation over sites between paralogs. A likelihood-ratio test was then used were under positive selection after gene duplication.
to test whether was significantly greater than zero, which
Tests for functional divergence:We estimated functional
would indicate functional divergence between paralogs. divergence between the␣4 gene duplicates by calculating the
coefficient of functional divergence () using the program DIVERGE version 1.04 (Gu 1999). The program examines amino acid substitution rates across duplicated genes and
RESULTS looks for correlation in evolutionary rates between paralogs.
If the evolutionary rate of an amino acid is different among
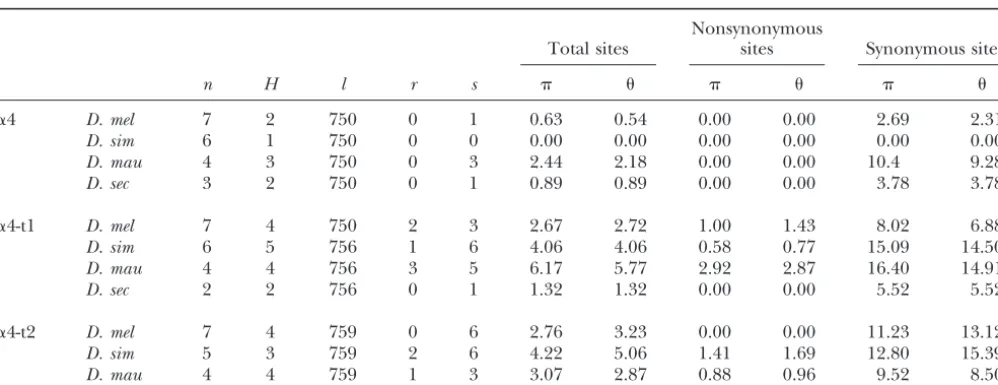
Polymorphism and divergence: Both testes-specific paralogs, it indicates functional divergence between
dupli-␣4-t1 and␣4-t2 genes have higher levels of nucleotide cated genes. More specifically, rapidly evolving sites in one
Figure2.—Tree topology used in likelihood-ratio tests of Figure3.—Representative diagram of two- and three-ratio evolutionary models using PAML (Yang1997). The numbers evolutionary models tested using PAML (Yang1997). Model on each of the branches represent the mean number of substi- 2A has one for the single branch immediately following tutions per codon along that branch. gene duplication (d), with anotherfor all other branches (r). Model 2B has one for all terminal branches of the gene being tested for selection (g) and another value of for all other branches in the tree (r). Model 2C has one (Table 1). Within each of the four species in the
melano-for the branch immediately following duplication (d), one gaster group, the ␣4 gene has no amino acid
replace-for all terminal branches of the gene being tested replace-for selection ment substitutions, with only a single synonymous change (g), and a thirdfor all other branches in the tree (r). occurring withinD. melanogasterand withinD. sechellia
and three synonymous changes within D. mauritiana.
However, in both the␣4-t1 and the␣4-t2 gene, we find allowing for variable selective pressures along branches several amino acid replacement polymorphisms within provide a better fit to the data (Table 3). Both two-ratio species. models have a significantly higher likelihood than the Both testes-specific genes also have significantly high- one-ratio model (2⌬l⫽18.9, 17.8;P⬍0.0001), indicat-er sequence divindicat-ergence between species in tindicat-erms of non- ing differences in selection acting on terminal branches synonymous nucleotide substitution rates compared to of the gene and differences in selection acting on the the ␣4 gene (Table 2) (pairwiset-tests; ␣4:␣4-t1, P ⫽ gene immediately after duplication compared to other 0.020;␣4:␣4-t2,P ⫽0.0048). In fact, the␣4-t1 gene is branches of the tree. The three-ratio model fits the data even more highly diverged than the ␣4-t2 gene (P ⫽ significantly better than both of the two-ratio models 0.032) and appears to be one of the more rapidly evolv- (2⌬l⫽27.2, 28.3;P⬍0.0001), suggesting that selective ing genes in Drosophila. The synonymous nucleotide forces on the␣4-t1 gene have changed and are different substitution rate (dS) is significantly higher in␣4-t1 rela- from the selective forces that acted on this gene immedi-tive to the somatic␣4 gene (P⫽0.044), but not signifi- ately after duplication. The estimate ofdN/dS() under cantly different from the testes-specific␣4-t2 gene (P⫽ model 2C for␣4-t1 is 0.282 after gene duplication, which 0.22). After normalizing for dS, however, the ratio of is only slightly higher than the estimate of for the dN:dSis significantly higher in␣4-t1 than in the␣4 gene gene (0.247); however, there is still a significantly better (P⫽0.00034), but there is no significant difference in fit to the three-ratio model than to either of the
two-dN:dSbetween the two testes-specific isoforms (P⫽0.15). ratio models.
TABLE 1
Nucleotide diversity and polymorphism of the␣4,␣4-t1, and␣4-t2 genes among species of theD. melanogastercomplex
Nonsynonymous
Total sites sites Synonymous sites
n H l r s
␣4 D. mel 7 2 750 0 1 0.63 0.54 0.00 0.00 2.69 2.31
D. sim 6 1 750 0 0 0.00 0.00 0.00 0.00 0.00 0.00
D. mau 4 3 750 0 3 2.44 2.18 0.00 0.00 10.4 9.28
D. sec 3 2 750 0 1 0.89 0.89 0.00 0.00 3.78 3.78
␣4-t1 D. mel 7 4 750 2 3 2.67 2.72 1.00 1.43 8.02 6.88
D. sim 6 5 756 1 6 4.06 4.06 0.58 0.77 15.09 14.50
D. mau 4 4 756 3 5 6.17 5.77 2.92 2.87 16.40 14.91
D. sec 2 2 756 0 1 1.32 1.32 0.00 0.00 5.52 5.52
␣4-t2 D. mel 7 4 759 0 6 2.76 3.23 0.00 0.00 11.23 13.12
D. sim 5 3 759 2 6 4.22 5.06 1.41 1.69 12.80 15.39
D. mau 4 4 759 1 3 3.07 2.87 0.88 0.96 9.52 8.50
n, number of sequences;H, number of haplotypes;l, length of sequence;r, number of amino acid replacement substitutions; s, number of synonymous substitutions. Diversity is multiplied by 10⫺3.
do not have a significantly higher likelihood than the branches but have ⫽∞following␣4-t2 duplication, suggesting that positive selection acted along this one-ratio model for the ␣4-t2 gene (2⌬l ⫽ 2.28, 3.2;
P⫽0.13, 0.07), suggesting that there are no significant branch withdS⫽ 0. Model B identified 15 sites in the
␣4-t2 gene to be under positive selection following du-differences in selection acting on this gene, even
follow-ing duplication. Under this line of analysis, it does not plication (Figure 4); however, it did not identify any positively selected sites following the␣4-t1 gene duplica-appear that selective constraints relaxed following gene
duplication in␣4-t2 as they did in␣4-t1. However, the tion.
Functional divergence:Estimates of the coefficient of data show a better fit to model 3 than to model 0 (2⌬l⫽
96.2;P⬍0.0001), indicating significant heterogeneity in functional divergence () are significantly greater than zero (Table 4), indicating that there is heterogeneity selection acting within the gene, so we compared
branch-site models of selection (models A and B) to allow for in evolutionary rates between homologous codons in
␣4,␣4-t1, and␣4-t2. The largest values ofare between differential selection on different codons along
individ-ual branches. the two testes-specific isoforms (␣4-t1 and ␣4-t2) and between␣4-t1 and the somatic␣4 gene. The␣4-t2 gene Both␣4-t1 and␣4-t2 show a significantly better fit to
models A and B than to models 1 and 3, respectively, and the somatic␣4 gene show a slightly higher correla-tion in evolucorrela-tionary rate, suggesting that they are not suggesting that there were differences in selection
act-ing across amino acid sites immediately after gene dupli- as functionally diverged as either of the genes is to the ␣4-t1 gene. However, there is no significant rate cation. For␣4-t1, twice the difference in likelihoods was
18.9 (P ⬍ 0.0001) for model A vs. model 1 and 15.7 correlation among amino acid sites between all three paralogs, indicating functional diversification between (P⫽0.0003) for model Bvs.model 3. For␣4-t2, twice
the difference in likelihoods was 81.2 (P⬍0.0001) for all three genes. Not all rapidly evolving sites in one paralog are rapidly evolving in the other two paralogs, model Avs.model 1 and 23.8 (P⬍0.0001) for model
Bvs.model 3. Parameters estimated under model B for suggesting that selection acts differently on homologous sites within duplicated genes.
the ␣4-t1 gene show that 78% of amino acid sites are
highly conserved across all lineages with ⫽0.047, that Pairwise comparisons between paralogs identified amino acids with higher-than-baseline probabilities of 21% of sites have ⫽0.29 across all lineages, and that
0.35% of sites are highly conserved or neutral on all site-specific rate differences (Figure 5). Although identi-fied amino acid residues with an odd ratio ⬎1 (i.e., other branches but have ⫽1.02 after␣4-t1
duplica-tion. Parameters estimated under model B for the␣4- posterior probability of a rate difference ⬎0.5) could be meaningful, a more stringent cutoff is suggested us-t2 gene show that 55% of amino acid sites are highly
conserved across all lineages with ⫽0.028, that 33% ing an odd ratio⬎2 (i.e., a posterior probability of rate difference⬎0.67;WangandGu2001). Therefore sites of sites have ⫽0.24 across all lineages, and that 12%
probabil-TABLE 2 changed their evolutionary pathways. Both have signifi-cantly higher divergence and are more polymorphic
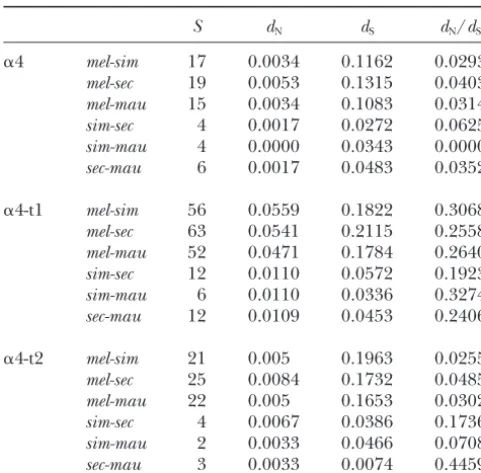
Measures of divergence in the ␣4, ␣4-t1, and ␣4-t2 genes
than the ubiquitously expressed ␣4 subunit, and the
among species of theD. melanogastercomplex
␣4-t1 gene seems to be one of the genes more highly S dN dS dN/dS diverged betweenD. melanogasterandD. simulans. These results are in agreement with previous findings of lower
␣4 mel-sim 17 0.0034 0.1162 0.0293
polymorphism and divergence of genes with wider tissue
mel-sec 19 0.0053 0.1315 0.0403
expression patterns (CoulthartandSingh1988) and
mel-mau 15 0.0034 0.1083 0.0314
of lower divergence of genes that are expressed in a
sim-sec 4 0.0017 0.0272 0.0625
sim-mau 4 0.0000 0.0343 0.0000 greater number of tissues (Duret and Mouchiroud
sec-mau 6 0.0017 0.0483 0.0352 2000).
Rapid evolution of duplicated genes: The classical
␣4-t1 mel-sim 56 0.0559 0.1822 0.3068
view of gene duplication is that the duplicate copy is
mel-sec 63 0.0541 0.2115 0.2558
free to evolve neutrally for some period of time, before
mel-mau 52 0.0471 0.1784 0.2640
obtaining a new function with associated selective
con-sim-sec 12 0.0110 0.0572 0.1923
sim-mau 6 0.0110 0.0336 0.3274 straints (Ohno1970). However, we were unable to find sec-mau 12 0.0109 0.0453 0.2406 evidence that the classical model pertains to the evolu-tion of␣4-t1 and ␣4-t2, as both genes show evidence
␣4-t2 mel-sim 21 0.005 0.1963 0.0255
that selective constraints were retained at the majority
mel-sec 25 0.0084 0.1732 0.0485
of sites following gene duplication, as opposed to
accu-mel-mau 22 0.005 0.1653 0.0302
mulating neutral mutations. Selective constraints in the
sim-sec 4 0.0067 0.0386 0.1736
sim-mau 2 0.0033 0.0466 0.0708 ␣4-t1 gene were relaxed following duplication and then sec-mau 3 0.0033 0.0074 0.4459 appeared to have increased only slightly over time, which may explain the higher polymorphism found in S, fixed nucleotide substitutions between species;dS, average
this gene. Our estimates of dN and dS after the early number of synonymous nucleotide substitutions per
synony-mous site between species;dN, average number of nonsynony- stages of␣4-t1 duplication are not consistent with neu-mous nucleotide substitutions per nonsynonyneu-mous site be- tral evolution in the broadest sense; however, a small tween species as calculated using the method ofYangand
portion of sites do show evidence of complete release Nielsen(2000).
of selective pressures. In the␣4-t2 gene we were also unable to detect broad-scale neutral evolution following gene duplication, but rather than finding a complete ity⬎0.67) may be more conservatively identified as
be-release of selective pressures on a small portion of sites, ing functionally diverged and provide a starting point
we detected signatures of positive selection having acted for further investigations into the functional differences
on several codons following duplication. among␣4,␣4-t1, and ␣4-t2.
More recently it was reported that likelihood-ratio tests of branch-site models (Yang and Nielsen 2002) DISCUSSION frequently detect positive selection in error under cer-tain conditions (Zhang 2004). Computer simulations We have shown here that both Drosophila␣4
testes-performed by Zhang indicate that when selective con-specific isoforms are evolving in a manner very different
straints are relaxed along branches being tested for se-from that of the more ubiquitously expressed␣4
sub-lection, positive selection is erroneously detected in up unit. The␣4 subunit shows very few nucleotide
substitu-to 70% of cases. However, when we tested branch-site tions and almost no polymorphism within the
melanogas-models in ␣4-t1, we did not detect positive selection
ter clade and seems to be among the more slowly
acting after gene duplication, despite evidence that se-evolving genes in Drosophila. Proteasomes have an
es-lective constraints were relaxed following␣4-t1 duplica-sential function in a variety of biological processes and
tion, similar to an evolutionary scheme that was shown are required to specifically degrade a diverse array of
to give a high rate of false positives. However, we did proteins across many different tissues, which may
ex-detect positive selection in␣4-t2 using branch-site mod-plain why the ubiquitously expressed␣4 subunit exhibits
els, but we found no evidence that selective constraints such strong functional and selective constraint.
How-on average were significantly relaxed or changed How-on the ever, there is only weak evidence that essential genes
branch following␣4-t2 duplication, a condition under evolve slower than nonessential genes (Yanget al.2003),
which the branch-site models may perform more accept-and proper reproductive functioning could also be
con-ably (Zhang 2004). Therefore, despite a high rate of sidered essential for an organism’s total fitness that
false detections of positive selection using branch-site includes both viability and reproductive ability. Both
testes-models of evolution under certain conditions, these specific␣4 subunits have become specialized in
TABLE 3
Parameter estimates from tests for selection
Model Log likelihood Parameter estimates
M0: one-ratio (one-site class) ⫺4263.70 ⫽0.0970
M3: discrete (two-site classes) ⫺4215.59 p0⫽0.595,0⫽0.0278 p1⫽0.405,1⫽0.220
M1: neutral ⫺4431.01 p0⫽0.379
p1⫽0.621
Branch-specific models
␣4-t1 2A: two-ratio ⫺4254.26 d⫽0.277;r⫽0.0778
2B: two-ratio ⫺4254.81 g⫽0.254;r⫽0.0797
2C: three-ratio ⫺4240.66 d⫽0.282;g⫽0.247;r⫽0.0537
␣4-t2 2A: two-ratio ⫺4262.56 d⫽0.502;r⫽0.0941
2B: two-ratio ⫺4262.10 g⫽0.132;r⫽0.0882
2C: three-ratio ⫺4260.83 d⫽0.357;g⫽0.130;r⫽0.0850
Branch-site models
␣4-t1 MA ⫺4421.55 p0⫽0.349;p1⫽0.481
p2⫹3⫽0.170,2⫽0.657
MB ⫺4207.72 p0⫽0.784,0⫽0.0472
p1⫽0.212,1⫽0.286 p2⫹3⫽0.00348,2⫽1.02
␣4-t2 MA ⫺4390.41 p0⫽0.391;p1⫽0.449
p2⫹3⫽0.160,2⫽∞
MB ⫺4203.69 p0⫽0.553,0⫽0.0281
p1⫽0.330,1⫽0.244 p2⫹3⫽0.117,2⫽∞
⫽dN/dS:dis the estimate ofon the branch immediately following gene duplication,gis the estimate offor all terminal branches leading to the gene, and ris the estimate offor the remaining branches. p⫽proportion of sites in each class of(0, 1, 2, and 3):p0is the proportion of sites that are highly conserved across branches,p1is the proportion of sites that are weakly constrained or are neutral across branches,p2is the proportion of sites that have ⬎1 on the branch being tested for selection but are highly conserved on other branches,p3is the proportion of sites that have ⬎1 on the branch being tested for selection but are neutral or weakly constrained on other branches.
␣4-t2 evolution, making it less likely that positive selec- retained by losing separate subfunctions from a multi-functional ancestral gene. Both testes-specific isoforms tion was detected in error.
The DDC model (Forceet al.1999) provides a better seem to be missing at least two functional regions com-pared to the somatic␣4 subunit (Belote et al. 1998), explanation for the evolution of the testes-specific ␣4
proteasome subunits from the data that we have, rather including a putative nuclear localization signal and the KEKE motif that may be important for protein-protein than a neutral accumulation of mutations. The DDC
model states that duplicated genes become selected and interactions. Similarly, the tissue specificity of␣4-t1 and
TABLE 4 faster than the other, 65 pairs had significantly different ratios of dN:dS, suggesting changes in functional
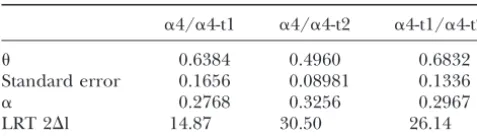
con-Estimates of the coefficient of functional divergence ()
straint, and 113 of these genes had ratios ofdN:dS ⬎1, suggesting positive selection. Moreover, they report that
␣4/␣4-t1 ␣4/␣4-t2 ␣4-t1/␣4-t2
in most duplicated genes, the gene that is evolving faster
0.6384 0.4960 0.6832 also has a higher rate of synonymous nucleotide substi-Standard error 0.1656 0.08981 0.1336
tutions, similar to the high value ofdSthat we found in
␣ 0.2768 0.3256 0.2967
␣4-t1. In Arabidopsis significantly reduced nucleotide
LRT 2⌬l 14.87 30.50 26.14
polymorphism in newly duplicated genes suggests that
␣, gamma shape parameter of rate variation across sites. selective sweeps are common following duplication and that positive selection plays an important role in dupli-cate gene evolution (MooreandPurugganan2003).
␣4-t2 compared to the more ubiquitously expressed so- Positive selection in duplicated genes is probably com-matic ␣4 subunit is suggestive of a more specialized mon across most taxa, as a study including bacteria, function that could have involved the loss of functional archae, and eukaryotes found that ratios of nonsynony-domains. Higher polymorphism in both␣4-t1 and␣4- mous-to-synonymous substitutions do not indicate wide-t2 suggests that functional constraints were either re- spread neutral evolution following gene duplication laxed or neutral relative to the somatic ␣4 gene, but (Kondrashovet al.2002).
do not clearly distinguish between selection regimes. Gene duplication and reproductive specialization: However, likelihood-ratio tests of evolutionary models The development of tissue-specific patterns of expres-suggest that both testes-specific isoforms experienced sion compared to the ancestral gene is a common fate either incomplete relaxed selective constraints at the of duplicated genes (LynchandForce2000); however, majority of sites (as in␣4-t1) or positive selection on a a specific type of subfunctionalization that has separated portion of sites (as in␣4-t2) as opposed to broad-scale reproductivevs.somatic functions of the␣4 gene seems neutral evolution during periods of loss of functional to have occurred. By partitioning these two processes domains and tissue specialization. Moreover, our tests there may be a greater flexibility in reproductive trait for functional divergence indicate that evolutionary evolution, which may allow for a specialization of testes rates among all three duplicate genes are different at function. A high number of genes show testes-specific many homologous amino acids (Figure 3). It is therefore expression in Drosophila, similar in proportion to those feasible that, due to subfunctionalization, the testes- specifically expressed in the brain (Andrews et al. specific␣4 proteasome subunits were not as constrained 2000). Genes with testes-biased expression may have at as many nucleotide sites as was the somatic gene, an increased chance of being under positive selection which allowed a higher rate of degenerative mutations (Meiklejohnet al.2003), and therefore genes that de-as predicted by the DDC model (Force et al.1999). velop new functions may be more important in males Relaxed selective constraints and positive selection due to a possible higher turnover of testes-expressed may be common following gene duplication (see Intro- genes (Oliver2003). Even within the Drosophila pro-duction). Similar patterns of selection are also seen in teasome, six of the 20S proteasome subunits and four other gene duplicates that are expressed in the male of the 19S regulatory cap subunits have also undergone testes.Maxwellet al.(2003) discovered that a member gene duplications resulting in testes-specific isoforms of the-defensin gene family that has a high expression (Maet al.2002), suggesting a trend toward the develop-level in the testes and brain exhibits signatures of posi- ment of a specialized male reproductive functioning tive selection. Similarly, the testes-specific Drosophila proteasome in Drosophila. It seems that gene duplication
nuclear transport factor-2-related gene has evolved more may be a major factor promoting increasing complexity rapidly under positive selection than the parental gene and reproductive specialization in the Drosophila protea-(Betra´nandLong2003). We report similar processes some and that reproductivevs.somatic subfunctionaliza-following ␣4 gene duplications, with relaxed selective tion is an important factor allowing for such specializa-constraint acting on ␣4-t1 and signatures of positive tion.
Figure5.—Pairwise comparisons between paralogous genes of the␣4 gene family identified sites showing evolutionary rate differences in Drosophila. The program DIVERGE version 1.04 (Gu1999) was used to identify amino acids with higher-than-baseline probabilities of site-specific rate differences. A more conservative cutoff for identifying functionally diverged sites is suggested to be when probabilities of rate heterogeneity are⬎0.67 (WangandGu2001).
evidence that the␣4-t1 gene diverged from␣4 more re- conclusive, the broader involvement of the newer copy (␣4-t1) may suggest that newly arisen genes are freer cently than␣4-t2 did, with a posterior probability of 95%
that ␣4 forms a clade with ␣4-t1 separate from ␣4-t2. from selective constraints to form biochemical linkages and thus provide new, alternate pathways for the evolu-Similarly, ␣4-t1 may not exist inD. virilis (Belote et al.
1998), or in D. pseudoobscura as ␣4-t2 does, supporting tion of new genetic systems. Sex- and reproduction-related genes are recognized as a class of rapidly evolv-the possibility that evolv-the ␣4-t1 gene duplicated after the
divergence ofD. virilisandD. pseudoobscurafrom themela- ing genes (SinghandKulathinal2000;Swansonand Vacquier 2002), particularly genes involved in male
nogasterclade. However, we cannot discount the possibility
that␣4-t1 has become a pseudogene in these species or reproduction (for examples, see Swanson and Vac-quier1995;Wyckoffet al.2000;Swansonet al.2001; that it has become too diverged for detection through
homology searches. Torgersonet al.2002), but it is unclear how the rapid evolution of genes within a genetic system affects the The␣4-t2 subunit appears to be involved only in
sper-matid elongation, whereas␣4-t1 is involved in spermatid evolutionary pathways of other genes within that system. For example, a high rate of evolutionary change in pro-elongation as well as being expressed in the primary
Glickman, M. H., andA. Ciechanover, 2001 The ubiquitin-protea-(or, alternatively, vice versa). This is similar to the
coevo-some proteolytic pathway: destruction for the sake of construc-lutionary relationship between male and female fertil- tion. Physiol. Rev.82:373–428.
ization proteins, for example, in abalone where rapid Goldman, N., andZ. Yang, 1994 A codon-based model of nucleotide substitution for protein-coding DNA sequences. Mol. Biol. Evol. evolution of the egg protein VERL elicits rapid evolution
11:725–736.
of the sperm protein lysin (Galindoet al.2003). There Gu, X., 1999 Statistical methods for testing functional divergence is likely a complex network of reproductive proteins after gene duplication. Mol. Biol. Evol.16:1664–1674.
Hellberg, M. E., G. W. MoyandV. D. Vacquier, 2000 Positive whose interactions may drive the rapid evolution of
re-selection and propeptide repeats promote rapid interspecific di-productive traits and even a single gene that evolves vergence of a gastropod sperm protein. Mol. Biol. Evol.17:458– rapidly may have a dramatic effect on the evolutionary 466.
Huelsenbeck, J. P., andF. R. Ronquist, 2001 MrBayes: Bayesian rates of the network of interactions that it may have
inference of phylogeny. Bioinformatics17:754–755.
with other genes. In this way sexual selection may affect Hughes, A. L.,1994 The evolution of functionally novel proteins an even wider variety of genes involved in reproduction, after gene duplication. Proc. R. Soc. Lond. B Biol. Sci. 256:
119–124. as we see not only rapid evolution of male reproductive
Johnson, M. E., L. Viggiano, J. A. Bailey, M. Abdul-Rauf, G. Good-proteins, but also the rapid and unique evolution of the
winet al., 2001 Positive selection of a gene family during the
␣4 testes-specific proteins that are involved in recycling emergence of humans and African apes. Nature413:514–519. Kondrashov, F. A., I. B. Rogozin, Y. I. WolfandE. V. Koonin, them.
2002 Selection in the evolution of gene duplications. Genome We thank Alberto Civetta and Richard Morton for their helpful Biol.3:research0008.1–0008.9.
comments on sequence analysis and manuscript revisions, and Paulo Kumar, S., K. Tamura, I. B. JakobsenandM. Nei, 2001 MEGA2: Nuin, Frances Raftis, and Alex Robertson for help with Bayesian analy- molecular evolutionary genetics analysis software. Bioinformatics
17:1244–1245. sis. We also owe thanks to Rob Kulathinal for insightful discussions
Lynch, M., andA. Force, 2000 The probability of duplicate gene during the preliminary stages of the project, to Ziheng Yang, and to
preservation by subfunctionalization. Genetics154:459–473. two anonymous reviewers for their many helpful comments. This work
Ma, J., E. KatzandJ. M. Belote, 2002 Expression of proteasome was supported by the Natural Sciences and Engineering Research
subunit isoforms during spermatogenesis inDrosophila
melanogas-Council of Canada through a grant to R.S.S. and a postgraduate
ter.Insect Mol. Biol.11:627–639. scholarship to D.G.T.
Martinez-Castilla, L. P., andE. R. Alvarez-Buylla, 2003 Adap-tive evolution in theArabidopsisMADS-box gene family inferred from its complete resolved phylogeny. Proc. Natl. Acad. Sci. USA 100:13407–13412.
LITERATURE CITED Massingham, T., L. J. Daviesand P. Lio`, 2001 Analysing gene function after duplication. BioEssays23:873–876.
Andrews, J., G. G. Bouffard, C. Cheadle, J. Lu, K. G. Beckeret Mathews, S., J. G. BurleighandM. J. Donoghue, 2003 Adaptive al., 2000 Gene discovery using computational and microarray
evolution in the photosensory domain of phytochrome A in early analysis of transcription in the Drosophila melanogaster testis. angiosperms. Mol. Biol. Evol.20:1087–1097.
Genome Res.10:2030–2043.
Maxwell, A. I., G. M. Morrisonand J. R. Dorin, 2003 Rapid Armbrust, E. V., andH. M. Galindo, 2001 Rapid evolution of a sequence divergence in mammalian-defensins by adaptive
evo-sexual reproduction gene in centric diatoms of the genus
Thalas-lution. Mol. Immunol.40:413–421. siosira. Appl. Environ. Microbiol.67:3501–3513.
Meiklejohn, C. D., J. Parsch, J. M. RanzandD. L. Hartl, 2003 Belote, J. M., M. MillerandK. A. Smyth, 1998 Evolutionary
con-Rapid evolution of male-biased gene expression in Drosophila. servation of a testes-specific proteasome subunit gene in
Drosoph-Proc. Natl. Acad. Sci. USA100:9894–9899.
ila.Gene215:93–100.
Moore, R. C., andM. D. Purugganan, 2003 The early stages of Betra´n, E., and M. Long, 2003 Dntf-2r, a young Drosophila
ret-duplicate gene evolution. Proc. Natl. Acad. Sci. USA100:15682– roposed gene with specific male expression under positive
Dar-15687. winian selection. Genetics164:977–988.
Ohno, S., 1970 Evolution by Gene Duplication. Springer Verlag, New Bouzat, J. L., L. K. McNeil, H. M. Robertson, L. F. Solter, J. E.
York. Nixonet al., 2000 Phylogenomic analysis of the␣proteasome
Ohta, T., 1994 Further examples of evolution by gene duplication gene family from early-diverging eukaryotes. J. Mol. Evol. 51:
revealed through DNA sequence comparisons. Genetics 138: 532–543.
1331–1337. Civetta, A., andR. S. Singh, 1995 High divergence of reproductive
Oliver, B., 2003 Fast males. Heredity91:535–536. tract proteins and their association with postzygotic reproductive
Page, R. D. M., 1996 TREEVIEW: an application to display phyloge-isolation in Drosophila melanogaster andDrosophila virilis group
netic trees on personal computers. Comput. Appl. Biosci. 12: species. J. Mol. Evol.41:1085–1095.
357–358. Coulthart, M. B., andR. S. Singh, 1988 High level of divergence of
Rodriguez-Trelles, F., R. Tarrı´oandF. J. Ayala, 2003 Conver-male-reproductive-tract proteins, betweenDrosophila melanogaster
gent neofunctionalization by positive Darwinian selection after and its sibling species,D. simulans.Mol. Biol. Evol.5:182–191.
ancient recurrent duplications of the xanthine dehydrogenase Duret, L., andD. Mouchiroud, 2000 Determinants of substitution
gene. Proc. Natl. Acad. Sci. USA100:13413–13417. rates in mammalian genes: expression pattern affects selection
Rozas, J., andR. Rozas, 1999 DnaSP version 3: an integrated pro-intensity but not mutation rate. Mol. Biol. Evol.17:68–74.
gram for molecular population genetics and molecular evolution Fares, M. A., andK. H. Wolfe, 2003 Positive selection and
subfunc-analysis. Bioinformatics15:174–175. tionalization of duplicated CCT chaperonin subunits. Mol. Biol.
Singh, R. S., 1990 Patterns of species divergence and genetic theo-Evol.20:1588–1597.
ries of speciation, pp. 231–265 inTopics in Population Biology and
Ferris, P. J., C. Pavlovic, S. FabryandU. W. Goodenough, 1997
Evolution, edited by K.Wohrmannand S. K.Jain. Springer-Ver-Rapid evolution of sex-related genes in Chlamydomonas. Proc.
lag, New York. Natl. Acad. Sci. USA94:8634–8639.
Singh, R. S., andR. J. Kulathinal, 2000 Sex gene pool evolution Force, A., M. Lynch, F. B. Pickett, A. Amores, Y.-L. Yanet al., 1999
and speciation: a new paradigm. Genes Genet. Syst.75:119–130. Preservation of duplicate genes by complementary, degenerative
Stoltzfus, A., 1999 On the possibility of constructive neutral evolu-mutations. Genetics151:1531–1545.
tion. J. Mol. Evol.49:169–181. Galindo, B. E., V. D. VacquierandW. J. Swanson, 2003 Positive
Swanson, W. J., andV. D. Vacquier, 1995 Extraordinary divergence selection in the egg receptor for abalone sperm lysin. Proc. Natl.
the acrosomal process of abalone spermatozoa. Proc. Natl. Acad. Wyckoff, G. J., W. WangandC.-I Wu, 2000 Rapid evolution of male reproductive genes in the descent of man. Nature403: Sci. USA92:4957–4961.
304–309. Swanson, W. J., andV. D. Vacquier, 2002 The rapid evolution of
Yang, Z., 1997 PAML: a program package for phylogenetic analysis reproductive proteins. Nat. Rev. Genet.3:137–144.
by maximum likelihood Comput. Appl. Biosci.13:555–556. Swanson, W. J., A. G. Clark, H. M. Waldrip-Dail, M. F. Wolfner
Yang, Z., andR. Nielsen, 2000 Estimating synonymous and nonsyn-andC. F. Aquadro, 2001 Evolutionary EST analysis identifies
onymous substitution rates under realistic evolutionary models. rapidly evolving male reproductive proteins in Drosophila. Proc.
Mol. Biol. Evol.17:32–43. Natl. Acad. Sci. USA98:7375–7379.
Yang, Z., andR. Nielsen, 2002 Codon-substitution models for de-Thompson, J. D., T. J. Gibson, F. Plewniak, F. JeanmouginandD. G.
tecting molecular adaptation at individual sites along specific Higgins, 1997 The CLUSTAL_X windows interface: flexible
lineages. Mol. Biol. Evol.19:908–917. strategies for multiple sequence alignment aided by quality
analy-Yang, J., Z. GuandW.-H. Li, 2003 Rate of protein evolution versus sis tools. Nucleic Acids Res.24:4876–4883.
fitness effect of gene delection. J. Mol. Evol.20:772–774. Torgerson, D. G., R. J. KulathinalandR. S. Singh, 2002
Mamma-Yuan, X., M. MillerandJ. M. Belote, 1996 Duplicated proteasome lian sperm proteins are rapidly evolving: evidence of positive
subunit genes inDrosophila melanogasterencoding testes-specific selection in functionally diverse genes. Mol. Biol. Evol.19:1973–
isoforms. Genetics144:147–157. 1980.
Zhang, J., 2004 Frequent false detection of positive selection by the Tsaur, S.-C., andC.-I Wu, 1997 Positive selection and the molecular
likelihood method with branch-site models. Mol. Biol. Evol.21: evolution of a gene of male reproduction,Acp26AaofDrosophila.
1332–1339. Mol. Biol. Evol.14:544–549.
Zhang, J., H. F. RosenbergandM. Nei, 1998 Positive Darwinian Tsaur, S.-C., C.-T. TingandC.-I Wu, 1998 Positive selection driving
selection after gene duplication in primate ribonuclease genes. the evolution of a gene of male reproduction,Acp26Aa, ofDrosoph- Proc. Natl. Acad. Sci. USA95:3708–3713.
ila: II. Divergence versus polymorphism. Mol. Biol. Evol. 15:
Zhang, J., Y. ZhangandH. F. Rosenberg, 2002 Adaptive evolution 1040–1046. of a duplicated pancreatic ribonuclease gene in a leaf-eating Van de Peer, Y., J. S. Taylor, I. BraaschandA. Meyer, 2001 The monkey. Nat. Genet.30:411–415.
ghost of selection past: rates of evolution and functional diver- Zhang, P., Z. GuandW.-H. Li, 2003 Different evolutionary patterns gence of anciently duplicated genes. J. Mol. Evol.53:436–446. between young duplicate genes in the human genome. Genome Wang, Y., andX. Gu, 2001 Functional divergence in the caspase Biol.4:R56.
gene family and altered functional constraints: statistical analysis